
Abstract
Covalently closed circular DNA (cccDNA) is the transcriptional template of hepatitis B virus (HBV). Extensive research over the past decades has unveiled the important role of cccDNA in the natural history and antiviral treatment of chronic HBV infection. cccDNA can persist in patients recovering from acute HBV infection for decades. This explains why HBV reactivation occasionally occurs in patients with resolved hepatitis B receiving intensive immunosuppressive agents. In addition, although advances in antiviral treatment dramatically improve the adverse outcomes of chronic hepatitis B (CHB), accumulating evidence demonstrates that current antiviral treatments alone, be they nucleos(t)ide analogs (NAs) or interferon (IFN), fail to cure most CHB patients because of the persistent cccDNA. NA suppresses HBV replication by directly inhibiting viral polymerase, while IFN enhances host immunity against HBV infection. Viral rebound often occurs after discontinuation of antiviral treatment. The loss of cccDNA can be induced by non-cytolytic destruction of cccDNA or immune-mediated killing of infected hepatocytes. It is known that NA has no direct effect on viral transcription or cccDNA stability. Therefore, the long half-life of hepatocytes leads to a very slow decline in cccDNA in patients under antiviral therapy. Novel antiviral agents targeting cccDNA or cccDNA-containing hepatocytes are thus required for curing chronic HBV infection.
Introduction
Hepatitis B virus (HBV) infection is a key global public health concern. More than 350 million people are chronically infected with HBV.Citation1 The advent of modern antiviral therapy has dramatically reduced the mortality and morbidity of patients with chronic hepatitis B (CHB). Currently, there are two types of antiviral therapy for HBV infection: nucleoside/nucleotide analogs (NAs) and interferon (IFN).Citation2 The former suppresses HBV replication by inhibiting its polymerase; the latter does so by its direct antiviral effects as well as indirect modulation of the host immune response. Only a small portion of patients can undergo HBsAg seroclearance with antiviral therapy; this response has been associated with several host and viral factors.Citation3,Citation4 Unfortunately, viral rebound often occurs following discontinuation of antiviral therapy.Citation5,Citation6,Citation7 Accumulating evidence has proven that the persistence of covalently closed circular DNA (cccDNA) is the major barrier to eradicating chronic HBV infection.Citation8 In addition, persistent cccDNA can occasionally be detected even in the liver of patients with resolved hepatitis B infections.Citation9,Citation10,Citation11,Citation12 More importantly, the persistent cccDNA appears to be replication competent at least in a significant portion of these patients, as evidenced by the fact that HBV reactivation occurs when they receive intensive immunosuppressive agents, such as chemotherapy. This review will discuss the molecular mechanisms of persistence of HBV cccDNA and its clinical significance as well as the potential strategies to cure chronic HBV infection.
cccDNA IS THE TRANSCRIPTIONAL TEMPLATE OF HBV
The life cycle of HBV
HBV is the prototype member of the family Hepadnaviridae. The genome of HBV is composed of partially double-stranded DNA, also known as relaxed-circular DNA (RC-DNA); it is 3.2 kb in length. Although HBV is a DNA virus, it replicates through reverse transcription.Citation13,Citation14 The replication cycle of HBV in infected hepatocytes has been extensively studied (Figure ).Citation15 Recently, sodium taurocholate cotransporting polypeptide (NTCP) has been identified as either the receptor or part of the receptor complex for HBV infection.Citation16 Following entry into hepatocytes, HBV undergoes uncoating of its capsid and releases its RC-DNA. The RC-DNA is then translocated to the nuclei of infected hepatocytes, where it is converted to cccDNA through a series of processes that have not been fully elucidated. The details of this process will be described below.
Figure 1 The replication cycle of HBV. HBV virions bind to the receptor NTCP on hepatocytes and are internalized. Nucleocapsids are released into the cytoplasm and then translocated to the nucleus, where the genome is converted into cccDNA through a poorly understood mechanism, most likely via the DNA repair mechanism. The HBV cccDNA serves as the template for transcription of the pregenomic and subgenomic RNAs. The pregenomic RNA is the template for both reverse transcription and translation of polymerase and core proteins. The polymerase binds to the packaging signal of the pregenomic RNA, and both of them are incorporated into the viral capsid, inside which RC-DNA is generated through reverse transcription. The resulting RC-DNA can either be enveloped in ER and secreted as progeny virions or be recycled back to the nucleus for cccDNA amplification. ER, endoplasmic reticulum; NTCP, sodium taurocholate cotransporting polypeptide.
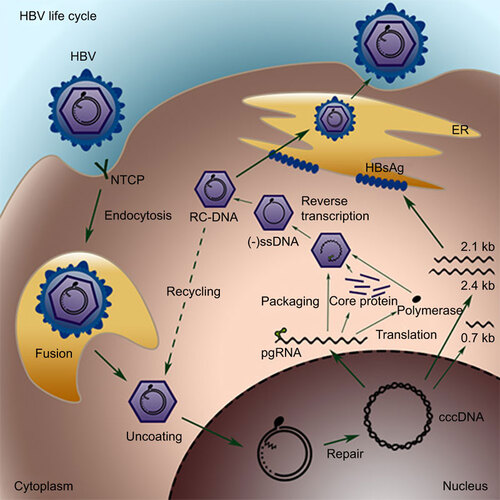
The resulting cccDNA is the template for transcription of the pregenomic RNA (pgRNA) and several subgenomic RNAs. The genome of HBV contains four open reading frames, precore/core, polymerase, preS1/S2/S and X domains, which are organized in an overlapping manner. The transcription of these four open reading frames is controlled by four distinct promoters and is regulated independently.Citation17,Citation18 The pgRNA serves as the template for reverse transcription and translation of the core and polymerase proteins. The viral polymerase binds to the packaging signal of the pgRNA and subsequently initiates the assembly of core proteins to a viral capsid, which incorporates the pgRNA–polymerase complex in the cytoplasm. Inside the capsid, the pgRNA undergoes reverse transcription, which is initiated by protein priming at the tyrosine residue of the polymerase. The resulting RC-DNA is partially double stranded. The minus strand is longer than a genome length, with a covalently bound polymerase and a redundant flap at the 5′ end. The plus strand synthesis is incomplete, and a gap exists down to the 3′ end. The mature capsid can be enveloped with HBV surface antigens in the endoplasmic reticulum (ER) and secreted to the blood. It can also re-enter the nuclei through a process called intracellular recycling to replenish the cccDNA pool.Citation19,Citation20 It has been estimated that the cccDNA pool can reach an average of 5–50 copies per cell, most likely through this intracellular recycling replenishment process.Citation19,Citation20,Citation21,Citation22
Conversion of RC-DNA to cccDNA
By comparison of the structures of RC-DNA and cccDNA, several required steps are deduced.Citation14 First, the 3′ downstream gap on the plus strand must be filled. Whether the viral polymerase or a cellular polymerase elongates the 3′ end of the plus strand remains unclear. The covalently bound polymerase at the 5′ end of the minus strand needs to be removed. The part of the cell in which the removal of the polymerase occurs is still debated; however, some previous studies have suggested that the bound polymerase is cleaved in the cytoplasm, and the resultant protein-free RC-DNA is subsequently transported to the nucleus.Citation23,Citation24,Citation25 The flap DNA and the RNA primer, located at the 5′ ends of the minus and plus strand DNA, respectively, have to be removed before ligation of the nicked DNA. It is generally assumed that DNA repair mechanisms play a role in this process. However, the host factors involved in this process remain elusive. Recently, it has been demonstrated that tyrosyl-DNA-phosphodiesterase 2 can cleave the bound polymerase at least in vitro, although its role in cccDNA formation in vivo is unclear.Citation26
PERSISTENCE OF HBV cccDNA DURING THE NATURAL HISTORY OF HBV INFECTION
Given that cccDNA is the transcriptional template for HBV, a correlation between the levels of intrahepatic cccDNA and serum HBV DNA during the natural history of HBV infection is expected. A cross-sectional natural history study revealed that cccDNA levels are consistently significantly higher in HBeAg-positive patients compared to HBeAg-negative patients, inactive carriers, and patients who underwent HBsAg clearance.Citation27 This is also consistent with observations of higher levels of intrahepatic HBV replicative intermediates and serum HBV DNA in HBeAg-positive patients.
HBsAg seroconversion, defined by the loss of HBsAg and appearance of anti-hepatitis B antibodies, is a typical serological consequence in patients with transient HBV infection and can also occur in a small proportion of patients with chronic hepatitis B.Citation28 It is generally believed that HBsAg seroconversion is closely related to the complete clearance of HBV infection. However, even after HBsAg seroconversion, HBV DNA is occasionally detectable in the liver and even the serum of patients. Previous studies have demonstrated that despite active maintenance of robust antiviral T-cell immunity, the HBV genome can persist in patients recovering from acute HBV infection for decades.Citation9,Citation10,Citation11 In particular, intrahepatic HBV cccDNA can sometimes be detected in these HBsAg-negative patients.Citation12,Citation27,Citation29 The intrahepatic cccDNA level in HBsAg-negative patients is usually lower than that in HBsAg-positive patients.Citation26 These patients are commonly denoted as having ‘occult HBV infection’, which is defined by the presence of the HBV genome in the liver and sometimes also the serum of HBsAg-negative patients.Citation30,Citation31
Notably, the HBV cccDNA that persists in the infected hepatocytes is replication competent rather than replication defective. This is evidenced by the fact that HBV reactivation occasionally occurs in patients with occult HBV infection who receive intensive immunosuppressive chemotherapy, particularly in lymphoma patients with rituximab-containing regimens.Citation32,Citation33 In addition, occult HBV infection carriers may be a source of HBV transmission through blood transfusion and orthotopic liver transplantation.Citation34 Therefore, the persistent intrahepatic replication-competent cccDNA in these patients provides a mechanistic explanation for these alarming events.
PERSISTENCE OF HBV cccDNA IS THE PRIMARY BARRIER TO ERADICATING CHRONIC HBV INFECTION
Experimental evidence from model systems
Curing chronic HBV infection requires elimination of HBV cccDNA from infected hepatocytes; therefore, it is important to know the stability and half-life of HBV cccDNA. By inhibition of viral replication with NAs, it is possible to estimate the stability and half-life of cccDNA in vitro and in vivo. Initially, the stability of cccDNA was determined in a tissue culture system by using non-dividing primary hepatocytes or hepatoma cell lines. However, the estimated half-life of cccDNA in culture systems is often confounded by the death of cultured cells, resulting in inconsistent conclusions from different systems. For example, using the congenitally infected primary duck hepatocyte culture system, the half-life of duck hepatitis B virus (DHBV) cccDNA was found to be very short, ranging from 3 to 5 days.Citation35 In contrast, studies in primary woodchuck hepatocytes and HepaRG cells showed that cccDNA was quite stable under the inhibition of NAs.Citation36,Citation37,Citation38
The half-life of hepadnavirus has also been investigated in vivo. Using inhibition of viral replication by NAs, the half-lives of the cccDNAs of DHBV and woodchuck hepatitis virus (WHV) in vivo were estimated to be 33–50 days and 35–57 days, respectively.Citation39,Citation40 Short-term antiviral NA treatment fails to cure hepadnavirus infection, and withdrawal of NAs inevitably leads to rebound viremia.Citation41,Citation42 Additionally, the use of the potent reverse transcriptase inhibitor entecavir in DHBV-infected ducks further suggested that the cccDNA is maintained by its stability rather than ongoing replication.Citation43 Notably, the half-life measurements in these studies are confounded by uncertainties. For example, NAs alone may not be able to completely inhibit viral replication in vivo. In addition, the turnover and division of infected hepatocytes may also affect the decline rate of the cccDNA. All these factors should be taken into consideration when the half-life of cccDNA is measured.
Reduction of HBV cccDNA in patients during current antiviral treatments
Current antiviral treatment involves two types of agents, IFN and NAs. NAs only block reverse transcription and have no direct effect on the existing cccDNA minichromosome, which is the template for viral transcription. This is analogous to the observation that highly active antiretroviral therapy has no effect on the integrated provirus of HIV.Citation44 No cure for HIV has been achieved by highly active antiretroviral therapy alone. Likewise, NA alone does not cure CHB. However, in contrast to highly active antiretroviral therapy for HIV, NA also fails to prevent the de novo formation of cccDNA from incoming virions.Citation45,Citation46,Citation47 Despite its lack of a direct effect on cccDNA, NA treatment can partially and transiently restore the immune response to HBV infection and may thus indirectly promote viral clearance.Citation48,Citation49
The intrahepatic level of cccDNA has been shown to be an indicator for treatment response.Citation50 To monitor the reduction of cccDNA during antiviral therapy, several assays to quantify intrahepatic cccDNA were developed.Citation27,Citation50,Citation51 The quantification of HBV cccDNA in infected cells is a challenging task. The cccDNA quantitative assays typically rely on a plasmid-safe DNase treatment and cccDNA-specific primers for amplification and detection of cccDNA.Citation27 However, the specificity of the quantitative PCR for cccDNA is always a concern because a vast excess of RC-DNA is present in infected hepatocytes.Citation52 A more reliable cccDNA quantitative assay is clearly needed. Interestingly, HBsAg levels are correlated with intrahepatic cccDNA levels in HBeAg-positive patients.Citation8,Citation27,Citation53 Recent studies also suggested that the serum HBsAg level can predict clinical outcomes in natural history and antiviral treatment of CHB.Citation54,Citation55 Therefore, quantification of HBsAg can serve as a surrogate marker of intrahepatic cccDNA. By using the cccDNA quantitative assay, it has been found that treatment with adefovir, entecavir or lamivudine for 48 weeks may result in the reduction of intrahepatic cccDNA by 0.8–1.0 log.Citation27,Citation56 Although significant, this reduction appears to be insufficient for the eradication of HBV cccDNA because viral rebound often occurs after withdrawal of NAs. Therefore, cccDNA persists even under long-term NA therapy.
In contrast, IFN is an immune modulatory agent. Despite a higher rate of adverse effects, IFN is an attractive therapeutic option because it provides higher rates of off-therapy immune control of HBV as well as HBsAg/HBeAg loss or seroconversion.Citation4,Citation57 The mechanisms by which IFN controls HBV infection include direct inhibition of HBV replication and clearance of infected hepatocytes through stimulation of cell-mediated immunity. Furthermore, using human chimeric severe combined immunodeficiency mice that lack adaptive immune cells, it was found that pegylated IFN-α reduced serum HBsAg and HBeAg levels by inhibiting viral transcription independent of immune cells; however, it did not significantly reduce intrahepatic cccDNA.Citation58 Interestingly, a recent study reported that IFN could result in the specific and non-hepatotoxic degradation of intrahepatic cccDNA.Citation59 However, data are scarce regarding the effects of IFN treatment on the reduction of intrahepatic cccDNA in patients, partly because a reliable intrahepatic quantitative cccDNA assay was not available until recently. A combination treatment with pegylated IFN-α2b and adefovir has been shown to induce a strong cccDNA decline of 2.4 logs and HBsAg reduction.Citation60 Another study demonstrated that combination therapy with pegylated IFN-α2b plus entecavir led to a cccDNA decrease of 1.4 logs.Citation61 Collectively, these findings suggest that IFN may result in more significant reduction of intrahepatic cccDNA than NA in CHB patients.
Viral kinetics during antiviral therapy
Several previous studies investigating viral kinetics in CHB patients receiving antiviral treatment provided new mechanistic insights into the persistence of cccDNA. In brief, CHB patients receiving lamivudine reported a high rate of viral production up to 1011 virions/day. They also found that the half-life of a virion was approximately 1 day, but the half-life of producing cells varied, ranging from 10 to 100 days.Citation62 A further study in patients receiving adefovir revealed an interesting biphasic decline profile of viral kinetics, which included an initial faster phase and a second slower phase.Citation63 The former may reflect the clearance of HBV particles from plasma with a half-life of approximately 1 day, whereas the latter most likely represents the slow destruction of the infected hepatocytes, with a half-life of approximately 18 days. Based on the deduced rate of cccDNA reduction, it is estimated to take at least 14.6 years to eradicate persistent cccDNA. Even with the use of a potent antiviral agent, such as entecavir or tenofovir, treatment still fails to achieve complete clearance of CHB within a finite treatment duration. A biphasic decline pattern could also be observed in studying the viral kinetics in CHB patients treated with entecavir or a combination of antiviral agents, indicating that NAs have little to no effect on the second slow decline of virion-producing cccDNA-containing cells.Citation64,Citation65
EPIGENETIC REGULATION OF THE REPLICATIVE ABILITY OF HBV cccDNA
In contrast to the naked DNA of a prokaryotic genome, the eukaryotic genome is packaged into highly ordered chromatin, which contains a repeating unit, known as the nucleosome, composed of histones and approximately 200 bp of DNA. Each nucleosome contains two copies of each of the four core histones: H2A, H2B, H3 and H4. Epigenetic alterations, including DNA methylation at dinucleotide CpG sites and post-translational covalent histone modifications (i.e., acetylation, methylation, phosphorylation, etc.), can regulate chromatin accessibility and thus control eukaryotic gene transcription.Citation66,Citation67,Citation68 Previous studies found that episomal HBV cccDNA is associated with chromatin and non-chromatin proteins in the nuclei of infected hepatocytes. The cccDNA is organized into a typical beads-on-a-string nucleosomal DNA and exists as a minichromosome.Citation21,Citation69 The HBV core protein has also been shown to preferentially bind to HBV double-stranded DNA, leading to a reduction in the nucleosomal spacing of the HBV nucleoprotein complex by 10%, from 200 bp to 180 bp.Citation69,Citation70
Recently, epigenetic regulation, including histone modification and DNA methylation, has been shown to play a role in the regulation of the transcriptional activity of cccDNA.Citation71 By using a novel chromatin immunoprecipitation assay for cccDNA, Pollicino et al.Citation72 demonstrated that the acetylation status of the cccDNA-bound H3 and H4 histones regulates the transcriptional activity of cccDNA. Additionally, histone hypoacetylation and histone deacetylase 1 recruitment to the cccDNA in liver tissue are correlated with low HBV viremia in CHB patients. Using the same approach with a cccDNA-specific chromatin immunoprecipitation, Belloni et al.Citation73 suggested that several non-histone proteins, including histone acetyltransferases and deacetylases, bind to the cccDNA minichromosome. Furthermore, viral protein HBx regulates the cccDNA transcription by prevention of hypoacetylation of cccDNA, illustrating the function of a non-chromatin protein in the regulation of cccDNA transcription.Citation73
DNA methylation also occurs in cccDNA. Three CpG islands have been identified in the HBV genome.Citation74 Some variations have also been found in the cccDNA CpG islands in different genotypes of HBV.Citation75 Methylation of cccDNA reduces the expression of viral proteins.Citation76 Interestingly, the levels of cccDNA methylation in HBeAg-negative patients are higher than those in HBeAg-positive patients.Citation77 Consistent with this observation, the intrahepatic HBV cccDNA in HBeAg-negative patients exhibits a lower replication capacity compared to that in HBeAg-positive patients.Citation78,Citation79 This indicates not only a decreased quantity of cccDNA, but also an impairment in its quality in HBeAg-negative patients. In addition, IFN-α can inhibit HBV transcription by altering the epigenetic modification of cccDNA minichromosomes,Citation80,Citation81 suggesting antiviral treatment may suppress HBV replication through epigenetic regulation.
MECHANISMS FOR THE DESTRUCTION OF HBV cccDNA
Curing chronic HBV infection requires elimination of the persistent HBV cccDNA. Several mechanisms may be required to work cooperatively to achieve this goal: killing of the infected hepatocytes by the cytolytic immune response, primarily by cytotoxic CD8 T cells; non-cytolytic clearance of cccDNA by the inflammatory cytokines, i.e., IFN-γ and tumor necrosis factor (TNF)-α; and dilution of cccDNA by hepatocyte proliferation and replacement with regenerating uninfected cells.Citation8,Citation82
In HBV transgenic mice and HBV-infected chimpanzees, HBV replication can be suppressed without destruction of the HBV genome-containing hepatocytes, which is so-called non-cytolytic clearance of HBV.Citation83,Citation84,Citation85 In chimpanzees with acute HBV infection, the HBV DNA declines before the peak of alanine transaminase (ALT), supporting the notion of non-cytolytic suppression of HBV replication. This non-cytolytic control of viral replication is dependent on virus-specific CD8 T cells and is primarily mediated by the pro-inflammatory cytokines IFN-γ and TNF-α.Citation83,Citation85,Citation86 Additionally, IFN-α also contributes to non-cytolytic clearance in HBV transgenic mice.Citation87 Further investigation into the molecular mechanisms involved in non-cytolytic clearance of virus revealed that IFN-γ and TNF-α suppress viral replication by eliminating the HBV nucleocapsid particles inside viral genomes and destabilizing viral RNA.Citation88 However, similar non-cytolytic clearance of virus could not be demonstrated in WHV or DHBV animal models. Delivery of IFN-γ did not provide any extra benefit nor lead to further destruction of cccDNA.Citation89,Citation90 The causes of these inconsistent observations across model systems remain unclear.
Recently, it has been suggested that apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3 (APOBEC3)-family proteins form an innate defense mechanism that inhibits HBV replication. The previous study showed that APOBEC3G inhibits the packaging of pregenomic RNA in the HBV capsid and thus, promotes the clearance of HBV DNA in a non-cytolytic manner.Citation91 A recent study also discovered that APOBEC3A is induced by IFN following DNA detection. It deaminates foreign double-stranded DNA cytidines to uridines, which are then converted by the uracil DNA glycosylase UNG2 to abasic lesions, and results in foreign DNA degradation. In contrast, IFN-induced APOBEC3A does not exert effects on genomic DNA.Citation92 The APOBEC3G also exerts its hypermutation effect on the HBV cccDNA, which is counteracted by UNG because inhibition of UNG extensively enhances the G-to-A or C-to-T hypermutation in cccDNA.Citation93 A recent paper also described that IFN-α and lymphotoxin-β-receptor activation can upregulate APOBEC3A and 3B cytidine deaminases, respectively, in HBV-infected cell lines and primary hepatocytes. The HBV core protein mediates the interaction of APOBEC3A and 3B with nuclear cccDNA, ultimately resulting in cccDNA degradation.Citation59
Another important mechanism for viral clearance is killing of the infected hepatocytes by cytotoxic CD8 T cells. CD8 T cells have been recognized to be an important factor in control of HBV infection. Experiments involving depletion of CD8 T cells confirmed their role in cytolytic and non-cytolytic effects on viral clearance.Citation94 In ducks chronically infected with DHBV, hepatocyte turnover was required for rapid viral clearance during antiviral therapy.Citation95 In an investigation of the transient WHV infection in woodchucks, apoptosis of the infected cells and subsequent replacement by the regenerated uninfected cells were responsible for the recovery from transient WHV infection.Citation96 To further analyze hepatocyte turnover for viral clearance, Summers et al.Citation97 developed a quantitative assay for the integrated genome of WHV, which serves as a cell lineage marker, to determine the proportion of hepatocyte turnover. The results indicated that during the resolution of transient infections, a large fraction of the infected hepatocyte population was killed and replaced by hepatocyte cell division.
The same research group also studied chronically WHV-infected woodchucks receiving antiviral therapy with the nucleoside analog 1-(2-fluoro-5-methyl-beta-L-arabino-furanosyl)uracil (L-FMAU). They found that although viral cccDNA declined 20- to 100-fold, the frequency of the integrated WHV remained relatively constant over the course of treatment; this result argued for the notion that the uninfected hepatocytes are derived from the infected hepatocyte population.Citation98 They also found clonal expansion of hepatocytes during chronic WHV infection. Approximately half of the liver is derived from the high copy number (i.e., >1000 cells) clones, indicating that a high degree of hepatocyte proliferation and selection occurred during the chronic period of WHV infection.Citation99 Taken together, clearance of hepadnavirus cccDNA in persistent hepadnavirus-infected animals and in CHB patients appears to require a high proportion of hepatocyte turnover, indicating a critical role for killing of the infected hepatocytes and regeneration of uninfected hepatocytes derived from the infected hepatocyte population.
A NOVEL AGENT TARGETING HBV cccDNA OR cccDNA-CONTAINING HEPATOCYTES IS A POTENTIALLY PROMISING STRATEGY TO ERADICATE HBV INFECTION
As discussed above, current antiviral treatments fail to eradicate chronic HBV infection. An ideal strategy for curing chronic hepatitis B should specifically destroy HBV cccDNA or eliminate the infected hepatocytes without overt hepatotoxicity. A recent study showed that IFN-α and lymphotoxin-β-receptor activation can induce non-cytolytic destruction of cccDNA without hepatotoxicity by stimulating APOBEC3A and APOBEC3G, respectively.Citation59 Therefore, lymphotoxin-β-receptor activation may serve as a therapeutic alternative for destruction of cccDNA. Although this is an exciting observation, several critical issues have been raised, including the potential uncertainty of cccDNA quantification.Citation52 Further studies are required to confirm the conclusions. Another approach is to use the sequence-specific genome-editing DNA endonucleases, which bear the potential to specifically disrupt HBV cccDNA.Citation100 Previous studies have shown that zinc-finger nucleases and transcription activator-like effector nucleases can specifically destroy HBV cccDNA.Citation101,Citation102,Citation103 Our preliminary data also demonstrated that the CRISPR/Cas9 system, another genome-editing tool,Citation104 can also be utilized to specifically disrupt HBV cccDNA. However, delivery of these large systems to target the cccDNA-containing hepatocytes in vivo remains challenging.
In addition to specific disruption of cccDNA, an alternative strategy for curing chronic hepatitis B is to eliminate cccDNA-containing hepatocytes. A cytotoxic T-cell response is responsible for viral clearance and liver injury during HBV infection.Citation91 Patients who recover from acute HBV infection possess a polyclonal and multispecific T-cell response,Citation105,Citation106,Citation107 indicating the role of T cells in viral clearance. Protzer et al.Citation108,Citation109 have engineered T cells with chimeric receptors directed against HBV surface proteins; these proteins allow HBV-infected hepatocytes to be specifically targeted and destroyed in vitro and in vivo. The effects of all the aforementioned strategies, including the currently used IFN and NA, on persistent cccDNA are summarized in .
Table 1 Current antiviral treatments and potential strategies that may eliminate persistent cccDNA
FUTURE PERSPECTIVES
Persistence of cccDNA in HBV infection has significant clinical consequences. Previous studies have clearly demonstrated that persistent cccDNA is responsible for HBV reactivation in patients with resolved hepatitis B receiving immunosuppressives. HBV cccDNA is also the major barrier to eradicating chronic HBV infection. Despite the introduction of potent NAs, HBV still cannot be cured by NAs alone. Many patients with chronic hepatitis B require long-term or even life-long use of NAs. Understanding the molecular mechanisms regulating the synthesis and maintenance of cccDNA provides mechanistic insight into its stability and possible weak points. Novel drugs targeting the stable pool of cccDNA or cccDNA-containing hepatocytes may allow the realization of the hope of eliminating persistent cccDNA for the purpose of curing chronic HBV infection in a manner parallel to the clearance of hepatitis C virus infection.Citation110
We thank Ms Shin-Hwa Yu (Department of Internal Medicine, National Taiwan University Hospital, Taiwan, China) for her assistance in drawing the figure. This work was supported by National Health Research Institute (grant NHRI-EX103- 10235SC) to Hung-Chih Yang and by National Taiwan University (grant 103R7559-3) to Jia-Horng Kao.
- Kao JH, Chen PJ, Chen DS.Recent advances in the research of hepatitis B virus-related hepatocellular carcinoma: epidemiologic and molecular biological aspects. Adv Cancer Res2010;108: 21–72.
- Dienstag JL.Hepatitis B virus infection. N Engl J Med2008;359: 1486–1500.
- Lin CL, Kao JH.Hepatitis B viral factors and treatment responses in chronic hepatitis B. J Formosan Med Assoc2013;112: 302–311.
- Kao JH.HBeAg-positive chronic hepatitis B: why do I treat my patients with pegylated interferon? Liver Int2014;34 Suppl 1: 112–119.
- Ahn SH, Chan HL, Chen PJ et al.Chronic hepatitis B: whom to treat and for how long? Propositions, challenges, and future directions. Hepatol Int2010;4: 386–395.
- Liang Y, Jiang J, Su M et al.Predictors of relapse in chronic hepatitis B after discontinuation of anti-viral therapy. Aliment Pharmacol Ther2011;34: 344–352.
- Perez-Cameo C, Pons M, Esteban R.New therapeutic perspectives in HBV: when to stop NAs. Liver Int2014;34 Suppl 1: 146–153.
- Zoulim F.New insight on hepatitis B virus persistence from the study of intrahepatic viral cccDNA. J Hepatol2005;42: 302–308.
- Michalak TI, Pasquinelli C, Guilhot S, Chisari FV.Hepatitis B virus persistence after recovery from acute viral hepatitis. J Clin Invest1994;94: 907.
- Rehermann B, Ferrari C, Pasquinelli C, Chisari FV.The hepatitis B virus persists for decades after patients' recovery from acute viral hepatitis despite active maintenance of a cytotoxic T-lymphocyte response. Nat Med1996;2: 1104–1108.
- Loriot MA, Marcellin P, Walker F et al.Persistence of hepatitis B virus DNA in serum and liver from patients with chronic hepatitis B after loss of HBsAg. J Hepatol1997;27: 251–258.
- Mason AL, Xu L, Guo L, Kuhns M, Perrillo RP.Molecular basis for persistent hepatitis B virus infection in the liver after clearance of serum hepatitis B surface antigen. Hepatology1998;27: 1736–1742.
- Summers J, Mason WS.Replication of the genome of a hepatitis B-like virus by reverse transcription of an RNA intermediate. Cell1982;29: 403–415.
- Nassal M.Hepatitis B viruses: reverse transcription a different way. Virus Res2008;134: 235–249.
- Seeger C, Mason WS.Hepatitis B virus biology. Microbiol Mol Biol Rev2000;64: 51–68.
- Yan H, Zhong G, Xu G et al.Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife2012;1: e00049.
- Moolla N, Kew M, Arbuthnot P.Regulatory elements of hepatitis B virus transcription. J Viral Hepat2002;9: 323–331.
- Quasdorff M, Protzer U.Control of hepatitis B virus at the level of transcription. J Viral Hepat2010;17: 527–536.
- Tuttleman JS, Pourcel C, Summers J.Formation of the pool of covalently closed circular viral DNA in hepadnavirus-infected cells. Cell1986;47: 451–460.
- Wu TT, Coates L, Aldrich CE, Summers J, Mason WS.In hepatocytes infected with duck hepatitis B virus, the template for viral RNA synthesis is amplified by an intracellular pathway. Virology1990;175: 255–261.
- Newbold JE, Xin H, Tencza M et al.The covalently closed duplex form of the hepadnavirus genome exists in situ as a heterogeneous population of viral minichromosomes. J Virol1995;69: 3350–3357.
- Zhang YY, Zhang BH, Theele D, Litwin S, Toll E, Summers J.Single-cell analysis of covalently closed circular DNA copy numbers in a hepadnavirus-infected liver. Proc Natl Acad Sci USA2003;100: 12372–12377.
- Gao W, Hu J.Formation of hepatitis B virus covalently closed circular DNA: removal of genome-linked protein. J Virol2007;81: 6164–6174.
- Guo H, Jiang D, Zhou T, Cuconati A, Block TM, Guo JT.Characterization of the intracellular deproteinized relaxed circular DNA of hepatitis B virus: an intermediate of covalently closed circular DNA formation. J Virol2007;81: 12472–12484.
- Guo H, Mao R, Block TM, Guo JT.Production and function of the cytoplasmic deproteinized relaxed circular DNA of hepadnaviruses. J Virol2010;84: 387–396.
- Jones SA, Boregowda R, Spratt TE, Hu J.In vitro epsilon RNA-dependent protein priming activity of human hepatitis B virus polymerase. J Virol2012;86: 5134–5150.
- Werle-Lapostolle B, Bowden S, Locarnini S et al.Persistence of cccDNA during the natural history of chronic hepatitis B and decline during adefovir dipivoxil therapy. Gastroenterology2004;126: 1750–1758.
- Chu CM, Liaw YF.Hepatitis B surface antigen seroclearance during chronic HBV infection. Antivir Ther2010;15: 133–143.
- Marusawa H, Uemoto S, Hijikata M et al.Latent hepatitis B virus infection in healthy individuals with antibodies to hepatitis B core antigen. Hepatology2000;31: 488–495.
- Brechot C, Thiers V, Kremsdorf D, Nalpas B, Pol S, Paterlini-Brechot P.Persistent hepatitis B virus infection in subjects without hepatitis B surface antigen: clinically significant or purely “occult”? Hepatology2001;34: 194–203.
- Raimondo G, Pollicino T, Cacciola I, Squadrito G.Occult hepatitis B virus infection. J Hepatol2007;46: 160–170.
- Yeo W, Chan TC, Leung NW et al.Hepatitis B virus reactivation in lymphoma patients with prior resolved hepatitis B undergoing anticancer therapy with or without rituximab. J Clin Oncol2009;27: 605–611.
- Hsu C, Tsou HH, Lin SJ et al.Chemotherapy-induced hepatitis B reactivation in lymphoma patients with resolved HBV infection: a prospective study. Hepatology2014;59: 2092–2100.
- Raimondo G, Pollicino T, Romano L, Zanetti AR.A 2010 update on occult hepatitis B infection. Pathol Biol (Paris)2010;58: 254–257.
- Civitico GM, Locarnini SA.The half-life of duck hepatitis B virus supercoiled DNA in congenitally infected primary hepatocyte cultures. Virology1994;203: 81–89.
- Moraleda G, Saputelli J, Aldrich CE, Averett D, Condreay L, Mason WS.Lack of effect of antiviral therapy in nondividing hepatocyte cultures on the closed circular DNA of woodchuck hepatitis virus. J Virol1997;71: 9392–9399.
- Dandri M, Burda MR, Will H, Petersen J.Increased hepatocyte turnover and inhibition of woodchuck hepatitis B virus replication by adefovir in vitro do not lead to reduction of the closed circular DNA. Hepatology2000;32: 139–146.
- Hantz O, Parent R, Durantel D, Gripon P, Guguen-Guillouzo C, Zoulim F.Persistence of the hepatitis B virus covalently closed circular DNA in HepaRG human hepatocyte-like cells. J Gen Virol2009;90(Pt 1): 127–135.
- Zhu Y, Yamamoto T, Cullen J et al.Kinetics of hepadnavirus loss from the liver during inhibition of viral DNA synthesis. J Virol2001;75: 311–322.
- Addison WR, Walters KA, Wong WW et al.Half-life of the duck hepatitis B virus covalently closed circular DNA pool in vivo following inhibition of viral replication. J Virol2002;76: 6356–6363.
- Le Guerhier F, Pichoud C, Guerret S et al.Characterization of the antiviral effect of 2′,3′-dideoxy-2′, 3′-didehydro-beta-L-5-fluorocytidine in the duck hepatitis B virus infection model. Antimicrob Agents Chemother2000;44: 111–122.
- Le Guerhier F, Pichoud C, Jamard C et al.Antiviral activity of beta-L-2′,3′-dideoxy-2′,3′-didehydro-5-fluorocytidine in woodchucks chronically infected with woodchuck hepatitis virus. Antimicrob Agents Chemother2001;45: 1065–1077.
- Reaiche GY, Le Mire MF, Mason WS, Jilbert AR.The persistence in the liver of residual duck hepatitis B virus covalently closed circular DNA is not dependent upon new viral DNA synthesis. Virology2010;406: 286–292.
- Siliciano JD, Siliciano RF.A long-term latent reservoir for HIV-1: discovery and clinical implications. J Antimicrob Chemother2004;54: 6–9.
- Kock J, Schlicht HJ.Analysis of the earliest steps of hepadnavirus replication: genome repair after infectious entry into hepatocytes does not depend on viral polymerase activity. J Virol1993;67: 4867–4874.
- Delmas J, Schorr O, Jamard C et al.Inhibitory effect of adefovir on viral DNA synthesis and covalently closed circular DNA formation in duck hepatitis B virus-infected hepatocytes in vivo and in vitro. Antimicrob Agents Chemother2002;46: 425–433.
- Kock J, Baumert TF, Delaney WE 4th, Blum HE, von Weizsacker F.Inhibitory effect of adefovir and lamivudine on the initiation of hepatitis B virus infection in primary tupaia hepatocytes. Hepatology 200;38: 1410–1418.
- Boni C, Penna A, Bertoletti A et al.Transient restoration of anti-viral T cell responses induced by lamivudine therapy in chronic hepatitis B. J Hepatol2003;39: 595–605.
- Boni C, Laccabue D, Lampertico P et al.Restored function of HBV-specific T cells after long-term effective therapy with nucleos(t)ide analogues. Gastroenterology2012;143: 963–973.
- Sung JJ, Wong ML, Bowden S et al.Intrahepatic hepatitis B virus covalently closed circular DNA can be a predictor of sustained response to therapy. Gastroenterology2005;128: 1890–1897.
- Wong DK, Yuen MF, Yuan H et al.Quantitation of covalently closed circular hepatitis B virus DNA in chronic hepatitis B patients. Hepatology2004;40: 727–737.
- Chisari FV, Mason WS, Seeger C.Virology. Comment on “Specific and nonhepatotoxic degradation of nuclear hepatitis B virus cccDNA”. Science2014;344: 1237.
- Chan HL, Wong VW, Tse AM et al.Serum hepatitis B surface antigen quantitation can reflect hepatitis B virus in the liver and predict treatment response. Clin Gastroenterol Hepatol2007;5: 1462–1468.
- Tseng TC, Liu CJ, Su TH et al.Serum hepatitis B surface antigen levels predict surface antigen loss in hepatitis B e antigen seroconverters. Gastroenterology2011;141: 517–525.
- Tseng TC, Kao JH.Clinical utility of quantitative HBsAg in natural history and nucleos(t)ide analogue treatment of chronic hepatitis B: new trick of old dog. J Gastroenterol2013;48: 13–21.
- Wong DK, Yuen MF, Ngai VW, Fung J, Lai CL.One-year entecavir or lamivudine therapy results in reduction of hepatitis B virus intrahepatic covalently closed circular DNA levels. Antivir Ther2006;11: 909–916.
- Perrillo R.Benefits and risks of interferon therapy for hepatitis B. Hepatology2009;49(5 Suppl): S103–S111.
- Allweiss L, Volz T, Lutgehetmann M et al.Immune cell responses are not required to induce substantial hepatitis B virus antigen decline during pegylated interferon-alpha administration. J Hepatol2014;60: 500–507.
- Lucifora J, Xia Y, Reisinger F et al.Specific and nonhepatotoxic degradation of nuclear hepatitis B virus cccDNA. Science2014: 343: 1221–1228.
- Wursthorn K, Lutgehetmann M, Dandri M et al.Peginterferon alpha-2b plus adefovir induce strong cccDNA decline and HBsAg reduction in patients with chronic hepatitis B. Hepatology2006;44: 675–684.
- Hagiwara S, Kudo M, Osaki Y et al.Impact of peginterferon alpha-2b and entecavir hydrate combination therapy on persistent viral suppression in patients with chronic hepatitis B. J Med Virol2013;85: 987–995.
- Nowak MA, Bonhoeffer S, Hill AM, Boehme R, Thomas HC, McDade H.Viral dynamics in hepatitis B virus infection. Proc Natl Acad Sci USA1996;93: 4398–4402.
- Tsiang M, Rooney JF, Toole JJ, Gibbs CS.Biphasic clearance kinetics of hepatitis B virus from patients during adefovir dipivoxil therapy. Hepatology1999;29: 1863–1869.
- Lewin SR, Ribeiro RM, Walters T et al.Analysis of hepatitis B viral load decline under potent therapy: complex decay profiles observed. Hepatology2001;34: 1012–1020.
- Wolters LM, Hansen BE, Niesters HG, DeHertogh D, de Man RA.Viral dynamics during and after entecavir therapy in patients with chronic hepatitis B. J Hepatol2002;37: 137–144.
- Jenuwein T, Allis CD.Translating the histone code. Science2001;293: 1074–1080.
- Lande-Diner L, Zhang J, Ben-Porath I et al.Role of DNA methylation in stable gene repression. J Biol Chem2007;282: 12194–12200.
- Li B, Carey M, Workman JL.The role of chromatin during transcription. Cell2007;128: 707–719.
- Bock CT, Schranz P, Schroder CH, Zentgraf H.Hepatitis B virus genome is organized into nucleosomes in the nucleus of the infected cell. Virus genes1994;8: 215–229.
- Bock CT, Schwinn S, Locarnini S et al.Structural organization of the hepatitis B virus minichromosome. J Mol Biol2001;307: 183–196.
- Levrero M, Pollicino T, Petersen J, Belloni L, Raimondo G, Dandri M.Control of cccDNA function in hepatitis B virus infection. J Hepatol2009;51: 581–592.
- Pollicino T, Belloni L, Raffa G et al.Hepatitis B virus replication is regulated by the acetylation status of hepatitis B virus cccDNA-bound H3 and H4 histones. Gastroenterology2006;130: 823–837.
- Belloni L, Pollicino T, de Nicola F et al.Nuclear HBx binds the HBV minichromosome and modifies the epigenetic regulation of cccDNA function. Proc Natl Acad Sci USA2009;106: 19975–19979.
- Vivekanandan P, Thomas D, Torbenson M.Hepatitis B viral DNA is methylated in liver tissues. J Viral Hepat2008;15: 103–107.
- Zhang Y, Li C, Zhang Y et al.Comparative analysis of CpG islands among HBV genotypes. PloS One2013;8: e56711.
- Vivekanandan P, Thomas D, Torbenson M.Methylation regulates hepatitis B viral protein expression. J Infect Dis2009;199: 1286–1291.
- Guo Y, Li Y, Mu S, Zhang J, Yan Z.Evidence that methylation of hepatitis B virus covalently closed circular DNA in liver tissues of patients with chronic hepatitis B modulates HBV replication. J Med Virol2009;81: 1177–1183.
- Laras A, Koskinas J, Dimou E, Kostamena A, Hadziyannis SJ.Intrahepatic levels and replicative activity of covalently closed circular hepatitis B virus DNA in chronically infected patients. Hepatology2006;44: 694–702.
- Volz T, Lutgehetmann M, Wachtler P et al.Impaired intrahepatic hepatitis B virus productivity contributes to low viremia in most HBeAg-negative patients. Gastroenterology2007;133: 843–852.
- Belloni L, Allweiss L, Guerrieri F et al.IFN-alpha inhibits HBV transcription and replication in cell culture and in humanized mice by targeting the epigenetic regulation of the nuclear cccDNA minichromosome. J Clin Invest2012;122: 529–537.
- Liu F, Campagna M, Qi Y et al.Alpha-interferon suppresses hepadnavirus transcription by altering epigenetic modification of cccDNA minichromosomes. PLoS Pathog2013;9: e1003613.
- Dandri M, Petersen J.Hepatitis B virus cccDNA clearance: killing for curing? Hepatology2005;42: 1453–1455.
- Guidotti LG, Ando K, Hobbs MV et al.Cytotoxic T lymphocytes inhibit hepatitis B virus gene expression by a noncytolytic mechanism in transgenic mice. Proc Natl Acad Sci USA1994;91: 3764–3768.
- Guidotti LG.Viral clearance without destruction of infected cells during acute HBV infection. Science1999;284: 825–829.
- Guidotti LG, Chisari FV.Noncytolytic control of viral infections by the innate and adaptive immune response. Annu Rev Immunol2001;19: 65–91.
- Phillips S, Chokshi S, Riva A, Evans A, Williams R, Naoumov NV.CD8+ T cell control of hepatitis B virus replication: direct comparison between cytolytic and noncytolytic functions. J Immunol2010;184: 287–295.
- McClary H, Koch R, Chisari FV, Guidotti LG.Relative sensitivity of hepatitis B virus and other hepatotropic viruses to the antiviral effects of cytokines. J Virol2000;74: 2255–2264.
- Guidotti LG, Ishikawa T, Hobbs MV, Matzke B, Schreiber R, Chisari FV.Intracellular inactivation of the hepatitis B virus by cytotoxic T lymphocytes. Immunity1996;4: 25–36.
- Jacquard AC, Nassal M, Pichoud C et al.Effect of a combination of clevudine and emtricitabine with adenovirus-mediated delivery of gamma interferon in the woodchuck model of hepatitis B virus infection. Antimicrob Agents Chemother2004;48: 2683–2692.
- Zhu Y, Cullen JM, Aldrich CE et al.Adenovirus-based gene therapy during clevudine treatment of woodchucks chronically infected with woodchuck hepatitis virus. Virology2004;327: 26–40.
- Turelli P, Mangeat B, Jost S, Vianin S, Trono D.Inhibition of hepatitis B virus replication by APOBEC3G. Science2004;303: 1829.
- Stenglein MD, Burns MB, Li M, Lengyel J, Harris RS.APOBEC3 proteins mediate the clearance of foreign DNA from human cells. Nat Struct Mol Biol2010;17: 222–229.
- Kitamura K, Wang Z, Chowdhury S, Simadu M, Koura M, Muramatsu M.Uracil DNA glycosylase counteracts APOBEC3G-induced hypermutation of hepatitis B viral genomes: excision repair of covalently closed circular DNA. PLoS Pathog2013;9: e1003361.
- Thimme R, Wieland S, Steiger C et al.CD8+ T cells mediate viral clearance and disease pathogenesis during acute hepatitis B virus infection. J Virol2003;77: 68–76.
- Fourel I, Cullen JM, Saputelli J et al.Evidence that hepatocyte turnover is required for rapid clearance of duck hepatitis B virus during antiviral therapy of chronically infected ducks. J Virol1994;68: 8321–8330.
- Guo JT, Zhou H, Liu C et al.Apoptosis and regeneration of hepatocytes during recovery from transient hepadnavirus infections. J Virol2000;74: 1495–1505.
- Summers J, Jilbert AR, Yang W et al.Hepatocyte turnover during resolution of a transient hepadnaviral infection. Proc Natl Acad Sci USA2003;100: 11652–11659.
- Summers J, Mason WS.Residual integrated viral DNA after hepadnavirus clearance by nucleoside analog therapy. Proc Natl Acad Sci USA2004;101: 638–640.
- Mason WS, Jilbert AR, Summers J.Clonal expansion of hepatocytes during chronic woodchuck hepatitis virus infection. Proc Natl Acad Sci USA2005;102: 1139–1144.
- Schiffer JT, Swan DA, Stone D, Jerome KR.Predictors of hepatitis B cure using gene therapy to deliver DNA cleavage enzymes: a mathematical modeling approach. PLoS Comput Biol2013;9: e1003131.
- Cradick TJ, Keck K, Bradshaw S, Jamieson AC, McCaffrey AP.Zinc-finger nucleases as a novel therapeutic strategy for targeting hepatitis B virus DNAs. Mol Ther2010;18: 947–954.
- Bloom K, Ely A, Mussolino C, Cathomen T, Arbuthnot P.Inactivation of hepatitis B virus replication in cultured cells and in vivo with engineered transcription activator-like effector nucleases. Mol Ther2013;21: 1889–1897.
- Chen J, Zhang W, Lin J et al.An efficient antiviral strategy for targeting hepatitis B virus genome using Transcription Activator-Like Effector Nucleases. Mol Ther2013: 22: 303–311.
- Mali P, Esvelt KM, Church GM.Cas9 as a versatile tool for engineering biology. Nat Methods2013;10: 957–963.
- Penna A, Chisari FV, Bertoletti A et al.Cytotoxic T lymphocytes recognize an HLA-A2-restricted epitope within the hepatitis B virus nucleocapsid antigen. J Exp Med1991;174: 1565–1570.
- Nayersina R, Fowler P, Guilhot S et al.HLA A2 restricted cytotoxic T lymphocyte responses to multiple hepatitis B surface antigen epitopes during hepatitis B virus infection. J Immunol1993;150: 4659–4671.
- Rehermann B, Fowler P, Sidney J et al.The cytotoxic T lymphocyte response to multiple hepatitis B virus polymerase epitopes during and after acute viral hepatitis. J Exp Med1995;181: 1047–1058.
- Bohne F, Protzer U.Adoptive T-cell therapy as a therapeutic option for chronic hepatitis B. J Viral Hepat2007;14 Suppl 1: 45–50.
- Krebs K, Bottinger N, Huang LR et al.T cells expressing a chimeric antigen receptor that binds hepatitis B virus envelope proteins control virus replication in mice. Gastroenterology2013;145: 456–465.
- Schmidt WN, Nelson DR, Pawlotsky JM, Sherman KE, Thomas DL, Chung RT.Direct-acting antiviral agents and the path to interferon independence. Clin Gastroenterol Hepatol2014;12: 728–737.