
Abstract
Foot and mouth disease virus (FMDV) was isolated from sloughed tongue epithelium of Egyptian cattle presenting with mouth lesions and ropy salivation in two Egyptian governorates (El-Fayoum and Dakahlia). The virus was isolated in Madin-Darby bovine kidney (MDBK) cells and identified by reverse transcription-polymerase chain reaction (RT-PCR). The complete genome was obtained by next generation sequencing. The strains isolated from El-Fayoum and Dakahlia were serotype A and O, respectively and both isolates had identity with the previously reported Egyptian strains. This study reports successive outbreaks of FMDV that occurred in Egypt during 2015–2016 and describes the dynamics of two outbreaks in addition to the use of cardiac biomarkers in the diagnosis of FMD-related myocarditis in calves and its clinical relevance. Serum cardiac troponin1 (cTn I) and creatinine kinase myocardial band (CK-MB) were measured. Mean serum cardiac troponin1 (cTn I) showed significant increase (P < 0.001) in FMDV-infected calves. The increase in fatal and recovered cases was (2.794 ± 0.502 ng/mL) and (1.196 ± 0.443 ng/mL), respectively, compared to the healthy control cases (0.014 ± 0.002 ng/mL). Thus, the serum cTn-I successfully diagnosed FMD-associated myocarditis in calves but not prognostic for the fatal cases. The FMDV sequences described in this study should further help in studying FMDV endemicity in Egypt, tracking the source of infection, selection of control strategies and vaccine updates. The study also determines the clinical relevance of cardiac biomarkers in diagnosis of FMDV-related myocarditis in infected calves.
1 Introduction
Foot and mouth disease (FMD) causes huge economical loss in Egypt in terms of reduction in the production of red meat, milk and milk byproducts. The disease is caused by foot-and-mouth disease virus (FMDV), which is a member of the genus Aphthovirus under Picornaviridae [Citation1]. Molecular epidemiology studies on FMDV in Egypt are lacking. Such studies are necessary to better understand the natural history of the disease, relationships among various serotypes, and to develop more effective prevention and control methods.
The FMDV is a small, non-enveloped, virus with an icosahedral capsid composed of 60 copies each of four structural proteins (VP1, VP2, VP3 and VP4), which surround a single stranded positive sense RNA genome of approximately 8500 nucleotides [Citation2]. The virus is classified into seven immunologically distinct serotypes i.e., A, C, O, Asia 1, SAT 1, SAT 2, and SAT 3. The viral genome is further subdivided into P1, P2 and P3 regions. The P1 region encodes leader proteinase (Lpro) and structural proteins 1A (VP4), 1B (VP2), 1C (VP3), and 1D (VP1). The P2 region encodes non–structural proteins 2A, 2B, and 2C, while P3 region encodes 3A, 3B (VPg), 3C protease and 3D polymerase [Citation3,Citation4]. The VP1 gene at the surface of the virion is the most variable and contains serotype-specific amino acid sequence that can help differentiate various serotypes [Citation3].
The FMD is a highly contagious transboundary disease that affects cloven-footed domesticated and wild animals in Africa, Asia and parts of South America [Citation5]. Over the last 65 years, the disease has become endemic in Egypt with three strains (A, O and SAT 2). The first FMD outbreak in Egypt was reported in 1950 and it was due to serotype SAT2 [Citation6]. The SAT2 reinvaded the country in 2012 and caused high mortality in ruminants [Citation7].
Egypt suffered from FMD outbreaks in 1958 with serotypes A and O; in 1961, 1964, 1965, 1970, 1974, 1983, 1987, 1989, 1993 and 2000 with serotype O; and in 1967 and 1972 with serotype A [Citation8]. On February 2006, serotype A caused six outbreaks in Ismailia and 12 additional outbreaks in seven other Egyptian governorates e.g., Alexandria (2 outbreaks), Behera (1 outbreak), Cairo (1 outbreak), Dakahlia (1 outbreak), Dumyat (5 outbreaks), Fayum (1 outbreak), and Menofia (1 outbreak). By April 6, 2006, 34 outbreaks of disease were reported. The results of the molecular typing suggested a relationship between Egyptian and East African strains [Citation9]. In 2012–2013, A serotype of Asian origin related to Iranian strain was detected [Citation10]. Serotype O has a long history of causing regular outbreaks in Egyptian livestock [Citation11,Citation12]. The outbreaks of FMD still occur all over Egypt although vaccination is obligatory in the country [Citation10,Citation13].
The FMDV is associated with sudden death in young calves. In such cases, no observable clinical signs are seen except for virus-induced damage in the myocardium [Citation14]. Generalized myositis aphthosa affects only calves during the acute stage. Other muscles such as hind quarter, diaphragm and tongue are also affected [Citation15]. Histopathological changes in the skeletal muscle are similar to those in heart during the acute stage of the disease [Citation16]. Recently, serum cardiac troponin1 (cTn I) and creatinine kinase myocardial band (CK-MB) have been used as diagnostic biomarkers as they are released into the blood soon after any micro injury in the myocardium [Citation14].
The reports of new FMD variants in the country have raised question related to genetic diversity of these variants as well as the need for better understanding of these variants. To answer some of these questions, we characterized FMDV isolates based on complete genome sequencing in order to understand the dynamics of serotypes A and O in calves. This follow up study also focused on mortality in calves and the relevance of cardiac biomarkers in diagnosis and prediction of mortality in infected calves.
2 Materials and methods
In the 2014–2015 outbreak, molecular epidemiology and evolution of FMDV in two provinces (El-Fayoum and Dakahlia) were investigated. During sampling, signalment and history from owners and veterinarians revealed the main complaint to be of high fatality rate in calves in spite of vaccination. In the 2015–2016 outbreak, the chief complaint was the same as above. Hence, we concentrated our efforts to predict the fatal cases before death to reduce the losses.
2.1 Molecular epidemiology and evolution study
2.1.1 Clinical samples
In December 2014 and January 2015, 20 samples of sloughed tongue epithelium were collected from cattle showing fever, ropy salivation with slobbering, smacking of lips and lameness. Ten samples were collected from each of the two governorates i.e., El-Fayoum and Dakahlia. The samples were homogenized in Eagle’s minimum essential medium (MEM) followed by centrifugation at 2000×g for 20 min. The supernatants were inoculated in baby hamster kidney-21 (BHK21) cells followed by incubation for 2 h at 37 °C. This was followed by the addition of MEM with Earle’s salts supplemented with 2% fetal bovine serum, 100 μg/mL streptomycin, and 100 U/mL penicillin. Inoculated cells were harvested when cytopathic effects (CPE) appeared in about 80% of the monolayer (about three days post inoculation). The infected cells were frozen and thawed three times followed by centrifugation at 1200g for 20 min.
2.1.2 Reverse transcription-polymerase chain reaction (RT-PCR)
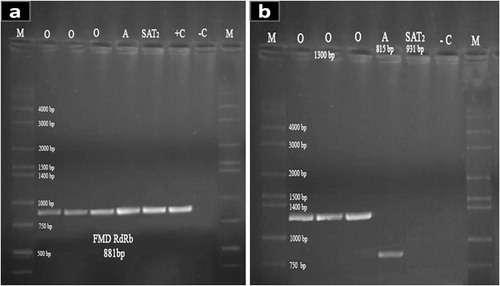
The QIAamp Viral RNA Mini kit (Qiagen, Valencia, CA) was used for viral RNA extraction from supernatants of infected cells. Conserved RdRp primer pair of forward 5′-TTCGAGAACGGCACDGTCGGA-‘3 and reverse 5′-CACGGAGATCAACTTCTCCTG-‘3 sequences was used to amplify a 881 bp product that confirmed the presence of FMDV infection [Citation10]. The serotype differentiation was done by using VP1 primers (). Extracted RNAs were subjected to RT-PCR using one-step RT-PCR kit (Qiagen, Valencia, CA). Amplification reactions (25 μL reaction mixture) were performed in a thermocycler (Mastercycler Eppendorf) under the following conditions: 50 °C for 30 min for reverse transcription and initial denaturation at 95 °C for 15 min; then 35 cycles consisting of 94 °C for 1 min for denaturation followed by annealing for 1 min at 52 °C for serotype O and SAT2 while for RdRp and serotype A, the reaction run at 54 °C, 55 °C, respectively. Extension was done at 72 °C for 1 min followed by one final extension step of 10 min at 72 °C. The amplicons were analyzed by 1.2% agarose gel electrophoresis in Tris-acetate EDTA buffer followed by staining with ethidium bromide.
Table 1 Forward and reverse VP1 primers with expected amplicon size and annealing temperatures.
2.1.3 Next generation sequencing (NGS)
One representative RNA sample from each of the governorates was subjected to whole genome sequencing. Total RNA was quantified by a fluorometric RiboGreen assay. The library was created using Illumina’s Truseq RNA sample preparation kit (Cat. # RS-122-2001) and sequenced using Illumina MiSeq platform with 250 paired end cycle run. The CLC genomic workbench 6.0 (http://www.clcbico.com) was used to analyze the resulting NGS reads, including trimming, sequence quality, and de novo assembly. Assembled contigs were analyzed by BLASTx and open reading frame was predicted by the ORF finder tool (https://www.ncbi.nlm.nih.gov/gorf/orfig.cgi).
2.1.4 Genome characterization and phylogenetic analysis
The study sequences were compared with previously reported FMDV sequences available in GenBank. Sequences were aligned by using the Clustal W in MEGA 6.0 (Molecular Evolutionary Genetic Analysis). A phylogenetic tree was constructed using the best fit Maximum Likelihood model in MEGA 6.0 based on lowest BIC score (Bayesian Information Criterion). The Maximum-Likelihood method used for constructing phylogenetic tree was GTR mode + G (Gamma distribution with 5 rate categories) for serotype O and GTR+G+I for serotype A [Citation17]. The evolutionary distances were computed using the p-distance in MEGA 6.0. The FMDV A and O serotype sequences were submitted to GenBank with accession numbers KP940474 and KP940473, respectively.
2.2 Prognostic study (cardiac biomarker study)
2.2.1 Animals used in follow up and cardiac marker detection
During the outbreak of 2015–2016, 119 saliva and nasal swabs were collected from suspected infected calves. Calves were aged 2–15 months belonging to El-Fayoum and Dakahlia governorates. Infection with serotypes A and O was confirmed by RT-PCR in 108 calves (40 males and 68 females). Blood samples were collected once upon admission. Serum samples were collected from blood and stored at -20 °C until used. Cardiac troponin I and CK-MB screening was performed in all confirmed infected calves. Ten clinically healthy calves with normal appetite, alertness, and good vital signs of the same age were used as a control. The control calves were belonged to a private farm in Dakahlia province, with no history of FMD infection. Follow up of all infected calves was done for up to 14 days.
2.2.2 Clinical assessment and laboratory tests
All confirmed infected calves were subjected to thorough clinical examination with vital signs monitoring. Special attention was paid to heart and lung examination. Transthoracic ultrasonographic examination of the heart and lung was performed. Serum samples were examined for serum cTn I by an enzyme-linked immunosorbent assay (ELISA) kit (Monobind Inc, Lake Forest, CA, USA) and CK-MB activity was measured using commercially available kits (Spectrum diagnostics, Germany). These tests were applied according to the manufacturers’ instructions.
2.2.3 Postmortem and histopathological examination
Postmortem examination of lung and heart was performed for all dead calves. Lesions were observed and samples were collected. Lung and heart specimens were taken and fixed in 10% buffered formalin followed by histopathological examination [Citation18].
2.2.4 Statistical analysis
The cardiac biomarkers screening results were statistically analyzed by one-way analysis of variance (ANOVA) using the GLM procedures of SPSS (2008); version 17.0. Statements of statistical significance are based on P ≤ 0.05. Differences among the means were estimated using the post hoc Duncan's new multiple range test [Citation19].
3 Results
3.1 Virus isolation
Cells on which the primary 20 samples were inoculated demonstrated CPE three days after inoculation and the titer of harvested viruses by TCID50 ranged between 102 and 104 log10. The FMD infection was confirmed in all samples by obtaining 881 bp size single bands with RdRp primers. All samples were negative against SAT2 serotype while ten samples of El-Fayoum were serotype A positive and other ten samples from Dakahlia were serotype O positive, without any mixed infection in both ().
3.2 Next generation sequencing
A representative isolate from El-Fayoum farm gave nearly complete genome of serotype A consisting of 7281 nt arranged as partial 164 nt 5′UTR, 6996 nt in ORF and partial 121 nt 3′UTR. The ORF nt can translate to 2331 aa. Representative strain from Dakahlia farm, however, was positive for serotype O and yielded a nearly complete genome sequence consisting of 7765 nt arranged as partial 669 nt 5′UTR, 6999 nt ORF, and partial 97 nt 3′UTR. ORF can translate to 2332 aa.
3.3 Phylogenetic analysis
The serotype A study sequence Egypt/2014/ KP940474/A showed 99.9% nt and aa identity with previously reported serotype A sequence from Egypt (EGY 9/2011; KC440881) (; ). The comparison of ORFs of study sequence with reference sequence EGY 9/2011 showed three synonymous substitutions from T to C at positions 615, 3117 and 4020 and three non-synonymous substitutions at positions 4148 (G to A), 4178 (A to G) and 4907 (C to T) which reflected amino acid changes, including Lysine to Arginine, Arginine to Glutamine and Valine to Alanine at position 1383, 1393 and 1636, respectively. Serotype A study sequenced showed low nt (84.8%) and aa (91.6%) identity with Egypt/2006/JF749843.

Table 2 Nucleotides and putative amino acid identities of serotype O/Egypt/2014/KP940473 and A/Egypt/2014/KP940474 complete ORF compared with representative serotypes.
Egypt/2014/ KP940473/O strain belonged to the same lineage and shared 94.5% and 92% nt identity with the closest known relatives e.g., Turkish (O1manisa/AY593823) and Indian (AY593828) strains, respectively, while aa identity was 97.3% with both (, ). The Egypt/2014/KP940473/O serotype had only 88.9% nt and 96.3% aa identity with the recently characterized strain from Egypt/2013 (KC440883). The aa alignment of both Egyptian sequences revealed that, of the 2332 aa along the genome, there was mutation in 85 aa (3.64%) ().

Table 3 Amino acid substitutions in O/Egypt/2014/KP940473 ORF genome in comparison with the latest isolated O serotype from Egypt AGZ15284/Egypt/2013.
3.4 Follow up of the outbreaks (prognostic study)
The number of calves confirmed to have FMD in El-Fayoum and Dakahlia governments in successive outbreaks (2015–2016) is shown in . By the end of the disease course (1–14 days), 37 (34.25%) and 71(65.74%) calves of 108 infected cases were died or completely recovered, respectively. The age of the fatal cases was 2–14 months, while in the recovered cases, it was 11–15 months. Clinical examination in the cases that ended by death revealed anorexia, recumbency, hurried respiration, tachycardia or bradycardia with cardiac arrhythmias, and variations of body temperature. These findings were associated with or without classical signs of FMD. Clinical findings in recovered cases included inappetence, fever, tachycardia with rhythmic pulse, excessive salivation, and vesicular lesions on tongue, feet and lameness.
Table 4 Number of recovered and dead calves after FMD serotype A and O outbreak in Egyptian El-Fayoum and Dakahlia governments 2015–2016.
Clinical alterations were observed during the course of the disease; the calves that were moribund and those prior to die showed inappetence to anorexia (86.5%), dullness (73%), incoordination (86.5%), recumbency (73%), constipation (37.8%), and sometimes loud breathing sound (27%). The vital signs alterations include tachycardia (75.6%), bradycardia (24.3%), cardiac arrhythmias (100%), and tachypnoea (rhythmic and shallow hurried respiration) (100%). The body temperature ranged from 36.5 to 41.5 °C. These findings associated with infrequently detected light ulcerations in the tongue and dental pad (27%). Auscultation of the heart revealed cardiac arrhythmias (100%) without any adventitious sounds or murmurs. Percussion of the lungs showed variable degree of bilateral symmetrical dullness ventrally (54%) while the lung auscultation revealed exaggerated vesicular sound (81%) with or without auscultation of the bronchial sound beyond its normal situation (51.3%).
During the disease course, the recovered cases hold body temperature ranged from (39.2–41.7 °C) that usually accompanied by slight tachycardia with rhythmic pulse and tachypnoea, excessive salivation, vesicular lesions on tongue and feet, and lameness were recorded. After recovery, fever subsides with retrieval of normal pulse and respiratory rate ranges ().
Table 5 Clinical alterations recorded during examination of fatal and recovered cases.
Ultrasonographic examination of the lungs revealed that 100% of the fatal cases had absence of reverberation artifacts and appearance of hypoechoic tissue within lower third of both lungs which sometimes extended dorsally. In all recovered cases, the reverberation artifacts of the lung was the only image. The ultrasonographic examination of heart did not reveal any detectable changes in heart dimensions and myocardium echogenicity in both fatal and recovered cases.
3.5 Cardiac marker detection in confirmed infected calves
Laboratory examination revealed that mean serum CTn I and CK-MB levels were significantly higher (P < 0.001) in fatal cases than in recovered cases and control group. The mean serum cTn I was significantly higher (P < 0.001) in recovered group than control one while, no significant was detected between both groups for CK-MB levels. The high levels of cTn I and CK-MB were detected in 37 (100%) and 20 (54%) calves out of 37 fatal cases, respectively. Mean while the increase levels of cTn I and CK-MB were detected in 29 (48.8%) and 24 (33.8%) among 71recovered cases, respectively ().
Table 6 Levels of CTn I and CK-MB in FMD infected dead and recovered calves compared with healthy control calves.
3.6 Postmortem and histopathological findings
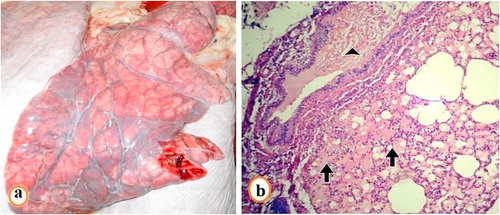
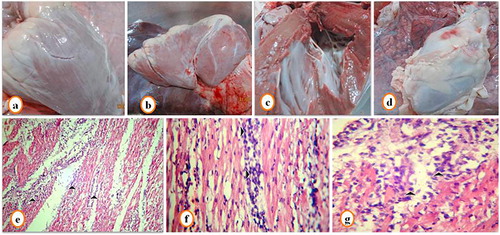
Postmortem examination of lungs in fatal cases revealed pulmonary congestion. Heart showed slight white streaks on the myocardium (a–c). Hemorrhages on the pericardial surface was observed in 5 of 37 fatal cases (). No other gross abnormalities were observed. Histopathological examination revealed presence of marked pulmonary edema with obliteration of alveoli with a homogenous pink fluid (). The heart showed myocarditis with myocardial degeneration and necrosis, and infiltration of muscle fiber with mononuclear cells especially lymphocytes (–).
4 Discussion
FMD molecular epidemiological studies can determine the variation and genetic relationship among the circulating FMDV serotypes in Egypt. This genetic variation can partially explain persistent occurrence of outbreaks even with obligatory governmental vaccination [Citation10,Citation13,Citation20]. The SAT2 serotype was not detected in Egypt after the 1950s but it re-invaded the country in 2012 while, both serotypes A and O are regularly detected [Citation7,Citation20,Citation21]. This study was aimed to quantify the extent of genetic diversity of FMDV in Egypt by sequencing. The importance of nt sequencing to show diversity among FMDV stains began with Beck and Strohmaier [Citation22]. Since differences in VP1 sequences are the basis for developing RT-PCR tests to identify different serotypes of FMDV, we used primers amplifying the VP1 region [Citation10,Citation20,Citation23]. We were unable to differentiate among subtypes as there were no substitutions in VP1 sequences indicating that naively using VP1 sequence comparison can achieve only limited success Carrillo et al. [Citation3] Bazid et al. [Citation24]. Previous VP1 sequences in the study by Salem et al. [Citation25] revealed that there were two distinct isolates of serotype O; one related to PanAsia-2 on Upper Egypt and Delta (Monofya and Kafer El-Shikh) and other related to East African strains in Sharkia governments. In addition, there were two distinct serotype A; one related to A-Iran 05 and the other related to African strains. Serotype SAT2 was detected in Aswan, Gharbia, Sharkia and Kafr El-Shikh. Further studies by Salam et al. [Citation21] revealed that type A belonged to Asia topotype and Iran 05 lineage, type O belonged to ME-SA topotype and PanAsia-2 lineage, and type SAT2 belonged to topotype VII. Phylogenetic analysis by Soltan et al. [Citation20] identified two distinct strains characterized as serotype A (African topotype) and serotype O topotype EA-3. Phylogenetic analysis concluded that SAT2 was the cause of the FMD outbreak during 2012–2013 [Citation26]. The results of our study are in contrast to those of Soltan et al., Salem et al. and Elhaig et al. [Citation20,Citation25,Citation26] but are in agreement with Salam et al. [Citation21]. We did not isolate any SAT2 serotype or serotypes A, O African strains, as both serotypes were related to Asian strains. We agree with Salem et al. [Citation25] that vaccine seeds need to be updated to include the current circulating serotypes of FMDV with continuous monitoring of the genetic changes in viruses from different locations in Egypt.
The next generation sequencing revealed many nt substitution in serotypes A and O; 48 (3.64%) aa substitution were identified in 85 sites. Eleven aa substitutions were detected in capsid sequences (VP1-4) (). Many aa substitutions were observed in capsid sequences of all analyzed serotypes by Lloyd-Jones et al. [Citation27]. The role of nt and aa substitutions in virulence and/ or immunity response needs more investigation. The increased number of detected substitutions indicates the presence of many serotypes and subtypes that increase the potential for genetic recombination and creation of new variant strains [Citation28]. Continued studies by next generation sequencing are recommended for the detection of new subtypes, tracking the source of infection, and updates in control strategy and vaccination.
The follow up of successive outbreaks (2015–2016) indicated that serotype O is the most predominant one; in fact, it is the most prevalent of the seven serotypes in many parts of the world [Citation29]. Most deaths occur due to serotype O outbreaks because of gaps in immunity against the current genetically different serotypes [Citation30].
Assessment of vital signs is a standard component of clinical examination in diseased animals. In FMD-infected calves, vital sign alterations indicate significant cardiopulmonary pathological changes that can be confirmed by physical examination of heart and lung. The previous investigations recorded FMD associated myocarditis in young infected calves (under 2 months) [Citation14,Citation31–Citation33]. Unexpectedly, our investigation recorded fatal cases in calves ranging in age from 2 to 14 months.
The great clinical alterations in FMD infected calves include the body temperature, pulse rate, respiration rate, and irregular cardiac rhythm in auscultation without FMD vesicular lesions also reported in suckling calves by Karapinar et al. [Citation33]. The myotropism of FMDV may occur with or without concurrent vesicular lesions. The presence of both myotropism and vesicles defined as expanded multi-tropism (the muscle and the epithelium), while the absence of vesicles suggests the substitution of tropism (the muscle, but not the epithelium) [Citation34].
Further ultrasound, examination of the heart did not give evidence for myocarditis and have not any benefit. Electrocardiogram (ECG) can be more efficient and more informative in early myocarditis diagnosis [Citation35]. Unfortunately, ECG did not included in the present study and further investigation is still in need.
In agreement with Aktas et al. and Karapinar et al. [Citation14,Citation33], the cardiopulmonary signs were mostly present in fatal cases, while the classical FMD signs were observed mostly in recovered cases. Sudden death in young calves is usually associated with damage to the myocardium [Citation14], which can be detected and evaluated through cTnI and CK-MB measurements. The plasma cTnI level in calf that died from FMD can reach 037.24 ng/mL in comparison to 0.01–0.02 ng/mL in the control group [Citation33]. Significant increase in cTn I was previously detected in cases of FMD myocarditis (11.48 ± 0.67 μg/L) in comparison to the non-myocarditis group (0.03 ± 0.06 μg/L) [Citation14]. Our study revealed significant increase in cTn I in fatal and recovered cases of FMD (). The significant increase (P < 0.001) in cTn I in both fatal and recovered cases of FMD infected calves may have attributed to the myotropism of the virus to the heart tissue. The significant increase (P < 0.001) in cTn I levels in fatal cases as compared to the recovered cases may be described as sub-clinical myocarditis in recovered cases. Cardiac arrhythmias, rather than myocarditis, may be additional causes for elevation of cTn I levels; 100% of fatal cases showed cardiac arrhythmia but none of the recovered cases showed arrhythmia. A relationship between cardiac arrhythmias and elevated levels of cTn I was reported in humans [Citation36].
Variation in the values of cTnI and CK-MB within the same diseased groups may be related to date of sampling post infection and the stage of clinical course. In the early stage following myocardial damage in humans, cardiac troponins appear in blood after 2 to 4 h and last up to 10 to 21 days [Citation37]. The time course of cardiac troponin release in blood as a response to myocardial damage has not been determined in cattle. In sheep, significant elevation in plasma cTnI concentration after one day of myocardial damage and a gradual decrease to physiological levels occur over the next 14 days. The cTn I levels of 16.93 ng/mL, 10.12 ng/mL and 5.46 ng/mL were detected at the first, second, and third day, respectively [Citation38].
The cTnI marker was superior to CK-MB for detection of myocyte injury in FMD infected groups. The marker cTn I has higher cardiac specificity, rapid release, and long half-life time in the blood. Meanwhile, CK–MB clears rapidly from the blood and returns to normal levels rapidly. So, it must be determined as soon as possible after the onset of symptoms [Citation39]. Although, both CK-MB and cTnI levels are useful and not mutually exclusive in the diagnosis, they cannot predict the risk of mortality in FMD infected calves [Citation40].
Postmortem examination of dead and euthanized calves revealed grossly normal hearts with only slight white streaks on the myocardium that were not highly demarcated. In agreement with Gunes et al. [Citation31], the heart showed normal consistency without softness and no other abnormalities were observed. There was no difference in post mortem examination of the heart between cases that died after 1 day and those died after 14 days from onset of the disease, suggestive another cause of death may be present. In contrast, Tunca et al. [Citation41] reported that the myocardium was soft and flaccid in consistency. Similar to Gunes et al. [Citation31], histopathological findings were included myocardial degeneration, necrosis, infiltrations of muscle fiber with mononuclear cells especially lymphocytes and few plasma cells [Citation33]. Finally, if we assume that the cause of death originated from the heart, it could be said that cardiac arrhythmias rather than myocarditis is the culprit for death in FMD infected calves.
5 Conclusions
The primarily investigation of FMD outbreak in two Egyptian governorates (El-Fayoum and Dakahlia) by RT-PCR revealed circulation of FMD serotypes A and O. Further investigation of the consecutive outbreaks indicates circulation of both serotypes in the two governorates with more endemicity of serotype O. The finding can proof a presence of vaccination failure against both serotypes. A complete genome sequences for both serotypes with complete relation analysis with associated viruses can help in detection of the source of infection and designing of new modified vaccines. Cardiac biomarkers (cTnI and CK-MB) can be used to detect and expect the degree of myocyte injury in FMD infected calves with superiority of cTnI over CK-MB.
Competing interests
None of the authors of this paper has a financial or personal relationship with other people or organization that could inappropriately influence or bias the content of the paper.
Acknowledgements
The authors thank the cattle farm owners and their veterinarians in Egypt for their co-operation. The authors are grateful to Dr. Hiam M. Fakhry, researcher in FMD unit in Veterinary Serum and Vaccine Research Institute, East of Cairo, Egypt for helping in virus isolation and identification.
Notes
Peer review under responsibility of Faculty of Veterinary Medicine, Cairo University.
References
- Knowles NJ, Hovi T, Hyypiä T, King AMQ, Lindberg AM, Pallansch MA, et al. Picornaviridae. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz E, editors. Viral taxonomy: 1st edition ninth report of the international committee on taxonomy of viruses; 2011, p. 855–880.
- M.J.GrubmanB.BaxtFoot and mouth diseaseClin Microbiol Rev172004465493
- C.CarrilloE.R.TulmanG.DelhonZ.LuA.CarrenoA.VagnozziComparative genomics of foot and mouth disease virusJ Virol79200564876504
- G.LinZ.CaiJ.WuX.WanL.XuG.RandyIdentifying a few foot-and-mouth disease virus signature nucleotide strings for computational genotypingBMC Bioinf92008279289
- N.J.KnowlesJ.HeY.ShangJ.WadsworthB.Valdazo-GonzálezH.OnosatoSoutheast Asian foot and mouth disease viruses in Eastern AsiaEmerg Infect Dis182012499501
- FAO (Food and agriculture organization). History of FMD disease in Egypt; 1999. http://www.fao.org/ag/againfo/commissions/eufmd/commissions/eufmd-home/reports/archive/33rd-general-session/fmd-in-egypt/en/.
- M.ShawkyM.Abd El-AtyH.M.FakryH.M.DaoudE.I.El-SayedW.G.MossadIsolation and molecular characterization of foot and mouth disease sat2 virus during outbreak 2012 in EgyptJ Vet Adv2013320126068
- W.VoslooA.D.S.BastosO.SangareS.K.HargreavesG.R.ThomsonReview of the status and control of foot and mouth disease in Sub-Saharan AfricaRev Sci Tech212002437449
- N.J.KnowlesJ.WadsworthS.M.ReidK.G.SwabeyA.A.El-KholyA.O.Abd El-RahmanFoot and mouth disease virus serotype A in EgyptEmerg Infect Dis13200715931596
- N.M.SobhyS.K.MorM.E.M.MohammedI.M.BastawecyH.M.FakhryC.R.B.YoussefPhylogenetic analysis of Egyptian foot and mouth disease virus endemic strainsJ Am Sci102014133138
- K.S.ZaherW.M.AhmedImpact of foot and mouth disease on oxidative status and ovarian activity in Egyptian buffaloesWorld J Zool3200817
- M.F.MandourM.M.AbdEl-daimS.A.M.AbdelwahabH.I.Abu ElnagaM.S.ElshahidyA.M.AzabMolecular characterization of foot and mouth disease viruses collected from Suez Canal area, Egypt from 2009 to 2011Global Anim Sci J1201311391153
- M.K.EL-BayoumyK.A.AbdelrahmanA.M.AllamT.K.FaragH.A.A.Abou-ZeinaM.A.KutkatMolecular characterization of foot and mouth disease virus collected from Al Fayoum and Beni-Suef governorates in EgyptGlob Vet132014828835
- M.S.AktasY.OzkanlarE.OrucI.SozdutmazA.KirbasMyocarditis associated with foot and mouth disease in suckling calvesVeterinarski arhiv852015273282
- R.L.CrowellB.J.LandauJ.SiakPicornarvirus receptors in pathogenesisK.Lonberg-HolmL.PhilipsonVirus receptors (PART 2)1981Chapman and Hall Ltd.London169184
- W.J.HadlowDiseases of skeletal muscleJ.R.M.InnesL.Z.SaundersComparative neuropathology1962Academic press Inc.London147243
- K.TamuraG.StecherD.PetersonA.FilipskiS.KumarMEGA, 6.0, Molecular evolutionary genetics analysis version 6.0Mol Biol Evol30201327252729
- M.SlaouiL.FietteHistopathology procedures: from tissue sampling to histopathological evaluationMethods Mol Biol69120116982
- D.B.DuncanMultiple range and multiple F testsBiometrics111995142
- M.A.SoltanA.H.NegmaldinM.M.El-DiastyS.M.G.MansourM.A.ElbadryR.P.WilkesMolecular characterization of circulating Foot and mouth disease virus (FMDV) serotype O topotype EA-3 and serotype A (African topotype) genotype IV in Egypt, 2016Vet Microbiol20820178993
- H.M.SalamA.A.MahmoudA.M.KhadrS.A.SalemT.M.ElshemeyA.H.AbdelrahmanIsolation, sequencing and phylogenetic analysis of FMD strains during 2011–2012 outbreaks in EgyptInt J Agri Inn Res34201423192473
- E.BeckK.StrohmaierSubtyping of European foot and mouth disease virus strains by approaches nucleotide sequence determinationJ Virol61198716211629
- Y.StramT.MoladD.ChaiB.GelmanH.YadinDetection and subtyping of foot-and-mouth disease virus in infected cattle by polymerase chain reaction and amplified VP1 sequencingJ Vet Diagn Invest719955255
- A.I.BazidH.A.HusseinS.S.BalalA.A.ElsanousiB.M.AhmedPhylogenetic analysis of foot and mouth disease virus type O in Egypt 2009Int J Virol1020142836
- Salem SH, Arafa A, Abohatab E, Saad A, Ahmed HA. Genotyping of foot and mouth disease virus (FMD) in Egypt during 2011–2012. 1st Conf of An Health Res Inst Assoc 2012; 411–419.
- M.M.ElhaigM.N.ElsheeryMolecular investigation of foot-and-mouth disease virus in domestic bovids from GharbiaEgypt. Trop Anim Health Prod46201414551462
- K.Lloyd-JonesM.MahapatraS.UpadhyayaD.J.PatonA.BabuG.HutchingsGenetic and antigenic characterization of serotype O FMD viruses from East Africa for the selection of suitable vaccine strainVaccine35201768426849
- M.MahapatraS.UpadhyayaS.AvisoA.BabuG.HutchingsS.ParidaSelection of vaccine strains for serotype O foot-and-mouth disease viruses (2007–2012) circulating in Southeast AsiaEast Asia Far East Vaccine35201771477153
- A.R.SamuelN.J.KnowlesFoot and mouth disease type O viruses exhibit genetically geographically distinct evolutionary lineages (topotypes)J Gen Virol822001609621
- J.DomenechJ.LubrothK.SumptionImmune protection in animals: the examples of rinderpest and foot-and-mouth diseaseJ Comp Path1422010 S120 - 24
- V.GunesH.M.ErdoganM.CitilK.OzcanAssay of cardiac troponins in the diagnosis of myocardial degeneration due to foot and mouth disease in a calfVet Rec1562005714715
- M.Y.GulbaharW.C.DawisT.GuvencM.YarimU.ParlakY.B.KabakMyocarditis associated with foot and mouth disease virus type O in lambsVet Pathol442007589599
- T.KarapinarD.DabakT.KulogluH.BulutHigh cardiac troponin I plasma concentration in a calf with myocarditisCan Vet J512010379399
- J.ArztB.BaxtM.J.GrubmanT.JacksonN.JuleffJ.RhyanThe pathogenesis of foot-and mouth disease II: Viral pathways in swine, small ruminants, and wildlife; myotropism, chronic syndromes, and molecular virus host interactionsTransbound Emerg Dis582011305326
- T.MorgeraA.Di LenardaL.DreasB.PinamontiF.HumarR.BussaniElectrocardio-graphy of myocarditis revisited: Clinical and prognostic significance of electrocardiographic changesAm Heart J1241992455467
- G.P.RajanR.ZellwegerCardiac troponin I as a predictor of arrhythmia and ventricular dysfunction in trauma patients with myocardial contusionJ Trauma572004801808
- S.KorffH.A.KatusE.GiannitsisDifferential diagnosis of elevated troponinsHeart922006987993
- F.LeonardiB.PasseriA.FussariP.De RazzaC.BeghiP.LorussoCardiac Troponin I (cTnI) concentration in an ovine model of myocardial ischemiaRes Vet Sci852008141144
- M.WeberM.RauK.MadlenerA.ElsaesserD.BankovicV.MitrovicDiagnostic utility of new immunoassays for the cardiac markers cTnI, myoglobin and CK-MB massClin Biochem38200510271030
- G.KornK.PotelThe lesions of foot and mouth disease in the skeletal muscles of calvesArch Exp Veterinar med81954606625
- R.TuncaM.SozmenH.ErdoganM.CitilE.UzluH.OzenDetermination of cardiac troponin I in the blood and heart of calves with foot and mouth diseaseJ Vet Diagn Invest202008598605