
Abstract
A wide range of influenza A viruses of pigs and birds have infected humans in the last decade, sometimes with severe clinical consequences. Each of these so-called zoonotic infections provides an opportunity for virus adaptation to the new host. Fortunately, most of these human infections do not yield viruses with the ability of sustained human-to-human transmission. However, animal influenza viruses have acquired the ability of sustained transmission between humans to cause pandemics on rare occasions in the past, and therefore, influenza virus zoonoses continue to represent threats to public health. Numerous recent studies have shed new light on the mechanisms of adaptation and transmission of avian and swine influenza A viruses in mammals. In particular, several studies provided insights into the genetic and phenotypic traits of influenza A viruses that may determine airborne transmission. Here, we summarize recent studies on molecular determinants of virulence and adaptation of animal influenza A virus and discuss the phenotypic traits associated with airborne transmission of newly emerging influenza A viruses. Increased understanding of the determinants and mechanisms of virulence and transmission may aid in assessing the risks posed by animal influenza viruses to human health, and preparedness for such risks.
Introduction
Influenza A virus is a negative sense, single-stranded, segmented RNA virus of the Orthomyxoviridae family. Influenza A viruses are classified on the basis of the antigenic properties of the glycoproteins hemagglutinin (HA) and neuraminidase (NA) into 18 HA subtypes (H1–H18) and 11 NA subtypes (N1–N11).Citation1 The combination of specific HA and NA subtypes is used in influenza A virus nomenclature (e.g., H5N1, H7N9, H3N2). All influenza A virus subtypes are circulating in wild birds, which are therefore considered the natural influenza A virus reservoir, except H17, H18, N10 and N11. Viruses of subtypes H17N10 and H18N11 were recently identified in bat samples, raising the possibility that birds are not the exclusive influenza A virus reservoir.Citation1
INFLUENZA EPIDEMICS AND PANDEMICS
Influenza viruses cause annually recurrent epidemics, which result in approximately three to five million cases of severe illness and 250 000 to 500 000 deaths worldwide.Citation2 Infection with influenza viruses results in protective immunity, which is at least in part mediated by antibodies against the viral surface glycoproteins HA and NA. During circulation in humans, the virus gradually accumulates point mutations in HA and NA, which allows the virus to escape host immunity. This phenomenon, also known as antigenic drift, explains the occurrence of seasonal influenza epidemics.
The introduction of a new virus subtype that has not previously been circulating in the human population is referred to as antigenic shift. Antigenic shift occurs upon the introduction of an influenza A virus from the animal reservoir with or without reassortment, i.e., the mixing of genes from two (or more) animal and human influenza A viruses. While influenza viruses are continuously changing by antigenic drift, antigenic shift happens only infrequently. In the last century, four human influenza A virus pandemics have occurred, at least three of which resulted from reassortment between human and animal influenza A viruses.Citation3,Citation4,Citation5
Spanish H1N1 pandemic
The 1918 ‘Spanish’ influenza pandemic was caused by an H1N1 virus, and is known as the deadliest single event recorded in recent human history, killing as many as 50 million people worldwide.Citation3 The mortality rates were unusually high among young adults, a phenomenon that remains poorly understood to this day. Although its origin has not been fully resolved,Citation3,Citation6 data suggest that the 1918 virus evolved from an avian virus around 1910, with or without adaptation in an intermediate host.Citation7
Asian H2N2 pandemic
The ‘Asian’ pandemic H2N2 virus of 1957 was responsible for approximately two million deaths globally.Citation8 During this pandemic, predominantly very young and very old individuals were affected. The H2N2 virus emerged upon reassortment between human and avian influenza viruses. The HA, NA and basic polymerase 1 (PB1) genes originated from an avian H2N2 virus and the remaining gene segments from the H1N1 virus that circulated prior to 1957.Citation4
Hong Kong H3N2 pandemic
In 1968, the circulating H2N2 virus was replaced by the ‘Hong Kong’ H3N2 virus, which has continued to circulate in humans to date. During the pandemic of 1968, an estimated one million people were killed worldwide.Citation8 The pandemic H3N2 virus emerged upon reassortment between avian and human influenza viruses; this time the HA and PB1 segments of the H2N2 virus were replaced by those of an avian H3 virus.Citation4 This pandemic was mild compared to the earlier pandemics, possibly as a result of prior immunity in the human population against e.g. the N2 NA.
Re-emergence of H1N1
In 1977, an influenza outbreak was detected that affected predominantly young adults. This outbreak was caused by the reintroduction of an H1N1 virus also known as the ‘Russian’ H1N1 virus. This virus was genetically very similar to viruses circulating in the early 1950s and lacked years of genetic evolution, suggesting that the 1977 H1N1 virus was accidentally released from a laboratory.Citation9 The reintroduced H1N1 virus did not replace the circulating H3N2 virus and subsequently these virus subtypes cocirculated in humans until the ‘Russian’ H1N1 virus was replaced in 2009 by the pandemic H1N1 influenza virus.
2009 H1N1 pandemic
A H1N1 virus caused the first influenza pandemic in the 21st century and continues to be detected worldwide.Citation10 In the spring of 2009, the H1N1 virus (A(H1N1)pdm09) was first detected in Mexico, after which it spread around the world in only a few months. In terms of mortality, the A(H1N1)pdm09 virus was relatively mild, and preferentially affected young adults. People over 65 years of age experienced a relatively mild infection because of cross-protective immunity to the A(H1N1)pdm09 virus, as this virus was antigenically similar to H1N1 viruses circulating before 1957.Citation11 The A(H1N1)pdm09 virus was a reassortant virus that contained the NA and matrix (M) genes from the ‘Eurasian swine’ influenza virus lineage, while other genes originate from a ‘triple reassortant’ swine influenza virus that had previously acquired its genes upon reassortment between human, avian and (classical) swine influenza viruses.Citation12 The fact that the A(H1N1)pdm09 virus emerged after reassortment in swine supports the hypothesis that swine can serve as a mixing vessel for the generation of new human influenza viruses.
PANDEMIC THREATS
In the last century, influenza viruses of three HA subtypes (H1, H2 and H3) thus acquired the ability to be transmitted efficiently among humans. In addition, influenza viruses of the H5, H6, H7, H9 and H10 subtypes are also considered to represent pandemic threats since they have crossed the species barrier and infected humans (). Fortunately, these influenza viruses so far lacked the ability to spread efficiently between humans.
Table 1 Zoonotic influenza A viruses affecting humans
Avian influenza viruses in humans
Almost all influenza virus subtypes are present in the avian reservoir as low pathogenic avian influenza (LPAI) viruses. In this reservoir, consisting primarily of birds living near wetlands and aquatic environment such as the Anseriformes and Charadriiformes, avian influenza viruses reassort frequently.Citation35 LPAI viruses are able to spread across the world via bird migration. Domestic ducks and geese may serve as ‘bridge species’ for transmission to domestic poultry. Both wild and domestic birds are suitable influenza virus hosts that mostly experience asymptomatic infections, despite high virus replication. Viruses of subtypes H5 and H7 are a notable exception, since they can evolve in poultry to become highly pathogenic avian influenza (HPAI) viruses, causing severe disease and mortality.
In 1997, a large outbreak of HPAI H5N1 virus infections in poultry in Hong Kong resulted in the first documented cases of direct transmission of avian viruses from poultry to humans.Citation36 Since 2003, HPAI H5N1 viruses have spread throughout Asia, Europe and Africa, causing severe outbreaks in poultry. During these outbreaks, HPAI H5N1 viruses have been detected in various mammals on numerous occasions. As of October 7, 661 cases of human HPAI H5N1 virus infection have been reported to the World Health Organization, of which more than half were fatal.Citation13 While most of the human cases occurred upon direct contact with infected poultry, a few family clusters of human-to-human transmission of HPAI H5N1 virus have been documented.Citation37,Citation38,Citation39 Fortunately, the HPAI H5N1 virus lacks the ability to spread efficiently between humans, a requirement for influenza pandemics to occur. However, because the H5N1 virus is able to infect humans, it is feared that this virus may mutate or reassort with circulating human influenza viruses, possibly resulting in better adaptation to humans and subsequent human-to-human transmission.
At the end of March 2013, a novel LPAI H7N9 virus emerged in China, resulting in 137 confirmed human infections including 45 fatalities.Citation13 This virus caused only mild or no symptoms in birds, and as a consequence stayed undetected in poultry for some time. However, in humans this virus caused severe pneumonia and acute respiratory distress syndrome in a large number of cases.Citation20 The H7N9 virus emerged upon multiple reassortment events between different avian influenza viruses.Citation40 Even though this novel H7N9 virus harbors mammalian adaption markers that have been associated with increased replication and transmission in mammals, only one case of (non-sustainable) transmission between humans has been reported.Citation41 Several other H7 influenza virus outbreaks in poultry have resulted in human cases of infection in the past. On August 14 2013, HPAI H7N7 was reported in poultry in Northern Italy and in three human cases of infection.Citation13 In 2003, a large outbreak of an HPAI H7N7 virus in poultry in the Netherlands resulted in 89 cases of human infection, one of which was fatal.Citation19 H7 viruses often displayed an unusual tissue tropism; the virus targeted the conjunctiva, resulting in conjunctivitis, a symptom rarely reported for other influenza virus subtypes.Citation42
Sporadically, H9N2 avian influenza viruses have been responsible for human infections.Citation27 H9N2 viruses have become endemic in poultry populations throughout Eurasia. Furthermore, this subtype has been isolated from pigs, and numerous reassortment events between H9N2 virus and other influenza virus subtypes, like the novel H7N9 virus, have been reported.Citation43 H9N2 viruses with either avian and human or human receptor specificity are now prevalent in many Eurasian countries, thereby increasing the possibility of this virus to infect humans.Citation44
To date, only one human case of avian H6N1 virus infection has been reported.Citation30 Nevertheless, avian H6 viruses have the ability to replicate in mice and ferrets without prior adaptation.Citation45 In addition, the frequent reassortment and the high prevalence of H6 viruses in birds have raised concerns about H6 viruses.Citation46
In 1968, influenza viruses of the H2 subtype have disappeared from humans. Therefore, an increasing proportion of the current world population is likely to be susceptible to infection with H2 viruses if they would re-emerge. Influenza viruses of the H2 subtype continue to circulate in several avian species and pigs, posing a potential pandemic threat.Citation47,Citation48
In March 2010, an outbreak of LPAI H10N7 in chickens was responsible for the infection of abattoir workers. Reported outbreaks of H10N7 are uncommon, but these human cases emphasize that this avian influenza virus subtype is able to cross the species barrier.Citation31
Swine influenza viruses in humans
Influenza viruses with pandemic potential may arise in pigs, because pig cells express both human and avian influenza virus receptors, thus providing an opportunity for adaptation of avian influenza viruses, and reassortment with human viruses when they infect the same cell. When the A(H1N1)pdm09 virus became enzootic in swine, this virus continued to reassort with other swine influenza viruses. Human infections with H1N2 viruses demonstrated the continuous threat of reassortant swine viruses.Citation49 Starting in 2011, a novel H3N2 variant virus (H3N2v) containing seven gene segments of swine influenza virus origin and the A(H1N1)pdm09 M gene segment was detected in 348 humans.Citation50 Although humans may have cross-protective immunity due to infections with the seasonal H3N2 viruses that circulated in the 1990s,Citation51 the H3N2v is antigenically distinct from the currently circulating seasonal H3N2 viruses and can thus potentially infect a large proportion of the human population. Human infections with swine influenza viruses appear to occur infrequently. However, the continuous reassortment with other influenza viruses and waning cross-protective immunity in humans may eventually lead to emergence of viruses with the potential to spread among humans.
MOLECULAR BASIS OF INFLUENZA A VIRUS VIRULENCE IN MAMMALS
Influenza viruses from the animal reservoir can harbor genetic traits responsible for increased host range and virulence in mammals. When influenza viruses cross the species barrier and infect humans, additional genetic changes are frequently introduced. It is of importance to identify and investigate (adaptive) phenotypic and genetic traits associated with host range and virulence, to estimate the risks that animal influenza viruses pose for human health.
HA
The HA glycoprotein is composed of two subunits, the globular surface subunit HA1 and the stalk-like transmembrane subunit HA2. The HA protein of influenza viruses is initially synthesized as a single polypeptide precursor (HA0), which is cleaved into HA1 and HA2 subunits by cellular proteases. The sequence of the cleavage site determines by which cellular proteases HA0 can be cleaved. The highly pathogenic phenotype of some H5 and H7 viruses is the result of the introduction of a multibasic cleavage site (MBCS) in HA and directly correlates with the high lethality of avian influenza viruses in poultry, primarily as a consequence of systemic virus replication in these hosts. HPAI H5N1 virus can also cause severe and/or fatal disease in a wide range of mammalian species, but the association between HA cleavability and systemic virus replication is less straightforward for influenza viruses in mammals as compared to poultry. Removal of the MBCS from HA of an HPAI H5N1 virus resulted in a virus that caused only respiratory tract infection in mice, in contrast to the systemic replication of the HPAI H5N1 virus, indicating that the MBCS was a major virulence factor in mice.Citation52 However, experimental infection of non-human primates with HPAI H5N1 virus caused respiratory tract disease only with no evidence of systemic virus replication.Citation53 Recent studies showed that the MBCS in an HPAI H5N1 virus was important for systemic spread in ferrets, since deletion of the MBCS from HPAI H5N1 virus resulted in replication that was restricted to the respiratory tract.Citation54,Citation55 However, insertion of an MBCS in a human H3N2 virus did not result in systemic spread in ferrets.Citation56 The increased tissue tropism and virulence of HPAI H5N1 virus in mammals is most likely caused by multiple factors, among which the MBCS is essential. However, the presence of appropriate receptors for virus attachment, presence of the proper proteases in host cells and local innate immune responses also influence the ability of influenza virus to replicate systemically.
The influenza virus replication cycle is initiated when the HA protein binds to sialic acid (SA) receptors on the host cell. Human influenza viruses preferentially bind to α2,6-linked SA receptors (α2,6-SA) which are predominant on epithelial cells in the human upper respiratory tract (URT), whereas avian influenza viruses bind to α2,3-linked SA (α2,3-SA) receptors, which are abundantly present on epithelial cells in the intestinal tract of birds and in the lower respiratory tract of humans. Structural studies have shown that receptor specificity is primarily determined by the receptor-binding domain (RBD); amino-acid substitutions in the RBD influence virus host range and cell and tissue tropism, and consequently, also virulence. For the H2 and H3 virus subtypes, amino-acid substitutions Q226L and G228S in the RBD (all amino-acid numbering hereafter refers to numbering in H3 HA) were shown to change the binding preference from avian to human receptors. For the H1 virus subtype, substitutions E190D and D225G were important for the change in preference from avian to human receptors. For the A(H1N1)pdm09 virus, the D225G residue was associated with increased disease severity,Citation57 presumably because viruses with this substitution acquired dual receptor specificity for α2,3-SA and α2,6-SA.Citation58
In the HA of HPAI H5N1 viruses, mutations have been described that changed the binding preference from α2,3-SA to α2,6-SA.Citation59 The majority of these amino-acid substitutions was located in or near the RBD. Several H5N1 strains with enhanced affinity for human-type α2,6-SA receptors have been described in Indonesia.Citation60 In Egypt, new sublineages of HPAI H5N1 viruses have emerged in local bird populations that displayed enhanced α2,6-SA binding, but retained binding affinity for α2,3-SA and were associated with increased attachment to cells of the human lower respiratory tract.Citation61
Characterization of the receptor binding preference of the novel H7N9 virus revealed that this virus can bind to both avian and human receptors.Citation62 The affinity for human receptors is associated with the mammalian adaptation mutation Q226L. However, because not all H7N9 isolates possess this amino-acid substitution, other mutations may contribute to the human receptor preference as well.
The dual receptor binding of avian H5N1 and H7N9 viruses, as well as the 1918 H1N1 virus, may be responsible, at least in part, for increased severity of disease in humans. These influenza viruses were able to infect epithelial cells that line the lower respiratory tract, the bronchi, bronchioles and type II pneumocytes, which predominantly contain the avian type α2,3-SA receptors.Citation63 It is plausible that when avian H5N1 and H7N9 viruses acquire stronger affinity for α2,6-SA receptors and lose affinity for α2,3-SA, this would increase their pandemic potential, but at the same time, would decrease virulence.
In addition to substitutions in the RBD, changes in glycosylation patterns of HA can affect host range and virulence of influenza viruses. In H1N1 viruses, glycosylation sites were shown to play an important role in antigenicity, virulence and affinity for the receptor.Citation64,Citation65 Introduction of extra glycosylation sites resulted in increased virulence of the A(H1N1)pdm09 virus in mice.Citation65 Loss of an N-linked glycosylation site located near the RBD in HA of H5N1 virus (position 158–160), combined with acquisition of human receptor specificity, enhanced H5N1 virus binding to α2,6-SA.Citation66 In addition, deletion of this glycosylation site increased the virulence of an H5N1 virus in mice.Citation67
NA
The balance between the activities of the HA and NA surface glycoproteins is crucial for virus replication and transmission, since HA binds to SA and NA cleaves SA from the host cell surface.Citation68 Changes in either the binding or cleavage activity can affect virus replication. In 2003, an outbreak of HPAI H7N7 virus in poultry in the Netherlands resulted in the death of one person and 89 human cases of conjunctivitis. When the sequence of the virus obtained from the fatal case was compared to the sequence of a virus isolated from a patient with conjunctivitis, four amino-acid substitutions in the NA gene were identified.Citation19 These mutations were subsequently shown to contribute to increased NA activity, resulting in more efficient replication in mammalian cells, most likely by preventing the formation of virus aggregates.Citation69
Upon transmission of avian influenza viruses form waterfowl to domestic birds, deletions in the NA stalk region—which are associated with increased virulence in poultry—have been detected frequently.Citation70 It is not clear how such shortened NA stalk regions influences virulence, but early steps of the viral replication cycle (binding, immune evasion, prevention of apoptosis) may be involved in addition to tuning of the HA/NA balance. The shortened NA stalk region of H5N1 viruses was also shown to contribute to virulence in mammalian hosts.Citation71
Polymerase complex
The influenza virus polymerase proteins, and in particular basic polymerase 2 (PB2), have been shown to be important for virus adaptation to mammalian species. Interspecies transmission requires adaptation of the viral polymerase to importin-α, which is responsible for translocation of the viral ribonucleoprotein complex to the nucleus, since avian and human influenza viruses require different importin-α.Citation72 Several amino-acid substitutions (E627K, D701N and G590S/Q591R) have been shown to compensate for the lack of efficient polymerase activity of avian influenza viruses in mammalian cells.Citation73,Citation74 The E627K substitution is the best characterized mammalian adaptation mutation. This substitution is associated with increased virulence of human HPAI H5N1 virus isolates and was found in a fatal human case of infection with HPAI H7N7 virus.Citation52,Citation69 In addition, most human H7N9 virus isolates contained 627 K, or the alternative mammalian adaptive substitution D701N. The D701N substitution in PB2 was found to increase virulence, and to expand the host range of avian H5N1 virus to mammalian hosts in the absence of E627K.Citation75,Citation76 The D701N substitution caused enhancement of binding of PB2 to importin-α1 in mammalian cells, resulting in increased transport of PB2 into the nucleus.Citation77 In terms of virulence, it has been shown that E627K in PB2 is a critical virulence determinant for clade 2.3.4 H5N1 viruses, whereas D701N in PB2 and other unknown virulence determinants appear to be involved in the high pathogenicity of clade 1 H5N1 viruses.Citation78 The A(H1N1)pdm09 virus did not contain the mammalian adaptation residues 627 K and/or 701 N. When the substitutions E627K or D701N were introduced in A(H1N1)pdm09, no increase in virulence was observed.Citation79 The lack of these mammalian adaptation markers in PB2 was shown to be compensated by substitutions G590S/Q591R, which may affect interaction with viral and/or cellular factors and promote virus replication in mammals.Citation73 Other mutations in PB1, acidic polymerase (PA), nucleoprotein (NP) and nucleic export protein have been described that can compensate for the poor polymerase activity of avian influenza viruses in human cells.Citation80
PB1-F2
The PB1-F2 protein contributes to viral pathogenicity by inducing apoptosis of infected cells. The 1918 H1N1 virus and various isolates of HPAI H5N1 virus specifically have an N66S amino-acid substitution, which is partially responsible for high virulence.Citation81 The PB1-F2 N66S variant reduced the production of interferon (IFN).Citation82 It was demonstrated that the PB1-F2 protein contributed to virulence of pandemic strains and the HPAI H5N1 virus through enhancement of the lung inflammatory response, resulting in increased pathology.Citation83 However, in some influenza viruses, like the A(H1N1)pdm09 virus, a stop codon prevents the expression of PB1-F2, and restoring this function had minimal impact on virulence.Citation84 In addition, PB1-F2 was shown to promote virulence and increase secondary bacterial pneumonia.Citation85
PA-X
A newly identified protein encoded by gene segment 3, termed PA-X, modulates the host response by repressing cellular gene expression, i.e., host-cell shut off.Citation86 PA-X-deficient viruses caused enhanced disease in mice, which was related to an accelerated host response. Truncation of the PA-X protein appeared to be associated with influenza virus lineages circulating in particular reservoirs, demonstrating that there may be some species specificity to the evolution of PA-X.Citation87
Non-structural protein 1 (NS1)
NS1 has been studied extensively as a molecular determinant of virulence. One of the most important roles of NS1 is that of an IFN antagonist, which is accomplished by several mechanisms.Citation88 Influenza viruses lacking NS1 are only able to replicate in cells or mice that have a compromised IFN response.Citation89 H5N1 viruses, unlike other human, avian and swine influenza viruses, are relatively resistant to the antiviral effects of IFN, which results in increased levels of pro-inflammatory cytokine expression.Citation90 This effect can be modulated by a D92E amino-acid substitution, which is responsible for increased virulence of HPAI H5N1 in mice and facilitates virus replication in the presence of IFN.Citation91
The C-terminus of NS1 contains a PDZ ligand domain (of the X–S/T–X–V type) that was shown to be an important determinant of virulence.Citation92 When this domain of NS1 of 1918 H1N1 and H5N1 HPAI viruses was introduced in a mouse-adapted influenza virus, it was shown to increase virulence.Citation93 Recent HPAI H5N1 virus isolates lack these motifs, which argues against a role of this domain for virulence for these H5N1 viruses.
Several amino-acid substitutions in the NS1 of H5N1 virus were shown to increase NS1 binding to CPSF30 (the cleavage and polyadenylation specificity factor 30-Kd subunit, involved in cellular pre-mRNA processing), which was also associated with enhanced viral replication.Citation94 The A(H1N1)pdm09 virus was not able to block host gene expression in both human and swine cell lines,Citation95 which is partially due to mutations that block CPSF30 binding. Additionally, A(H1N1)pdm09 virus has a truncated NS1 protein with a 11 amino-acid deletion at its C-terminus, and therefore, lacks the PDZ-binding domain.Citation5 However, after restoring these functions in A(H1N1)pdm09 virus, they did not appear to have a significant effect on virulence of A(H1N1)pdm09 virus in various animal models.Citation95
More studies are required to identify determinants of efficient replication of animal influenza viruses in mammals. One starting point for such studies could be the genome sequences of human and avian influenza viruses, to identify species-specific amino acids. For example, 32 amino-acid residues in PB2, PA, NP, M1 and NS1 were identified to be different between human and avian viruses, of which 13 were conserved among 1918, 1957 and 1968 pandemic influenza viruses.Citation96 These host-specific markers can help us identify the minimal requirements for adaptation of avian viruses to mammals and may have prognostic and diagnostic values for improving pandemic preparedness.
MOLECULAR BASIS OF INFLUENZA A VIRUS TRANSMISSION IN MAMMALS
Several influenza virus subtypes from the animal reservoir have the ability to infect the human host but most of these are not further transmitted via aerosols or respiratory droplets (hereafter referred to as ‘airborne’ transmission) between humans. On rare occasions however, influenza pandemics may be the consequence of influenza virus zoonosis. It is important to understand the mechanisms by which influenza viruses emerge, adapt to the human host and acquire the ability of airborne transmission between humans and other mammals. Below, we discuss viral characteristics that were shown to be important for airborne transmission between mammals. Molecular signatures that may contribute to airborne transmission of H5 and H9 influenza virus subtypes are summarized in .
Table 2 Molecular signatures in avian influenza viruses (possibly) affecting airborne transmission between ferrets
HA
From previous pandemics, we know that only two substitutions in the HA RBD may be required to confer binding preference for receptors on cells of the human URT.Citation98 When previous pandemic viruses were evaluated for their receptor recognition, it was demonstrated that amino-acid substitutions in HA resulting in a human receptor binding preference were a prerequisite for cross-species transmission and human adaptation of avian influenza viruses.Citation99,Citation100 However, just changing the receptor specificity of HPAI H5N1 virus was insufficient to confer airborne transmission between ferrets.Citation14,Citation15,Citation101 A reassortant virus carrying the surface proteins of an avian H9N2 virus with the other genes derived from a human H3N2 virus was transmitted efficiently via the airborne route between ferrets. The airborne transmissible H9 HA protein contained the Q226L RBD substitution and two others; T189A and R192G.Citation29 Inefficient transmission between ferrets was observed for a virus containing a clade 2.2 H5N1 influenza virus HA with human receptor specificity, a human NA gene and six other gene segments from a clade 1 HPAI H5N1 virus.Citation97 This study elaborated on the complexity of genetic changes in influenza viruses required for adaptation and airborne transmission. Herfst et al.Citation15 demonstrated that the HPAI H5N1 virus A/Indonesia/5/05 (clade 2.1), harboring E627K in PB2 and Q226L and G228S in HA, was not transmitted via the airborne route between ferrets. However, when this virus was forced to adapt to replicate in cells of the mammalian URT by repeated passage in ferrets, this virus was transmitted. Besides the three substitutions that were introduced by reverse genetics, only few additional substitutions were observed in the virus genome, of which two in HA were consistently detected in airborne-transmissible virus isolates (H110Y and T160A). The receptor binding properties of this airborne-transmissible H5N1 virus HA were determined via structural and biophysical studies, which showed increased affinity for human receptors and decreased affinity for avian receptors.Citation102 Imai et al.Citation14 showed that similar changes in the HA that changed the receptor binding preference (N224K and Q226L) yielded an airborne-transmissible H5, but also here additional substitutions in HA were required. This virus contained the HPAI H5 HA of A/Vietnam/1203/04 (clade 1) and the remaining seven gene segments from a A(H1N1)pdm09 virus. The airborne virus had only four amino-acid substitutions in HA compared to the wild-type HA. Quantitative biophysical measurements demonstrated that this airborne-transmissible H5 HA acquired specificity for human over avian receptors, comparable to previous pandemic strains.Citation103,Citation104 The Q226L amino-acid substitution contributed substantially to the human receptor binding preference and to the marked decrease in avian receptor binding. The studies on the airborne-transmissible H5 viruses enabled structural analyses of the HA RBD from these viruses, and consequently, highlighted the features necessary for human receptor binding.
An N-linked glycosylation site in HA was also shown to play a role in airborne transmission of H5N1 viruses. This glycosylation site is located near the RBD at position 158–160, and deletion of this site was demonstrated to be beneficial for human receptor binding affinity. Loss of this glycosylation site was found to be critical for H5N1 virus virulence in mice Citation105 and H5N1 virus transmissibility in a guinea pig direct contact model.Citation106 In the airborne H5 viruses of Herfst et al.Citation14 and Imai et al.,Citation15 the same glycosylation site was deleted through different substitutions, suggesting that the loss of this glycosylation site was critical for airborne transmission between mammals. Many H5N1 viruses, in particular within the H5N1 lineage circulating in Egypt (clade 2.2), lost their putative N-linked glycosylation site in nature. Recent analysis suggested that avian H5N1 viruses from Egypt lacking the glycosylation site transmit more readily from poultry to humans than those that possess the glycosylation site.Citation107 These viruses, fortunately, did not cause human-to-human transmission, implying that these viruses also require additional mutations.
Recent publications on the novel H7N9 virus demonstrated limited airborne transmission of this virus between ferrets.Citation22,Citation23,Citation24,Citation25,Citation108 Most human virus isolates harbor the Q226L substitution responsible for binding to the URT in mammals. However, this amino-acid substitution on its own may not be sufficient for efficient airborne transmission between ferrets. It is important to note that it is difficult to obtain quantitative data on airborne transmission in ferrets to compare efficiency of transmission between human and avian viruses, in part due to animal ethics considerations in relation to the required group size.Citation109 This difficulty is further complicated by differences in animal breed, age, gender, set-up of the airborne-transmission experiments and climatic conditions during the experiments between different research groups, which may all affect transmission efficiency.
Refining the human receptor binding preference of an avian influenza virus thus appeared to be necessary for viruses to become airborne. Recently, it was suggested that the stability of HA in an acidic environment might be an important additional requirement for airborne transmission between mammals. Upon influenza virus attachment and endocytosis, a low endosomal pH triggers a conformational change in the HA protein to expose the fusion peptide and trigger a fusion reaction between the viral and endosomal membranes, after which viral ribonucleoproteins are released in the cytoplasm. Phenotype analysis in the study of Imai et al.Citation14 demonstrated the requirement of a T318I amino-acid substitution in HA to lower the pH of membrane fusion, thus compensating for a decrease in HA stability caused by human receptor binding mutations in the airborne-transmissible H5. In addition, this mutation increased the thermostability of the virus. It has been shown that viruses which fuse at low pH have a higher thermostability than those fusing at a higher pH.Citation110 These environmental factors like temperature and pH are likely to affect airborne transmission. One of the substitutions consistently present in airborne-transmissible A/Indonesia/5/05 virus, H110Y, was likely to be involved in HA stability as well.Citation15,Citation102 Several other amino-acid substitutions are known to decrease the pH threshold for activation of HA, and could thus determine a similar phenotypic trait. To investigate whether mutations that alter the pH threshold for fusion (Y23H, H24Q, K58I) affected the fitness of H5N1 virus, mice were infected with wild-type and mutant H5N1 viruses A/chicken/Vietnam/C58/04 (clade 1). Mutant K58I—with the lowest threshold for fusion of pH 5.4—demonstrated increased replication and virulence, suggesting that adaptation of avian H5N1 influenza virus for replication in mice is supported by a low pH threshold for fusion.Citation111 The same substitution was responsible for increased replication of H5N1 A/Vietnam/1203/2004 (clade 1) in the URT of ferrets. In the absence of changes in receptor binding preference, this mutation did not alter the transmissibility, since neither the wild-type virus, nor the K58I mutant virus, resulted in direct contact transmission between ferrets.Citation112 The pH threshold for fusion is thus likely to be an important determinant for airborne transmission between mammals only in the context of a virus with appropriate human receptor binding preference. Comparison of the airborne-transmissible H5N1 viruses to the novel H7N9 virus may suggest that a lower pH threshold for fusion may be required for increased airborne transmissibility of the latter virus between mammals.Citation22 Additional research is necessary to identify substitutions in the H7N9 virus that are responsible for HA stabilization, which may confer enhanced transmissibility between mammals. Identification of amino-acid substitutions that decrease the pH threshold of fusion and increase the thermostability is important for influenza virus surveillance and risk assessment of currently circulating avian H5 and H7 influenza viruses. However, it is important to note that the increased acid stability and thermostability of HA may only be a surrogate marker for another phenotype, such as stability in aerosols or respiratory droplets or other conditions in the environment. Nevertheless, transmission via the airborne route requires the HA protein to be stable in the hosts’ environment.
Polymerase complex
As described above, the adaptation of the viral RNA polymerase is crucial for avian influenza viruses, since these polymerases have relatively poor activity in mammalian cells. Several amino-acid substitutions in the ribonucleoprotein complex are known to contribute to the host-range restriction of influenza viruses and increase replication in the mammalian URT. High virus replication in the URT is critical for airborne transmission since decreased replication abolished airborne transmission between mammals.Citation113 In the study of Herfst et al.,Citation15 the airborne-transmissible H5N1 virus possessed the E627K amino-acid substitution in PB2. However, it has yet to be elucidated if this residue was crucial for airborne transmission and whether it can be substituted by functionally equivalent mutations. The airborne-transmissible H5N1 viruses of Herfst et al. possessed several additional substitutions in the polymerase genes, which may have contributed to airborne transmission. A K627E substitution was found to reduce transmission in a direct contact transmission model for HPAI H5N1 virus. Introduction of D701N in combination with the K627E substitution resulted in more efficient transmission in mammals, a phenotype more similar to that of the wildtype viruses, suggesting that this residue can compensate for the absence of 627K.Citation75 In the context of a human H3N2 virus, introduction of K627E resulted in less efficient transmission in guinea pigs.Citation75 Studies on the A(H1N1)pdm09 virus demonstrated that this virus was transmitted efficiently via the airborne route between ferrets.Citation114 Nevertheless, acquisition of the D701N substitution in A(H1N1)pdm09 virus resulted in more severe disease and increased transmission.Citation115 These data indicate that amino-acid positions 627 and 701 in PB2 are critical determinants of airborne transmission between mammals in diverse virus backgrounds.
Reassortment
Previously, it was believed that avian influenza viruses may not cause the next influenza pandemic directly, as pigs were considered a necessary intermediate host to facilitate reassortment between avian, swine and human influenza viruses. In addition, pigs were thought to serve as a critical host for initial mammalian adaptation. However, the role of pigs as mixing vessel for avian and human influenza viruses may not be unique, as humans can be infected with avian influenza viruses directly from an avian reservoir. Reassortment of the H5N1 virus with contemporary human influenza viruses did not easily yield an airborne-transmissible virus between mammals.Citation116,Citation117,Citation118 However, more recently, it was shown that several reassortant H5N1 virus variants were transmitted via the airborne route between guinea pigs.Citation16 This H5 virus, which revealed dual specificity for avian and human receptors, required the PA and NS genes of the A(H1N1)pdm09 virus to acquire airborne transmissibility between guinea pigs. In addition, the NP, M and NA genes from A(H1N1)pdm09 virus were important to enhance airborne transmissibility between mammals. These results appear to be in contrast to the study of Imai et al.,Citation14 in which a reassortant virus with a mutant H5 HA with human receptor specificity in a A(H1N1)pdm09 virus backbone failed to transmit via the airborne route between ferrets. This difference may be explained by the use of different lineages of H5 HAs. In addition, it is unclear how the guinea pig and ferret models compare with respect to H5 virus transmission studies. However, as mentioned above, the avian-human reassortant influenza virus in the study of Imai et al. transmitted via the airborne route after passaging in ferrets, when a few additional genetic changes in HA were acquired. These two studies thus demonstrated that different airborne-transmissible H5N1 strains can emerge upon reassortment between human and avian influenza viruses, as was previously also shown for avian H9N2 virus.
Fortunately, at present, reassortment of avian H5, H7 and H9 viruses with contemporary human influenza viruses has not been detected in nature. This may in part be explained by the relatively low number of human cases of avian influenza virus infections, and the low probability of double infections with human influenza viruses in a single host. In addition, influenza viruses have to infect the same cell, and similar timing of infection is necessary to initiate gene reassortment. However, co-infections of avian and human influenza viruses in humans or pigs may provide new opportunities for reassortment, and evidence from pandemics of the last century indicates that the likelihood of such reassortment is not negligible.
The emergence of the A(H1N1)pdm09 virus demonstrated that pigs can be the direct source of pandemic influenza. Reassortment was responsible for the unique constellation of virus genes, which was shown to be a critical determinant of airborne transmission between ferrets,Citation114 in particular the Eurasian swine-origin gene segments.Citation119 The emergence of H1N2 and H3N2v swine viruses underlines that swine populations should be monitored closely for the emergence of influenza viruses with pandemic potential. When the airborne transmissibility of these viruses was assessed, the H1N2 virus acquired substitutions in HA (D222G) and NA (S315N) during a single ferret passage that resulted in airborne transmission among ferrets. Reverse genetics studies further indicated that these amino-acid substitutions contributed substantially to the airborne transmissibility.Citation34 The viral RNA polymerase was also shown to promote airborne transmission.Citation120 Moreover, four H3N2v viruses, collected from 2009 until 2011, also possessed the capacity to spread between cohoused ferrets, and the 2010 and 2011 H3N2v viruses transmitted efficiently to naive ferrets via the airborne route.Citation33 These findings support the continuous threat of swine influenza viruses to humans, and the need for continued surveillance.
Unknown determinants of transmission
The role of other viral proteins in airborne transmission has so far remained elusive. The study of Zhang et al.Citation16 demonstrated that the PA, NS, M, NA and NP genes are important to confer airborne transmission; however, the key amino-acid substitutions remain unknown. In addition, it has been suggested that H5N1 viruses that acquire a long stalk NA through reassortment may be more likely to be transmitted among humans.Citation121
Although it is known that influenza virus morphology affects virus production, the relative contribution of spherical and more elongated versions of influenza virus to transmission is still unclear. Early work showed that most influenza virus strains isolated from humans are predominantly filamentous and, upon continued passage in egg or tissue culture, adopt a more spherical morphology, which correlated with increased virus titers.Citation122 It is possible that increased levels of influenza virus production by spherical strains will results in more efficient influenza virus transmission.Citation119 The contribution of substitutions in the M gene segment on virus morphology and transmission warrants further investigation.
FURTHER THOUGHTS ON AIRBORNE TRANSMISSION OF INFLUENZA VIRUSES
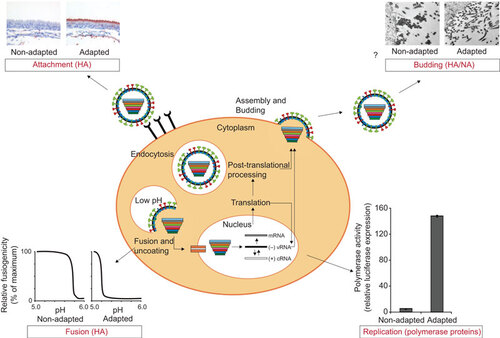
The influenza virus traits that are important for airborne transmission are slowly becoming apparent (Figure ): (i) attachment to and replication in appropriate cells of the mammalian airways, which is facilitated by the receptor binding preference of HA; (ii) increased binding via the deletion of a potential N-glycosylation site in HA; (iii) (acid) stabilization of HA; (iv) increased viral replication induced by the viral polymerase genes; and (v) virus shedding as single particles rather than aggregates, which may be facilitated by amino-acid substitutions in the HA and NA proteins.Citation69
Figure 1 Traits important for airborne transmission of influenza virus between mammals. Increased binding of HA to appropriate cells of the URT of mammalian host cells contributes to the airborne transmissibility. Increased acid stability of the HA was also observed in airborne transmissible virus. Adaptation of the polymerase complex is necessary to facilitate increased replication in the mammalian host cell and is a likely determinant of airborne transmissibility. Although this has not (yet) been shown directly, the release of single viral particles instead of aggregates may aid the airborne transmissibility of the virus, which may be regulated by the HA–NA balance. Virus attachment and budding images were described previously.Citation123 Fusion and replication data are unpublished. The fusion picture is hypothetical and shows a non-adapted HA with a high pH threshold for fusion and an adapted HA with a decreased pH threshold for fusion. The replication figure contains data of the Indonesia/5/05 polymerase complex and Indonesia/5/05 airborne transmissible polymerase complex by Herfst et al.Citation15
The studies on transmission of H9N2, H5N1, H7N9 and other influenza viruses are of importance to elucidate common mechanisms of transmission of influenza viruses. The results of such work may ultimately help to better assess the risk posed by the current HPAI H5N1 and H7N9 virus epizootics for human health. Increased fundamental understanding of the contribution of specific mutations or reassortment events and their associated biological traits to transmission is urgently needed. It is important to study common biological properties of transmissible viruses rather than genetic changes in a singular virus strain, to generalize the mechanisms leading to increased transmission. Data on the biological properties of the transmissible H5N1 virus may be extrapolated to the H7N9 virus and future zoonotic viruses, in order to assess the risk of a new pandemic and to implement appropriate control measures on time. Mechanistic studies on the phenotypic traits associated with each of the identified amino-acid substitutions from previous pandemic virus would be very informative.
Since it is impossible to predict which influenza viruses will preferentially emerge and may cause the next pandemic, it is key to continuously monitor the genetic composition of circulating influenza strains in animals and to assess their infection and transmission potential in animal models. Since multiple evolutionary trajectories could yield viruses transmissible via the airborne route between mammals, monitoring the genetic composition of virus isolates alone may be misleading and phenotypic assays should be added to surveillance studies to evaluate the threats posed by animal influenza viruses. Since virus isolates consist of quasi-species, monitoring plaque purified viruses or viruses generated from plasmid clones has to be considered for phenotypic assays. Information that reveals genetic changes which could possibly alter the virulence and transmission potential may help to better assess the risks of potential emerging influenza viruses. In addition, intervention strategies and ‘stamping-out’ procedures can be prioritized and targeted better. Whether a next pandemic will emerge only after reassortment is questionable, since airborne transmission of a wholly avian virus between mammals has now been shown to be possible for both laboratory-generated and natural avian influenza viruses.
The authors thank Dr Sander Herfst and Dr Mathilde Richard from the Department of Viroscience, Erasmus MC Rotterdam for helpful discussions. The authors are supported by NWO VICI grant 91896613, EU FP7 programs ANTIGONE and EMPERIE, and NIAID/NIH contract HHSN266200700010C.
- Tong S, Zhu X, Li Y et al.New world bats harbor diverse influenza a viruses. PLoS Pathog2013;9: e1003657.
- World Health Organization. Influenza (seasonal).Fact sheet No. 211.Geneva: WHO, 2009.Available at http://www.who.int/mediacentre/factsheets/fs211/en/index.html (accessed 9 December 2013).
- Taubenberger JK, Morens DM.1918 Influenza: the mother of all pandemics. Emerg Infect Dis2006;12: 15–22.
- Scholtissek C, Rohde W, von Hoyningen V, Rott R.On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology1978;87: 13–20.
- Neumann G, Noda T, Kawaoka Y.Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature2009;459: 931–939.
- Smith GJ, Bahl J, Vijaykrishna D et al.Dating the emergence of pandemic influenza viruses. Proc Natl Acad Sci USA2009;106: 11709–11712.
- Qi L, Davis AS, Jagger BW et al.Analysis by single-gene reassortment demonstrates that the 1918 influenza virus is functionally compatible with a low-pathogenicity avian influenza virus in mice. J Virol2012;86: 9211–9220.
- Kilbourne ED.Influenza pandemics of the 20th century. Emerg Infect Dis2006;12: 9–14.
- Nakajima K, Desselberger U, Palese P.Recent human influenza A (H1N1) viruses are closely related genetically to strains isolated in 1950. Nature1978;274: 334–339.
- Dawood FS, Jain S, Finelli L et al.Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med2009;360: 2605–2615.
- Hancock K, Veguilla V, Lu X et al.Cross-reactive antibody responses to the 2009 pandemic H1N1 influenza virus. N Engl J Med2009;361: 1945–1952.
- Garten RJ, Davis CT, Russell CA et al.Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science2009;325: 197–201.
- World Health Organization. Influenza at the human–animal interface.Geneva: WHO, 2013.Available at http://www.who.int/influenza/human_animal_interface/Influenza_Summary_IRA_HA_interface_7October13.pdf (accessed 7 October 2013).
- Imai M, Watanabe T, Hatta M et al.Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature2012;486: 420–428.
- Herfst S, Schrauwen EJ, Linster M et al.Airborne transmission of influenza A/H5N1 virus between ferrets. Science2012;336: 1534–1541.
- Zhang Y, Zhang Q, Kong H et al.H5N1 hybrid viruses bearing 2009/H1N1 virus genes transmit in guinea pigs by respiratory droplet. Science2013;340: 1459–1463.
- Ostrowsky B, Huang A, Terry W et al.Low pathogenic avian influenza A (H7N2) virus infection in immunocompromised adult, New York, USA, 2003. Emerg Infect Dis2012;18: 1128–1131.
- Tweed SA, Skowronski DM, David ST et al.Human illness from avian influenza H7N3, British Columbia. Emerg Infect Dis2004;10: 2196–2199.
- Fouchier RA, Schneeberger PM, Rozendaal FW et al.Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc Natl Acad Sci USA2004;101: 1356–1361.
- Gao R, Cao B, Hu Y et al.Human infection with a novel avian-origin influenza A (H7N9) virus. N Engl J Med2013;368: 1888–1897.
- Centers for Disease Prevention.Notes from the field: Highly pathogenic avian influenza A (H7N3) virus infection in two poultry workers—Jalisco, Mexico, July 2012. MMWR Morb Mortal Wkly Rep2012;61: 726–727.
- Richard M, Schrauwen EJ, de Graaf M et al.Limited airborne transmission of H7N9 influenza A virus between ferrets. Nature2013;501: 560–563.
- Zhu H, Wang D, Kelvin DJ et al.Infectivity, transmission, and pathology of human H7N9 influenza in ferrets and pigs. Science2013;341: 183–186.
- Watanabe T, Kiso M, Fukuyama S et al.Characterization of H7N9 influenza A viruses isolated from humans. Nature2013;501: 551–555.
- Belser JA, Gustin KM, Pearce MB et al.Pathogenesis and transmission of avian influenza A (H7N9) virus in ferrets and mice. Nature2013;501: 556–559.
- Butt KM, Smith GJ, Chen H et al.Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. J Clin Microbiol2005;43: 5760–5767.
- Peiris M, Yuen KY, Leung CW et al.Human infection with influenza H9N2. Lancet1999;354: 916–917.
- Cheng VC, Chan JF, Wen X et al.Infection of immunocompromised patients by avian H9N2 influenza A virus. J Infect2011;62: 394–399.
- Sorrell EM, Wan H, Araya Y, Song H, Perez DR.Minimal molecular constraints for respiratory droplet transmission of an avian-human H9N2 influenza A virus. Proc Natl Acad Sci USA2009;106: 7565–7570.
- Yuan J, Zhang L, Kan X et al.Origin and molecular characteristics of a novel 2013 avian influenza A H6N1 virus causing human infection in Taiwan. Clin Infect Dis2013;57: 1367–1368.
- Arzey GG, Kirkland PD, Arzey KE et al.Influenza virus A (H10N7) in chickens and poultry abattoir workers, Australia. Emerg Infect Dis2012;18: 814–816.
- Centers for Disease Control and Prevention. Reported infections with variant influenza viruses in the United States since 2005.Atlanta, GA: CDC, 2013.Available at http://www.cdc.gov/flu/swineflu/variant-cases-us.htm (accessed 5 November 2013).
- Pearce MB, Jayaraman A, Pappas C et al.Pathogenesis and transmission of swine origin A(H3N2)v influenza viruses in ferrets. Proc Natl Acad Sci USA2012;109: 3944–3949.
- Pascua PN, Song MS, Lee JH et al.Virulence and transmissibility of H1N2 influenza virus in ferrets imply the continuing threat of triple-reassortant swine viruses. Proc Natl Acad Sci USA2012;109: 15900–15905.
- Hatchette TF, Walker D, Johnson C, Baker A, Pryor SP, Webster RG.Influenza A viruses in feral Canadian ducks: extensive reassortment in nature. J Gen Virol2004;85(Pt 8): 2327–2337.
- de Jong JC, Claas EC, Osterhaus AD, Webster RG, Lim WL.A pandemic warning? Nature1997;389: 554.
- Kandun IN, Wibisono H, Sedyaningsih ER et al.Three Indonesian clusters of H5N1 virus infection in 2005. N Engl J Med2006;355: 2186–2194.
- Wang H, Feng Z, Shu Y et al.Probable limited person-to-person transmission of highly pathogenic avian influenza A (H5N1) virus in China. Lancet2008;371: 1427–1434.
- Ungchusak K, Auewarakul P, Dowell SF et al.Probable person-to-person transmission of avian influenza A (H5N1). N Engl J Med2005;352: 333–340.
- Lam TT, Wang J, Shen Y et al.The genesis and source of the H7N9 influenza viruses causing human infections in China. Nature2013;502: 241–244.
- Qi X, Qian YH, Bao CJ et al.Probable person to person transmission of novel avian influenza A (H7N9) virus in Eastern China, 2013: epidemiological investigation. BMJ2013;347: f4752.
- de Wit E, Fouchier RA.Emerging influenza. J Clin Virol2008;41: 1–6.
- Liu D, Shi W, Shi Y et al.Origin and diversity of novel avian influenza A H7N9 viruses causing human infection: phylogenetic, structural, and coalescent analyses. Lancet2013;381: 1926–1932.
- Matrosovich MN, Krauss S, Webster RG.H9N2 influenza A viruses from poultry in Asia have human virus-like receptor specificity. Virology2001;281: 156–162.
- Gillim-Ross L, Santos C, Chen Z et al.Avian influenza h6 viruses productively infect and cause illness in mice and ferrets. J Virol2008;82: 10854–10863.
- Munster VJ, Baas C, Lexmond P et al.Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog2007;3: e61.
- Krauss S, Obert CA, Franks J et al.Influenza in migratory birds and evidence of limited intercontinental virus exchange. PLoS Pathog2007;3: e167.
- Ma W, Vincent AL, Gramer MR et al.Identification of H2N3 influenza A viruses from swine in the United States. Proc Natl Acad Sci USA2007;104: 20949–20954.
- Centers for Disease Control and Prevention. H1N2 variant virus detected in Minnesota. Atlanta, GA: CDC, 2012.Available at http://www.cdc.gov/flu/spotlights/h1n2v-cases-mn.htm (accessed 7 September 2012).
- Nelson MI, Vincent AL, Kitikoon P, Holmes EC, Gramer MR.Evolution of novel reassortant A/H3N2 influenza viruses in North American swine and humans, 2009–2011. J Virol2012;86: 8872–8878.
- Shu B, Garten R, Emery S et al.Genetic analysis and antigenic characterization of swine origin influenza viruses isolated from humans in the United States, 1990–2010. Virology2012;422: 151–160.
- Hatta M, Gao P, Halfmann P, Kawaoka Y.Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science2001;293: 1840–1842.
- Rimmelzwaan GF, Kuiken T, van Amerongen G, Bestebroer TM, Fouchier RA, Osterhaus AD.Pathogenesis of influenza A (H5N1) virus infection in a primate model. J Virol2001;75: 6687–6691.
- Schrauwen EJ, Herfst S, Leijten LM et al.The multibasic cleavage site in H5N1 virus is critical for systemic spread along the olfactory and hematogenous routes in ferrets. J Virol2012;86: 3975–3984.
- Suguitan AL Jr, Matsuoka Y, Lau YF et al.The multibasic cleavage site of the hemagglutinin of highly pathogenic A/Vietnam/1203/2004 (H5N1) avian influenza virus acts as a virulence factor in a host-specific manner in mammals. J Virol2012;86: 2706–2714.
- Schrauwen EJ, Bestebroer TM, Munster VJ et al.Insertion of a multibasic cleavage site in the haemagglutinin of human influenza H3N2 virus does not increase pathogenicity in ferrets. J Gen Virol2011;92(Pt 6): 1410–1415.
- Kilander A, Rykkvin R, Dudman SG, Hungnes O.Observed association between the HA1 mutation D222G in the 2009 pandemic influenza A(H1N1) virus and severe clinical outcome, Norway 2009–2010. Euro Surveill2010;15: 19498.
- Chutinimitkul S, Herfst S, Steel J et al.Virulence-associated substitution D222G in the hemagglutinin of 2009 pandemic influenza A(H1N1) virus affects receptor binding. J Virol2010 Nov;84: 11802–11813.
- Chutinimitkul S, van Riel D, Munster VJ et al.In vitro assessment of attachment pattern and replication efficiency of H5N1 influenza A viruses with altered receptor specificity. J Virol2010;84: 6825–6833.
- Yamada S, Suzuki Y, Suzuki T et al.Haemagglutinin mutations responsible for the binding of H5N1 influenza A viruses to human-type receptors. Nature2006;444: 378–382.
- Watanabe Y, Ibrahim MS, Ellakany HF et al.Acquisition of human-type receptor binding specificity by new H5N1 influenza virus sublineages during their emergence in birds in Egypt. PLoS Pathog2011;7: e1002068.
- van Riel D, Leijten LM, de Graaf M et al.Novel avian-origin influenza A (H7N9) virus attaches to epithelium in both upper and lower respiratory tract of humans. Am J Pathol2013;183: 1137–1143.
- Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y.Avian flu: influenza virus receptors in the human airway. Nature2006;440: 435–436.
- Sun X, Jayaraman A, Maniprasad P et al.N-linked glycosylation of the hemagglutinin protein influences virulence and antigenicity of the 1918 pandemic and seasonal H1N1 influenza A viruses. J Virol2013;87: 8756–8766.
- Zhang Y, Zhu J, Li Y et al.Glycosylation on hemagglutinin affects the virulence and pathogenicity of pandemic H1N1/2009 influenza A virus in mice. PLoS ONE2013;8: e61397.
- Wang W, Lu B, Zhou H et al.Glycosylation at 158N of the hemagglutinin protein and receptor binding specificity synergistically affect the antigenicity and immunogenicity of a live attenuated H5N1 A/Vietnam/1203/2004 vaccine virus in ferrets. J Virol2010;84: 6570–6577.
- Chen H, Bright RA, Subbarao K et al.Polygenic virulence factors involved in pathogenesis of 1997 Hong Kong H5N1 influenza viruses in mice. Virus Res2007;128: 159–163.
- Wagner R, Matrosovich M, Klenk HD.Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev Med Virol2002;12: 159–166.
- de Wit E, Munster VJ, van Riel D et al.Molecular determinants of adaptation of highly pathogenic avian influenza H7N7 viruses to efficient replication in the human host. J Virol2010;84: 1597–1606.
- Munier S, Larcher T, Cormier-Aline F et al.A genetically engineered waterfowl influenza virus with a deletion in the stalk of the neuraminidase has increased virulence for chickens. J Virol2010;84: 940–952.
- Matsuoka Y, Swayne DE, Thomas C et al.Neuraminidase stalk length and additional glycosylation of the hemagglutinin influence the virulence of influenza H5N1 viruses for mice. J Virol2009;83: 4704–4708.
- Gabriel G, Klingel K, Otte A et al.Differential use of importin-alpha isoforms governs cell tropism and host adaptation of influenza virus. Nat Commun2011;2: 156.
- Mehle A, Doudna JA.Adaptive strategies of the influenza virus polymerase for replication in humans. Proc Natl Acad Sci USA2009;106: 21312–21316.
- Subbarao EK, London W, Murphy BR.A single amino acid in the PB2 gene of influenza A virus is a determinant of host range. J Virol1993;67: 1761–1764.
- Steel J, Lowen AC, Mubareka S, Palese P.Transmission of influenza virus in a mammalian host is increased by PB2 amino acids 627K or 627E/701N. PLoS Pathog2009;5: e1000252.
- Li Z, Chen H, Jiao P et al.Molecular basis of replication of duck H5N1 influenza viruses in a mammalian mouse model. J Virol2005;79: 12058–12064.
- Gabriel G, Herwig A, Klenk HD.Interaction of polymerase subunit PB2 and NP with importin alpha1 is a determinant of host range of influenza A virus. PLoS Pathog2008;4: e11.
- Le QM, Ito M, Muramoto Y et al.Pathogenicity of highly pathogenic avian H5N1 influenza A viruses isolated from humans between 2003 and 2008 in northern Vietnam. J Gen Virol2010;91(Pt 10): 2485–2490.
- Herfst S, Chutinimitkul S, Ye J et al.Introduction of virulence markers in PB2 of pandemic swine-origin influenza virus does not result in enhanced virulence or transmission. J Virol2010;84: 3752–3758.
- Manz B, Schwemmle M, Brunotte L.Adaptation of avian influenza a virus polymerase in mammals to overcome the host species barrier. J Virol2013;87: 7200–7209.
- Conenello GM, Zamarin D, Perrone LA, Tumpey T, Palese P.A single mutation in the PB1-F2 of H5N1 (HK/97) and 1918 influenza A viruses contributes to increased virulence. PLoS Pathog2007;3: 1414–1421.
- Varga ZT, Ramos I, Hai R et al.The influenza virus protein PB1-F2 inhibits the induction of type I interferon at the level of the MAVS adaptor protein. PLoS Pathog2011;7: e1002067.
- McAuley JL, Chipuk JE, Boyd KL, van de Velde N, Green DR, McCullers JA.PB1-F2 proteins from H5N1 and 20 century pandemic influenza viruses cause immunopathology. PLoS Pathog2010;6: e1001014.
- Hai R, Schmolke M, Varga ZT et al.PB1-F2 expression by the 2009 pandemic H1N1 influenza virus has minimal impact on virulence in animal models. J Virol2010;84: 4442–4450.
- McAuley JL, Hornung F, Boyd KL et al.Expression of the 1918 influenza A virus PB1-F2 enhances the pathogenesis of viral and secondary bacterial pneumonia. Cell Host Microbe2007;2: 240–249.
- Jagger BW, Wise HM, Kash JC et al.An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science2012;337: 199–204.
- Shi M, Jagger BW, Wise HM, Digard P, Holmes EC, Taubenberger JK.Evolutionary conservation of the PA-X open reading frame in segment 3 of influenza A virus. J Virol2012;86: 12411–12413.
- Garcia-Sastre A.Induction and evasion of type I interferon responses by influenza viruses. Virus Res2011;162: 12–18.
- Garcia-Sastre A, Egorov A, Matassov D et al.Influenza A virus lacking the NS1 gene replicates in interferon-deficient systems. Virology1998;252: 324–330.
- Peiris JS, Yu WC, Leung CW et al.Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet2004;363: 617–619.
- Seo SH, Hoffmann E, Webster RG.Lethal H5N1 influenza viruses escape host anti-viral cytokine responses. Nat Med2002;8: 950–954.
- Obenauer JC, Denson J, Mehta PK et al.Large-scale sequence analysis of avian influenza isolates. Science2006;311: 1576–1580.
- Jackson D, Hossain MJ, Hickman D, Perez DR, Lamb RA.A new influenza virus virulence determinant: the NS1 protein four C-terminal residues modulate pathogenicity. Proc Natl Acad Sci USA2008;105: 4381–4386.
- Twu KY, Kuo RL, Marklund J, Krug RM.The H5N1 influenza virus NS genes selected after 1998 enhance virus replication in mammalian cells. J Virol2007;81: 8112–8121.
- Hale BG, Steel J, Medina RA et al.Inefficient control of host gene expression by the 2009 pandemic H1N1 influenza A virus NS1 protein. J Virol2010;84: 6909–6922.
- Finkelstein DB, Mukatira S, Mehta PK et al.Persistent host markers in pandemic and H5N1 influenza viruses. J Virol2007;81: 10292–10299.
- Chen LM, Blixt O, Stevens J et al.In vitro evolution of H5N1 avian influenza virus toward human-type receptor specificity. Virology2012;422: 105–113.
- Matrosovich M, Tuzikov A, Bovin N et al.Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J Virol2000;74: 8502–8512.
- Pappas C, Viswanathan K, Chandrasekaran A et al.Receptor specificity and transmission of H2N2 subtype viruses isolated from the pandemic of 1957. PLoS ONE2010;5: e11158.doi: https://doi.org/10.1371/journal.pone.0011158.
- Tumpey TM, Maines TR, van Hoeven N et al.A two-amino acid change in the hemagglutinin of the 1918 influenza virus abolishes transmission. Science2007;315: 655–659.
- Maines TR, Chen LM, van Hoeven N et al.Effect of receptor binding domain mutations on receptor binding and transmissibility of avian influenza H5N1 viruses. Virology2011;413: 139–147.
- Zhang W, Shi Y, Lu X, Shu Y, Qi J, Gao GF.An airborne transmissible avian influenza H5 hemagglutinin seen at the atomic level. Science2013;340: 1463–1467.
- Lu X, Shi Y, Zhang W, Zhang Y, Qi J, Gao GF.Structure and receptor-binding properties of an airborne transmissible avian influenza A virus hemagglutinin H5 (VN1203mut). Protein Cell2013;4: 502–511.
- Xiong X, Coombs PJ, Martin SR et al.Receptor binding by a ferret-transmissible H5 avian influenza virus. Nature2013;497: 392–396.
- Rudneva IA, Ilyushina NA, Timofeeva TA, Webster RG, Kaverin NV.Restoration of virulence of escape mutants of H5 and H9 influenza viruses by their readaptation to mice. J Gen Virol2005;86(Pt 10): 2831–2838.
- Gao Y, Zhang Y, Shinya K et al.Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog2009;5: e1000709.
- Neumann G, Macken CA, Karasin AI, Fouchier RA, Kawaoka Y.Egyptian H5N1 influenza viruses-cause for concern? PLoS Pathog2012;8: e1002932.
- Zhang Q, Shi J, Deng G et al.H7N9 influenza viruses are transmissible in ferrets by respiratory droplet. Science2013;341: 410–414.
- Belser JA, Maines TR, Katz JM, Tumpey TM.Considerations regarding appropriate sample size for conducting ferret transmission experiments. Future Microbiol2013;8: 961–965.
- Ruigrok RW, Martin SR, Wharton SA, Skehel JJ, Bayley PM, Wiley DC.Conformational changes in the hemagglutinin of influenza virus which accompany heat-induced fusion of virus with liposomes. Virology1986;155: 484–497.
- Zaraket H, Bridges OA, Russell CJ.The pH of activation of the hemagglutinin protein regulates H5N1 influenza virus replication and pathogenesis in mice. J Virol2013;87: 4826–4834.
- Zaraket H, Bridges OA, Duan S et al.Increased acid stability of the hemagglutinin protein enhances H5N1 influenza virus growth in the upper respiratory tract but is insufficient for transmission in ferrets. J Virol2013;87: 9911–9922.
- Lowen AC, Steel J, Mubareka S, Carnero E, Garcia-Sastre A, Palese P.Blocking interhost transmission of influenza virus by vaccination in the guinea pig model. J Virol2009;83: 2803–2818.
- Munster VJ, de Wit E, van den Brand JM et al.Pathogenesis and transmission of swine-origin 2009 A(H1N1) influenza virus in ferrets. Science2009;325: 481–483.
- Zhou B, Pearce MB, Li Y et al.Asparagine substitution at PB2 residue 701 enhances the replication, pathogenicity, and transmission of the 2009 pandemic H1N1 influenza A virus. PLoS ONE2013;8: e67616.
- Schrauwen EJ, Bestebroer TM, Rimmelzwaan GF, Osterhaus AD, Fouchier RA, Herfst S.Reassortment between Avian H5N1 and human influenza viruses is mainly restricted to the matrix and neuraminidase gene segments. PLoS ONE2013;8: e59889.
- Jackson S, van Hoeven N, Chen LM et al.Reassortment between avian H5N1 and human H3N2 influenza viruses in ferrets: a public health risk assessment. J Virol2009;83: 8131–8140.
- Maines TR, Chen LM, Matsuoka Y et al.Lack of transmission of H5N1 avian-human reassortant influenza viruses in a ferret model. Proc Natl Acad Sci USA2006;103: 12121–12126.
- Lakdawala SS, Lamirande EW, Suguitan AL Jr et al.Eurasian-origin gene segments contribute to the transmissibility, aerosol release, and morphology of the 2009 pandemic H1N1 influenza virus. PLoS Pathog2011;7: e1002443.
- Pascua PN, Song MS, Kwon HI et al.The homologous tripartite viral RNA polymerase of A/swine/Korea/CT1204/2009(H1N2) influenza virus synergistically drives efficient replication and promotes respiratory-droplet transmission in ferrets. J Virol2013;87: 10552–10562.
- Blumenkrantz D, Roberts KL, Shelton H, Lycett S, Barclay WS.The short stalk length of highly pathogenic avian influenza H5N1 virus neuraminidase limits transmission of pandemic H1N1 virus in ferrets. J Virol2013;87: 10539–10551.
- Kilbourne ED, Murphy JS.Genetic studies of influenza viruses. I. Viral morphology and growth capacity as exchangeable genetic traits. Rapid in ovo adaptation of early passage Asian strain isolates by combination with PR8. J Exp Med1960;111: 387–406.
- Sorrell E, Schrauwen E, Linster M, de Graaf M, Herfst S, Fouchier R.Predicting ‘airborne’ influenza viruses: (trans-) mission impossible? Curr Opin Virol2011;1: 635–642.