
Abstract
Monoclonal antibodies (mAbs), which currently constitute the main class of biotherapeutics, are now recognized as major medical tools that are increasingly being considered to fight severe viral infections. Indeed, the number of antiviral mAbs developed in recent years has grown exponentially. Although their direct effects on viral blunting have been studied in detail, their potential immunomodulatory actions have been overlooked until recently. The ability of antiviral mAbs to modulate antiviral immune responses in infected organisms has recently been revealed. More specifically, upon recognition of their cognate antigens, mAbs form immune complexes (ICs) that can be recognized by the Fc receptors expressed on different immune cells of infected individuals. This binding may be followed by the modulation of the host immune responses. Harnessing this immunomodulatory property may facilitate improvements in the therapeutic potential of antiviral mAbs. This review focuses on the role of ICs formed with different viral determinants and mAbs in the induction of antiviral immune responses in the context of both passive immunotherapies and vaccination strategies. Potential deleterious effects of ICs on the host immune response are also discussed.
THERAPEUTIC POTENTIAL OF ANTIVIRAL MONOCLONAL ANTIBODIES
Monoclonal antibodies (mAbs) have gained an important place in the therapeutic arsenal against severe human diseases. More than 50 mAbs have been approved or are under review for human use, and several hundred are currently being tested in the clinic,Citation1, Citation2 most of them to treat patients suffering from a variety of cancers or inflammatory diseases.
Concerning antiviral mAbs, only one, directed against respiratory syncytial virus (RSV), has been approved for the prophylactic treatment of pediatric infections. However, employing mAbs as antiviral drugs is under consideration for the treatment of several chronic and acute severe viral infections, especially to address the public health emergencies such as the recent Ebola virus and Middle East respiratory syndrome coronavirus outbreaks.Citation3, Citation4, Citation5, Citation6, Citation7 Illustrating this trend, the number of antiviral mAbs developed and tested in preclinical and clinical trials has grown exponentially in the past 10 years and includes mAbs directed against life-threatening agents, such as human immunodeficiency virus (HIV), hepatitis B virus (HBV), hepatitis C virus (HCV), influenza virus, dengue virus, Ebola virus and severe acute respiratory syndrome virus coronavirus, among others.Citation8, Citation9, Citation10, Citation11, Citation12, Citation13, Citation14, Citation15, Citation16, Citation17, Citation18, Citation19, Citation20, Citation21, Citation22 Importantly, recent clinical data have also demonstrated the efficacy of anti-HIV mAbs in controlling viremia, when administered to HIV-infected patients, lending strong support to the idea that mAbs could broaden the therapeutic arsenal against severe viral infections.Citation23, Citation24 Their use as antiviral agents is all the more likely to be considered given that multiple biological activities may account for their therapeutic effects.
Although a few mAbs have been developed to inhibit the recognition of viral receptors or co-receptors on the surface of target cells, most antiviral mAbs have been selected for their ability to neutralize virions through the binding of their antigen-binding (Fab) fragment to viral surface antigens essential for entry into host cells. However, the biological activity of antibodies is also mediated by the fragment crystallizable region (Fc) moiety. Thus, it is interesting to note that most antiviral mAbs in use are immunoglobulin (Ig)-Gs displaying a variety of effector functions, including binding to both complement and Fcγ receptors (FcγRs). Different types of FcγRs are expressed in a regulated manner by many cells of the immune system, including B cells, natural killer cells, dendritic cells (DCs), monocytes/macrophages, granulocytes and mast cells, and their engagement by the Fc antibody moiety is essential for regulating the antibody effector functions.Citation25, Citation26 Upon recognition of their target antigens, antiviral mAbs can facilitate virus elimination via two types of complement-mediated mechanisms: (i) inactivation of viral particles and/or phagocytosis of opsonized virus mediated by cells of the innate immune system () and (ii) opsonization and subsequent lysis of infected cells, when viral antigens are also expressed on the cell surface (for example, envelope (Env) glycoprotein of lentiviruses such as HIV) via complement-dependent cytotoxicity. In addition to complement-mediated actions, recognition of FcγRs can entail antibody-dependent cellular phagocytosis and antibody-dependent cell-mediated cytotoxicity ().Citation20, Citation27, Citation28, Citation29, Citation30 Finally, antiviral mAbs also have a role in viral blunting by inhibiting cell-to-cell viral transmission.Citation31
Figure 1 Multiple Fc-mediated activities of antiviral mAbs (monoclonal antibodies). Antiviral mAbs can opsonize viruses, as well as infected cells in situations when viral antigens are also expressed on their surface (A). This can lead to viral clearance through various immune-mediated mechanisms. The Fc domain allows the binding of complement to free virions, inducing direct virolysis (B). Fcγ- and complement receptors can recognize opsonized virions, leading to their phagocytosis by cells of the innate immune system (C). Infected cells can also be eliminated by complement-dependent cytotoxicity (CDC), antibody-dependent cellular phagocytosis (ADCP) and/or antibody-dependent cell-mediated cytotoxicity (ADCC) mediated by innate immunity effector cells expressing FcγRs (D–F). Immune complexes (ICs) made with mAbs and different viral determinants (virions or infected cells) can be recognized by FcγRs expressed on antigen-presenting cells such as dendritic cells (DCs) (G). IC recognition by DCs subsequently leads to enhanced antigen uptake and presentation, allowing the induction of stronger antiviral immune responses.
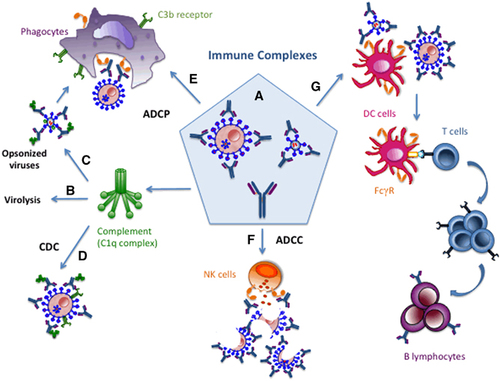
In addition to controlling the viral propagation by these mechanisms, the opsonization of viral particles and/or infected cells by therapeutic antiviral mAbs of the IgG type leads to the formation of immune complexes (ICs) recognizable by the FcγRs expressed on antigen-presenting cells (APCs) such as DCs. This can potentially affect the endogenous antiviral adaptive immune response of passive immunotherapy-treated individuals. Despite the fact that the immunoregulatory functions of antibodies (as well as ICs) have been known for a long time, and have been reported in different experimental settings and physiopathological situations,Citation25, Citation32, Citation33, Citation34, Citation35, Citation36, Citation37, Citation38 the immunomodulatory role of mAbs with clinical potential as antiviral drugs has only recently been considered.
This review mainly focuses on the induction of antiviral immune responses by ICs in both passive immunotherapies and vaccination strategies. The potential deleterious effects of antiviral antibodies on the host immune dysfunction and/or viral propagation are also discussed.
ANTIVIRAL MABS CAN ENHANCE THE HOST ANTIVIRAL IMMUNE RESPONSE IN AN FC-DEPENDENT MANNER
Only recently have studies addressed whether and how passive immunotherapies utilizing antiviral mAbs are able to enhance the antiviral immunity in infected individuals. This is largely due to the limited availability of suitable immunocompetent animal models of viral infection that allow in-depth investigations of the endogenous immune response. The concept that passive immunotherapies utilizing antiviral mAbs can induce long-term protective immunity has recently been established using an immunocompetent mouse model, consisting of short immunotherapies given to young animals infected with the FrCasE murine leukemia virus. The induction of such ‘vaccine-like’ effects by antiviral mAbs, as well as some of the mechanisms involved, are reviewed in detail elsewhere.Citation39 In brief, the inoculation of mouse pups with FrCasE is fatal because the antiviral immune response is too weak to control the viral propagation. In contrast, treatment with a neutralizing mAb for several days shortly after infection blunts viral propagation and induces a lifelong protective antiviral immunity composed of both a highly neutralizing humoral response and a cytotoxic CD8+ T-cell response.Citation40, Citation41, Citation42, Citation43, Citation44, Citation45 This induction of protective immunity strictly depends on the Fc fragment of the neutralizing mAbs.Citation43, Citation44 Moreover, the formation of ICs composed of the administered mAbs and infected cells rather than virions is crucial for the enhanced antiviral immune response.Citation43 Such ICs are recognized by the FcγRs expressed by DCs, which facilitate ICs internalization and lead to stronger activation and more efficient antigen presentation by DCs, eventually leading to stronger cytotoxic T-lymphocyte (CTL) responses. An Fc-mediated effect that occurs concurrently is the inhibition of regulatory T-cell (Treg) expansion. This depends on the mAb effector functionsCitation45 and occurs rapidly. Moreover, it is necessary for the development of the protective humoral and cellular responses, as Treg-mediated immunosuppression is observed in all cases of chronic viral infections, where it dampens antiviral immune responses, thereby permitting the establishment of chronicity. Finally, breastfeeding and placental transfer of maternal anti-FrCasE Igs induced by mAb immunotherapy not only led to the viral propagation blunting in infected pups, but also to the induction of long-lasting protective humoral immunity in these animals.Citation42 This is a particularly interesting observation when one considers that the FrCasE model is reminiscent of perinatal infection by HIV, including breastfeeding-mediated mother-to-child virus transmission.
Other evidence for the induction of ‘vaccine-like’ effects by antiviral mAbs comes from studies in several preclinical models of human viral infections and from HIV-infected patients.
In a mouse model of RSV infection, the administration of a neutralizing mAb directed against the virus attachment protein G induced a shift in the adaptive immune response from Th2- to Th1-type, leading to sustained and enhanced humoral and CD8+ T-cell responses.Citation46 However, this effect was not Fc-dependent, but rather due to the ability of the therapeutic mAb to counteract the intrinsic immunosuppressive activity of the RSV G protein.
mAb-driven enhancement of the humoral response has also been reported in two preclinical models of henipavirus infection in African green monkeys.Citation47, Citation48 Recovery from both Hendra and Nipah virus-induced disease correlated with the development of host antibody responses consequent to the administration of the highly neutralizing 102.4 mAb. This Hendra and Nipah virus cross-reactive mAb is currently being considered for human use.
Finally, anti-HIV antibodies can modulate immune responses in infected organisms. Such effects were initially reported in several non-human primate models of HIV infection and then observed in infected humans. Macaques were infected with different strains of simian immunodeficiency virus (SIV) or simian HIV (SHIV, a chimeric virus in which HIV Env substitutes for that of SIV and allows for the assessment of the antiviral effects of anti-HIV antibodies) following different protocols. These experiments showed that the administration of highly neutralizing antibodies (either mAbs or polyclonal Igs) enhanced both the humoral and cellular antiviral immune responses of treated animals.Citation8, Citation49, Citation50, Citation51 Interestingly, recent clinical data describe the elicitation of host humoral responses in viremic subjects upon single injection of the potent 3BNC117 anti-HIV mAb.Citation52 However, the mechanisms leading to the stimulation of antiviral immune responses in these preclinical models of HIV infection or in infected patients remain uncharacterized. Moreover, it is unknown whether these antiviral responses have genuine protective vaccine-like effects. In any case, these important observations open new avenues for the improvement of mAb-based antiviral HIV therapies. Moreover, as the in vivo activity of anti-HIV-1 bNAbs, including viral load control, was recently shown to crucially depend on Fc effector functions,Citation53, Citation54 an important issue is identifying that Fc–FcγRs interactions are involved in the induction of vaccine-like effects by antiviral mAbs.
ICS ENHANCE DC ACTIVATION AND INDUCE STRONGER ANTIVIRAL T-CELL RESPONSES: EVIDENCE FROM IN VITRO STUDIES
To understand the mechanisms underlying the enhancement of antiviral responses by ICs, several in vitro studies have addressed whether antibody-mediated viral uptake by DCs could lead to stronger activation of these cells and the development of stronger virus-specific CD4+ and CD8+ T-cell responses in an Fc-dependent manner. Such an increase in the cellular immune response has been reported in different infectious settings using ICs made with different types of antigens, including recombinant viral proteins and whole virions, as well as infected cells ().
Table 1 In vitro studies of T-cell responses modulation by IC-activated DC
Concerning ICs made with viral proteins, several reports have shown that ICs made up of anti-HBV mAbs and the hepatitis B surface antigen (HBsAg) can affect DC function and enhance T-cell responses. HBsAg/anti-HBV ICs significantly increased the uptake of the immunocomplexed HBsAg antigen, and augmented the in vitro proliferation of virus-specific T cells and their production of interferon (IFN)-γ.Citation55 Moreover, DCs from HBV-infected patients incubated with HBsAg/anti-HBV ICs showed higher expression of major histocompatibility complex (MHC)-II molecules and higher production of interleukin (IL)-12. IC-loaded DCs also enhanced production of IL-2 and IFN-γ by co-cultured T cells.Citation56 Interestingly, the therapeutic efficacy of HBsAg/anti-HBV ICs has been tested in clinical trials (see below) in HBV-infected patients with encouraging results.Citation60, Citation61, Citation62 More recently, in experiments aimed at visualizing immunopotentialization by HBsAg/anti-HBV ICs (see below), live-cell imaging revealed that ICs were internalized via the FcγRs of APCs and were subsequently transported through early and late endosomes into lysosomes, where they co-localized with MHC-I and MHC-II molecules.Citation63 Consistent with the latter observation, the administration of DCs loaded with HBsAg/anti-HBV ICs to mice increased the number of IFN-γ- and tumor necrosis factor-α-producing CD8+ and CD4+ T cells. Similarly, in an SIV infection setting, the incubation of APCs with ICs made with a recombinant full-length Gag p55 protein and an anti-p55 IgG increased SIV capsid cross-presentation. Capsid cross-presentation was dependent on FcγR-mediated uptake of the immunocomplexed SIV capsid protein, and required its proteasomal and endosomal degradation to generate stronger Gag-specific CD8+ T-cell responses.Citation57 From a mechanistic standpoint, these studies indicate that antiviral antibodies might enhance the priming and expansion of virus-specific CD4+ and CD8+ T cells by both promoting the secretion of key cytokines and facilitating the uptake and cross-presentation of viral Ags by FcγR-expressing DCs.
Immune-complexed whole virions have also been shown to affect the functional activation of DCs. The stimulation of DCs with ICs composed of SIV virions and highly neutralizing SIV-hyperimmune sera (SVIG) led to the increased virus-specific CD4+ T-cell responses in an Fc-dependent manner.Citation58 In contrast, DCs stimulated with ICs made of HIV-1 and a polyclonal IgG pool from HIV-infected subjects showed only weak HIV-specific CTL-stimulating activity. This suggested that opsonization of HIV-1 by IgGs might be associated with decreased CTL-stimulatory DC activity.Citation59 However, not all IgG isotypes display equivalent effector functions. Therefore, the undefined nature of the antibodies (both in terms of predominant isotypes and neutralization potential) used to form the HIV-ICs in these experiments might explain these observations. Whether HIV-ICs made with highly neutralizing anti-HIV mAbs of a specific IgG isotype might induce stronger CD8+ T-cell responses is an important issue deserving further investigation (). Moreover, the nature of the viral determinant present in ICs might also be crucial in the stimulation of antiviral responses. Interestingly, as mentioned above, in the mouse FrCasE infection model, ICs made up of a neutralizing mAb and infected cells, but not those made with virions, efficiently induce strong Gag-specific CD8+ T-cell responses with high cytotoxic activity.Citation43 This observation shows that the viral and cellular ICs can trigger different immunologic outcomes. In the case of FrCasE, this is explained by the fact the FrCasE-GagL CTL immunodominant epitope is, at best, poorly incorporated into virions. Taken together, these data indicate that the uptake of cellular ICs might allow the presentation of a broader viral antigenic repertoire, leading to stronger effects on immunity ().
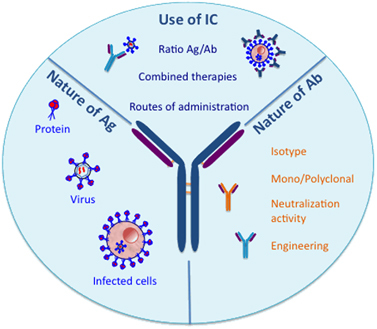
Figure 2 Parameters to consider for achieving optimal IC-mediated modulation of antiviral immune responses. The optimization of vaccine-like-effect-inducing protocols will require the consideration of several parameters such as the nature of the antigen (that is, purified viral proteins, whole virions and infected cells) and the antibody (that is, monoclonal vs polyclonal, nature of the isotype, engineered Fc domain with improved effector functions and so on) used to form the immunogenic ICs, as differences in these parameters might impact immunological outcomes. In addition, whether the optimized ICs are used alone or in combination with immunostimulatory molecules might also be of paramount importance. Several other parameters, including the IC dosage, the route of administration, the choice of adjuvant and the immunological status of patients, will also have to be considered.
MODULATION OF ANTIVIRAL IMMUNE RESPONSES BY ICS FORMED WITH ENDOGENOUS ANTIBODIES DURING THE COURSE OF VIRAL INFECTIONS
ICs formed with endogenous antibodies generated in virally infected mice have been shown to influence antiviral cellular immune responses in several models (). Notably, the highly neutralizing humoral response generated against the FrCasE retrovirus in mAb-treated-infected mice was demonstrated to limit the viral propagation and to enhance memory cellular responses long after the disappearance of the therapeutic mAb (which occurs within two weeks post administration, reflecting the natural IgG lifespan in vivo). IC-mediated activation of DCs upon binding to FcγR was key for this effect.Citation43 Similarly, in an influenza virus infection model, ICs formed with endogenous antiviral antibodies promoted more sustained antigen presentation by DCs, resulting in stronger CD8+ T-cell proliferation.Citation64 Interestingly, such prolonged antigen presentation by DCs was dependent on virus-specific, isotype-switched antibodies that facilitated the capture and cross-presentation of viral antigens by FcγR-expressing DCs. In addition, serum antibodies can affect the virus-specific CD4+/CD8+ T-cell balance in an Fc-dependent manner during RSV infection.Citation65 An enhanced ratio of RSV-neutralizing to -non-neutralizing antibodies profoundly enhanced the CD4+ T-cell response. In a murine lymphocytic choriomeningitis virus (LCMV) infection model, endogenous virus-specific antibodies could stimulate innate immunity and thereby positively affect both the induction and the maintenance of the virus-specific CD8+ T-cell response. Notably, anti-LCMV antibodies limited viral replication in peripheral organs, but allowed replication of the virus in the marginal zone of the spleen, promoting CD8+ T-cell priming.Citation66 Interestingly, anti-LCMV antibodies were also reported to be essential for long-term maintenance of the memory CTL response in infected mice.Citation67, Citation68
Table 2 Modulation of immune responses by ICs involving antiviral host humoral responses
These observations, together with the in vitro studies described above, demonstrate that virus-specific antibodies can promote the acquisition, processing and presentation of antigens that are subsequently instrumental for priming T-cell responses and programming functional CD8+ memory in an Fc-dependent manner. They strengthen the concept that antiviral antibodies can regulate the quality and function of antiviral T-cell responses through the formation of ICs. Moreover, they also provide a rationale for developing novel IC-based therapeutic vaccination strategies.
ENHANCEMENT OF ANTIVIRAL IMMUNE RESPONSES IN IC-BASED VACCINATION STRATEGIES
In 1961, Terres and WolinsCitation69 demonstrated the ability of ICs to induce higher antibody titers than antigens alone. Since then, the immunogenic potential of ICs, alone or in combination with different types of adjuvants, has been tested in several viral infection systems, including animal models of human infections, for example, those involving HBV, HIV, RSV or flaviviruses. The immunostimulatory are principally attributed to the ability of Fc antibody fragments to recruit the host immune system. However, evidence also implicates Fab fragments in modulation of the antiviral immune response, although the outcomes are less documented and were proposed to occur via alterations in antigen conformation and/or in the exposure of specific epitopes.
We describe the enhancement in antiviral immune responses observed in IC-based vaccination experiments below ().
Table 3 Vaccine strategies based on immunogenic ICs
HBV
ICs have been tested as vaccines to augment protective immune responses in different animal models of HBV infection. In ducks, ICs made with duck HBsAg and rabbit anti-duck HBsAg (DHBsAg/anti-DHB) were used as immunogens in the form of solid matrix–antibody–antigen complexes (SMAA). Such SMAAs contained killed Staphylococcus aureus as a solid matrix and mAb-opsonized viruses.Citation79 They were initially shown to induce both humoral and CTL responses against the paramyxovirus simian virus 5 in immunized mice.Citation80 Immunization of HBV-infected ducks with SMAA-based DHBsAg/anti-DHB ICs led to the viral clearance in 60% of infected ducks. Notably, the administration of DHBsAg/anti-DHB ICs lacking Staphylococcus aureus showed decreased immunization efficiency, suggesting that the bacteria-based solid matrix functions as an adjuvant.Citation70
ICs have also been tested as a therapeutic vaccine against HBV infection in miceCitation63, Citation71, Citation72 and woodchucks.Citation73 BALB/c mice immunized with HBsAg/anti-HBV ICs produced the increased levels of virus-specific antibodies.Citation71 Moreover, administering HBsAg/anti-HBV ICs to BALB/c mice via intranasal inhalation induced both mucosal and systemic Th1-polarized immune responses, when administered with adjuvants such as cholera toxin or oligodeoxynucleotides containing immunostimulatory CpG motifs (CpG). This was not observed using HBsAg alone.Citation74 In addition, the administration of HBsAg/anti-HBV ICs to HBsAg-positive transgenic mice decreased the serum HBsAg levels and induced stronger CTL responses than HBsAg alone.Citation72 Notably, the co-administration of ICs and a plasmid coding for HBsAg increased the antiviral immune response induced by ICs, indicating the adjuvant effect of DNA in this setting. A similar effect was also reported in a woodchuck model of HBV infection: immunization of woodchuck hepatitis virus (WHV)-infected animals with WHV surface antigen/anti-WHV antibody ICs combined with a DNA vaccine resulted in a higher reduction of both viral load and antigenemia relative to ICs alone.Citation73 Interestingly, the WHV-infected animals were pretreated with lamivudine (a potent HBV antiviral drug able to enhance T-cell responses in chronically HBV-infected patients) before IC/DNA immunization, suggesting that combination strategies should be considered in treating chronic HBV infections ().
The enhancement of antiviral immune responses by ICs in vitro and in preclinical models of HBV infection paved the way for the development of IC-based therapeutic vaccination strategies against chronic viral hepatitis B infection. A therapeutic vaccine composed of yeast-derived recombinant HBsAg/anti-HBV immunogenic complexes (YICs) has been tested in a series of clinical trials. This vaccine approach was initially shown to be safe and to induce higher titers of HBsAg antibodies, as well as to increase serum IFN-γ and IL-2 levels in a phase I trial.Citation60 Importantly, a subpopulation of chronic viral hepatitis B patients showed a decrease in serum HBV viral load and HBsAg levels together with an increase in anti-HBsAg antibody titers in subsequent phase II trials.Citation61, Citation62 From a mechanistic standpoint, recent data showing that the administration of YIC-loaded DCs to mice increased both CD8+ and CD4+ T-cell responsesCitation63 suggest that the improved immune responses induced by YICs might account for the antiviral effect observed in a fraction of patients. In an attempt to enhance the immunogenic potential of YIC-based vaccines, a phase III trial tested the therapeutic effect of a higher number of IC doses. Unfortunately, overstimulation with YIC decreased the vaccine efficiency due to host immune fatigue.Citation81 This suggests that vaccination protocols must be optimized and must take into account both the nature and the dose of ICs, as well as other parameters such as the route of administration, the type of adjuvant and the immunological status of patients to achieve efficient protective immunization ().
HIV
In 1988, a study immunized healthy volunteers with HIV peptides.Citation82 The authors found that compared with free antigen, recall immunization with ICs induced stronger T-cell responses through uncharacterized mechanisms. Other reports also describe alteration of the anti-HIV response by ICs.Citation75, Citation76, Citation77, Citation83 In particular, the immunization of immunocompetent mice with ICs containing a recombinant HIV-1 gp120 Env glycoprotein and a mAb (654-D mAb) directed to the CD4-binding site induced a higher virus-neutralizing antibody response relative to free antigen. As described above, humoral responses were further increased upon the co-administration of ICs and monophosphoryl lipid-A/dimethyldioctadecylammonium adjuvants. Notably, the interaction of the anti-CD4-binding site mAb with HIV-1 gp120 induced conformational changes in the latter, leading to the enhanced antigenicity and immunogenicity of neutralizing epitopes localized in the HIV-1 V3 loop.Citation75 These observations highlight the ability of anti-HIV-1 antibodies to induce antigenic alterations in specific HIV-1 gp120 epitopes upon IC formation. Interestingly, further improvement in the immunogenicity of the V3 loop was obtained in ICs generated with gp120 mutants lacking site-specific N-linked glycans.Citation76, Citation77, Citation83 Taken together, these observations suggest that the ability of ICs to stimulate the induction of neutralizing antibodies is dictated by the nature of the antigen, as well as the specificity and affinity of the mAbs utilized. These results also indicate the potential contribution of Fab-mediated activities in the enhancement of antiviral humoral responses by ICs.
Flavivirus
Tsouchnikas et al.Citation84 investigated the influence of immunization with ICs on the specificity of antibody responses using the E protein of the tick-borne encephalitis virus as an immunogen. Mice were immunized with a dimeric soluble form of E (sE) alone or in complex with mAbs specific for each of the three domains of E. The antibody response induced by these ICs was compared with that observed after immunization with sE alone. Unexpectedly, immunization with ICs did not change the extent of the overall antibody response in immunized mice. However, substantially different antibody responses were observed between the different ICs. These differences most likely reflected an epitope-shielding phenomenon and antibody-mediated structural changes that led to the dissociation of the sE dimer. Thus, such phenomena can profoundly influence the fine specificity of antibody responses to the same immunogen and must be considered in IC-based vaccination strategies.
RSV
As mentioned above, serum anti-RSV antibodies can affect virus-specific T-cell responses.Citation65 On the basis of this, Kruijsen et al.Citation78 tested whether ICs made with the commercial RSV-neutralizing mAb palivizumab could influence adaptive immune response priming after intranasal administration. Substantial anti-RSV T-cell priming and B-cell responses were observed in mice receiving RSV-ICs, resulting in predominant Th1-type CD4+ T-cell response and IgG2c antibody responses. Importantly, the ICs also primed anti-RSV CD8+ T cells. These data have important implications for the prophylaxis and treatment of pediatric RSV infections. Nevertheless, interactions between ICs and neonatal versus adult innate and adaptive immune systems still need to be investigated because mouse studies have revealed potential antibody-induced neonatal autoimmunity in certain settings.Citation85, Citation86
ICS AND IMMUNE DYSFUNCTION
In the course of viral infections, the formation of ICs composed of viral determinants and the resulting host humoral responses can potentially produce deleterious effects. Persistent ICs are formed in a variety of chronic viral infections and may lead to unregulated and protracted FcγR signaling. This may lead to immune dysfunction instead of stimulating antiviral immune responses. In this regard, the high levels of ICs formed during LCMV infection interfere with FcγR-mediated activities.Citation87, Citation88 These endogenously formed ICs were shown to outcompete the effector functions of exogenously administered therapeutic mAbs, in particular binding to FcγRs expressed by immune cells. Persistent endogenous ICs are also linked to dysfunctional B-cell responses in HIV infection, including the suppression of antiviral IgA responses and impaired production of neutralizing antibodies (reviewed in Moir et al.Citation89). The composition of ICs might also negatively affect the efficiency of the antiviral immune response. For instance, the composition of ICs has been shown to be dynamic throughout the course of HIV infections due to changes in both antibody specificities and virion levels. Notably, circulating ICs are initially comprised of antibodies that opsonize both infectious and non-infectious virions. This results in a decrease in the availability of antibodies able to blunt viral propagation. This phenomenon probably contributes to the reduced efficiency of the antibodies generated during acute infection.Citation90 Changes in circulating ICs have also been reported in HCV infections. The level of circulating ICs is low in acutely infected patients, whereas chronically infected individuals show a high proportion of immunocomplexed HCV, raising the possibility that ICs may have a role in the pathogenesis of HCV, namely liver damage.Citation91 Moreover, the formation of ICs with non-neutralizing antibodies may also lead to the antibody-dependent enhancement of viral infection of FcγR-expressing cells. This happens in several viral infections, including those by the dengue virus.Citation92, Citation93, Citation94, Citation95 Along this line, the binding of ICs to FcγRs on monocytes/macrophages can paradoxically suppress innate immunity, induce IL-10 production and bias responses from Th1 toward Th2. This in turn leads to the increased infectious outputs by infected cells via intrinsic antibody-dependent enhancement.Citation94, Citation96
Finally, ICs have also been reported to have a role in increasing viral loads in the context of gene transfer-based vaccination strategies. In the STEP HIV-1 vaccine trial, which evaluated a replication-defective adenovirus type 5 vector vaccine, the ICs formed with pre-existing anti-Adv5 antibodies improved the environment for HIV-1 replication in T cells. This may have been due to the IC-driven activation of a DC–T-cell axis that induces the activation of CD4+ T cells and leads to a permissive environment for HIV-1 infection. This environment probably explains the increased propagation of HIV-1 infection among adenovirus type 5-seropositive vaccine recipients.Citation97
IMPROVEMENT OF POTENTIAL VACCINE-LIKE EFFECTS OF MAB-BASED IMMUNOTHERAPIES
Several approaches can be considered to enhance the immunomodulatory potential of antiviral mAbs, both alone and in the form of ICs, in particular through combining neutralizing mAbs and IC-based vaccination strategies with other therapies. A first possibility would consist of inhibiting immunosuppressive mechanisms in infected individuals by either depleting the Treg response, as suggested by Nasser et al.Citation45 or targeting immune checkpoints, the latter strategy having already led to improved immune responses against both viral infections and cancer.Citation98, Citation99, Citation100, Citation101 In addition, the combination of antiviral mAbs with different immunostimulatory agents can also be envisaged. Because the primary structure and glycosylation pattern of the Fc fragment are both essential for antibody effector functions due to their impact on the engagement of type I and type II FcR family members,Citation26, Citation27 Fc engineering might also represent another approach, not only to improve direct antiviral effects, but also to induce stronger vaccine-like effects. In this regard, identification of the main FcRs and FcR-mediated mechanisms involved in enhancing the antiviral immune response will be of utmost importance. Taking into account that the various IgG isotypes display different effector functions and interact differently with FcγRs, the careful selection of antiviral mAb subclasses is also crucial for enhancing antiviral immune responses. Finally, as FcγR polymorphisms have already been associated with differences in viral disease progression and the therapeutic efficiency of anticancer mAbs, it will be important to evaluate the extent to which such polymorphisms can affect the vaccine-like effects induced by mAb-based antiviral immunotherapies.
CONCLUSION
The therapeutic potential of antiviral mAbs is now widely accepted, and their use as antiviral drugs is increasingly under consideration. The diverse biological activities of these mAbs lead to the direct control of viral propagation and the modulation of antiviral immunity. This provides a novel rationale for their use in diverse prophylactic and therapeutic approaches. The improvement in both humoral and cellular responses achieved through the administration of mAbs, either free or in the form of immunogenic ICs, offers new therapeutic options. The challenge now is to improve our understanding of how ICs convert mAb-based immunotherapies from ‘passive’ to ‘active’ and to exploit the underlying mechanisms. This conversion will be crucial in reaching the goal of using antiviral mAbs to induce long-lasting protective immunity against life-threatening viral infections.
This work was supported by grants from the ‘Ligue Nationale contre le Cancer’, the ‘Fondation ARC’, ‘Sidaction’ and the ‘Fondation pour la Recherche Médicale’. Jennifer Lambour, Mar Naranjo-Gomez, Marc Piechaczyk and Mireia Pelegrin are part of the ‘MabImprove Labex’, a public grant overseen by the French National Research Agency (ANR) as part of the ‘Investissements d’Avenir’ program (reference: ANR-10-LABX -53-01) that also supported this work. We are grateful to Drs Eric Kremer (Institute of Molecular Genetics of Montpellier, France) and Robert Hipskind (Institute of Molecular Genetics of Montpellier, France) for critical reading of the manuscript.
Related Research Data
- IraniV,GuyAJ,AndrewD,BeesonJG,RamslandPA,RichardsJS.Molecular properties of human IgG subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases.Mol Immunol2015; 67:171–182.
- VacchelliE,EggermontA,GalonJet al.Trial watch: monoclonal antibodies in cancer therapy.Oncoimmunology2013; 2:e22789.
- BornholdtZA,TurnerHL,MurinCDet al.Isolation of potent neutralizing antibodies from a survivor of the 2014 Ebola virus outbreak.Science2016; 351:1078–1083.
- CortiD,MisasiJ,MulanguSet al.Protective monotherapy against lethal Ebola virus infection by a potently neutralizing antibody.Science2016; 351:1339–1342.
- CortiD,ZhaoJ,PedottiMet al.Prophylactic and postexposure efficacy of a potent human monoclonal antibody against MERS coronavirus.Proc Natl Acad Sci USA2015; 112:10473–10478.
- JohnsonRF,BagciU,KeithLet al.3B11-N, a monoclonal antibody against MERS-CoV, reduces lung pathology in rhesus monkeys following intratracheal inoculation of MERS-CoV Jordan-n3/2012.Virology2016; 490:49–58.
- QiuX,WongG,AudetJet al.Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp.Nature2014; 514:47–53.
- BarouchDH,WhitneyJB,MoldtBet al.Therapeutic efficacy of potent neutralizing HIV-1-specific monoclonal antibodies in SHIV-infected rhesus monkeys.Nature2013; 503:224–228.
- BerryJD,GaudetRG.Antibodies in infectious diseases: polyclonals, monoclonals and niche biotechnology.N Biotechnol2011; 28:489–501.
- BothL,BanyardAC,van DolleweerdC,WrightE,MaJK,FooksAR.Monoclonal antibodies for prophylactic and therapeutic use against viral infections.Vaccine2013; 31:1553–1559.
- CortiD,LanzavecchiaA.Broadly neutralizing antiviral antibodies.Annu Rev Immunol2013; 31:705–742.
- de JongYP,DornerM,MommersteegMCet al.Broadly neutralizing antibodies abrogate established hepatitis C virus infection.Sci Transl Med2014; 6:254ra129.
- DejnirattisaiW,WongwiwatW,SupasaSet al.A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus.Nat Immunol2015; 16:170–177.
- FlegoM,AscioneA,CianfrigliaM,VellaS.Clinical development of monoclonal antibody-based drugs in HIV and HCV diseases.BMC Med2013; 11:4.
- FlyakAI,IlinykhPA,MurinCDet al.Mechanism of human antibody-mediated neutralization of Marburg virus.Cell2015; 160:893–903.
- FlyakAI,ShenX,MurinCDet al.Cross-reactive and potent neutralizing antibody responses in human survivors of natural Ebolavirus infection.Cell2015; 164:392–405.
- GautamR,NishimuraY,PeguAet al.A single injection of anti-HIV-1 antibodies protects against repeated SHIV challenges.Nature2016; 533:105–109.
- HessellAJ,JaworskiJP,EpsonEet al.Early short-term treatment with neutralizing human monoclonal antibodies halts SHIV infection in infant macaques.Nat Med2016; 22:362–368.
- KrawczykA,ArndtMA,Grosse-HovestLet al.Overcoming drug-resistant herpes simplex virus (HSV) infection by a humanized antibody.Proc Natl Acad Sci USA2013; 110:6760–6765.
- MarascoWA,SuiJ.The growth and potential of human antiviral monoclonal antibody therapeutics.Nat Biotechnol2007; 25:1421–1434.
- TanY,NgQ,JiaQ,KwangJ,HeF.A novel humanized antibody neutralizes H5N1 influenza virus via two different mechanisms.J Virol2015; 89:3712–3722.
- WilliamsKL,Sukupolvi-PettyS,BeltramelloMet al.Therapeutic efficacy of antibodies lacking Fcgamma receptor binding against lethal dengue virus infection is due to neutralizing potency and blocking of enhancing antibodies [corrected].PLoS Pathog2013; 9:e1003157.
- CaskeyM,KleinF,LorenziJCet al.Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117.Nature2015; 522:487–491.
- LynchRM,BoritzE,CoatesEEet al.Virologic effects of broadly neutralizing antibody VRC01 administration during chronic HIV-1 infection.Sci Transl Med2015; 7:319ra206.
- BruhnsP,JonssonF.Mouse and human FcR effector functions.Immunol Rev2015; 268:25–51.
- PinceticA,BournazosS,DiLilloDJet al.Type I and type II Fc receptors regulate innate and adaptive immunity.Nat Immunol2014; 15:707–716.
- EulerZ,AlterG.Exploring the potential of monoclonal antibody therapeutics for HIV-1 eradication.AIDS Res Hum Retroviruses2014; 31:13–24.
- HessellAJ,HaigwoodNL.Neutralizing antibodies and control of HIV: moves and countermoves.Curr HIV/AIDS Rep2012; 9:64–72.
- SuB,MoogC.Which antibody functions are important for an HIV vaccine?Front Immunol2014; 5:289.
- NimmerjahnF,GordanS,LuxA.FcgammaR dependent mechanisms of cytotoxic, agonistic, and neutralizing antibody activities.Trends Immunol2015; 36:325–336.
- MalbecM,PorrotF,RuaRet al.Broadly neutralizing antibodies that inhibit HIV-1 cell to cell transmission.J Exp Med2013; 210:2813–2821.
- BruhnsP.Properties of mouse and human IgG receptors and their contribution to disease models.Blood2012; 119:5640–5649.
- GetahunA,HeymanB.How antibodies act as natural adjuvants.Immunol Lett2006; 104:38–45.
- HeymanB.Antibodies as natural adjuvants.Curr Top Microbiol Immunol2014; 382:201–219.
- HjelmF,CarlssonF,GetahunA,HeymanB.Antibody-mediated regulation of the immune response.Scand J Immunol2006; 64:177–184.
- NimmerjahnF,RavetchJV.Fcgamma receptors as regulators of immune responses.Nat Rev Immunol2008; 8:34–47.
- SchwabI,NimmerjahnF.Intravenous immunoglobulin therapy: how does IgG modulate the immune system?Nat Rev Immunol2013; 13:176–189.
- BradyLJ.Antibody-mediated immunomodulation: a strategy to improve host responses against microbial antigens.Infect Immun2005; 73:671–678.
- PelegrinM,Naranjo-GomezM,PiechaczykM.Antiviral monoclonal antibodies: can they be more than simple neutralizing agents?Trends Microbiol2015; 23:653–665.
- GrosL,DrejaH,FiserAL,PlaysM,PelegrinM,PiechaczykM.Induction of long-term protective antiviral endogenous immune response by short neutralizing monoclonal antibody treatment.J Virol2005; 79:6272–6280.
- GrosL,PelegrinM,MichaudHAet al.Endogenous cytotoxic T-cell response contributes to the long-term antiretroviral protection induced by a short period of antibody-based immunotherapy of neonatally infected mice.J Virol2008; 82:1339–1349.
- GrosL,PelegrinM,PlaysM,PiechaczykM.Efficient mother-to-child transfer of antiretroviral immunity in the context of preclinical monoclonal antibody-based immunotherapy.J Virol2006; 80:10191–10200.
- MichaudHA,GomardT,GrosLet al.A crucial role for infected-cell/antibody immune complexes in the enhancement of endogenous antiviral immunity by short passive immunotherapy.PLoS Pathog2010; 6:e1000948.
- NasserR,PelegrinM,MichaudHAet al.Long-lasting protective antiviral immunity induced by passive immunotherapies requires both neutralizing and effector functions of the administered monoclonal antibody.J Virol2010; 84:10169–10181.
- NasserR,PelegrinM,PlaysM,GrosL,PiechaczykM.Control of regulatory T cells is necessary for vaccine-like effects of antiviral immunotherapy by monoclonal antibodies.Blood2013; 121:1102–1111.
- Boyoglu-BarnumS,ChirkovaT,ToddSOet al.Prophylaxis with a respiratory syncytial virus (RSV) anti-G protein monoclonal antibody shifts the adaptive immune response to RSV rA2-line19F infection from Th2 to Th1 in BALB/c mice.J Virol2014; 88:10569–10583.
- BossartKN,GeisbertTW,FeldmannHet al.A neutralizing human monoclonal antibody protects african green monkeys from hendra virus challenge.Sci Transl Med2011; 3:105ra103.
- GeisbertTW,MireCE,GeisbertJBet al.Therapeutic treatment of Nipah virus infection in nonhuman primates with a neutralizing human monoclonal antibody.Sci Transl Med2014; 6:242ra282.
- JaworskiJP,KobieJ,BrowerZet al.Neutralizing polyclonal IgG present during acute infection prevents rapid disease onset in simian-human immunodeficiency virus SHIVSF162P3-infected infant rhesus macaques.J Virol2013; 87:10447–10459.
- NgCT,JaworskiJP,JayaramanPet al.Passive neutralizing antibody controls SHIV viremia and enhances B cell responses in infant macaques.Nat Med2010; 16:1117–1119.
- WatkinsJD,SiddappaNB,LakhasheSKet al.An anti-HIV-1 V3 loop antibody fully protects cross-clade and elicits T-cell immunity in macaques mucosally challenged with an R5 clade C SHIV.PLoS One2011; 6:e18207.
- SchoofsT,KleinF,BraunschweigMet al.HIV-1 therapy with monoclonal antibody 3BNC117 elicits hoost immune responses against HIV-1.Science2016; 352:997–1001.
- BournazosS,KleinF,PietzschJ,SeamanMS,NussenzweigMC,RavetchJV.Broadly neutralizing anti-HIV-1 antibodies require Fc effector functions for in vivo activity.Cell2014; 158:1243–1253.
- Halper-StrombergA,LuCL,KleinFet al.Broadly neutralizing antibodies and viral inducers decrease rebound from HIV-1 latent reservoirs in humanized mice.Cell2014; 158:989–999.
- CelisE,ChangTW.HBsAg-serum protein complexes stimulate immune T lymphocytes more efficiently than do pure HBsAg.Hepatology1984; 4:1116–1123.
- ZhengBJ,ZhouJ,QuDet al.Selective functional deficit in dendritic cell—T cell interaction is a crucial mechanism in chronic hepatitis B virus infection.J Viral Hepat2004; 11:217–224.
- VillingerF,MayneAE,BostikPet al.Evidence for antibody-mediated enhancement of simian immunodeficiency virus (SIV) Gag antigen processing and cross presentation in SIV-infected rhesus macaques.J Virol2003; 77:10–24.
- YamamotoT,IwamotoN,YamamotoHet al.Polyfunctional CD4+ T-cell induction in neutralizing antibody-triggered control of simian immunodeficiency virus infection.J Virol2009; 83:5514–5524.
- PoschW,CardinaudS,HamimiCet al.Antibodies attenuate the capacity of dendritic cells to stimulate HIV-specific cytotoxic T lymphocytes.J Allergy Clin Immunol2012; 130:1368–1374 e1362.
- XuDZ,HuangKL,ZhaoKet al.Vaccination with recombinant HBsAg-HBIG complex in healthy adults.Vaccine2005; 23:2658–2664.
- YaoX,ZhengB,ZhouJet al.Therapeutic effect of hepatitis B surface antigen-antibody complex is associated with cytolytic and non-cytolytic immune responses in hepatitis B patients.Vaccine2007; 25:1771–1779.
- XuDZ,ZhaoK,GuoLMet al.A randomized controlled phase IIb trial of antigen-antibody immunogenic complex therapeutic vaccine in chronic hepatitis B patients.PLoS One2008; 3:e2565.
- LiuH,GengS,WangBet al.Immuno-potentiating pathway of HBsAg-HBIG immunogenic complex visualized.Hum Vaccin Immunother2015; 12:77–84.
- LeonB,Ballesteros-TatoA,RandallTD,LundFE.Prolonged antigen presentation by immune complex-binding dendritic cells programs the proliferative capacity of memory CD8 T cells.J Exp Med2014; 211:1637–1655.
- KruijsenD,BakkersMJ,van UdenNOet al.Serum antibodies critically affect virus-specific CD4+/CD8+ T cell balance during respiratory syncytial virus infections.J Immunol2010; 185:6489–6498.
- DuhanV,KhairnarV,FriedrichSKet al.Virus-specific antibodies allow viral replication in the marginal zone, thereby promoting CD8(+) T-cell priming and viral control.Sci Rep2016; 6:19191.
- BachmannMF,HunzikerL,ZinkernagelRM,StorniT,KopfM.Maintenance of memory CTL responses by T helper cells and CD40-CD40 ligand: antibodies provide the key.Eur J Immunol2004; 34:317–326.
- BergthalerA,FlatzL,VerschoorAet al.Impaired antibody response causes persistence of prototypic T cell-contained virus.PLoS Biol2009; 7:e80.
- TerresG,WolinsW.Enhanced immunological sensitization of mice by the simultaneous injection of antigen and specific antiserum. I. Effect of varying the amount of antigen used relative to the antiserum.J Immunol1961; 86:361–368.
- WenYM,QuD,ZhouSH.Antigen-antibody complex as therapeutic vaccine for viral hepatitis B.Int Rev Immunol1999; 18:251–258.
- BasalpA,CirakogluB,BermekE.Enhancement of the immune response to hepatitis B virus vaccine by antigen specific IgM.Immunol Lett2000; 73:1–6.
- ZhengBJ,NgMH,HeLFet al.Therapeutic efficacy of hepatitis B surface antigen-antibodies-recombinant DNA composite in HBsAg transgenic mice.Vaccine2001; 19:4219–4225.
- LuM,YaoX,XuYet al.Combination of an antiviral drug and immunomodulation against hepadnaviral infection in the woodchuck model.J Virol2008; 82:2598–2603.
- McCluskieMJ,WenYM,DiQ,DavisHL.Immunization against hepatitis B virus by mucosal administration of antigen-antibody complexes.Viral Immunol1998; 11:245–252.
- HioeCE,ViscianoML,KumarRet al.The use of immune complex vaccines to enhance antibody responses against neutralizing epitopes on HIV-1 envelope gp120.Vaccine2009; 28:352–360.
- KumarR,TuenM,LiH,TseDB,HioeCE.Improving immunogenicity of HIV-1 envelope gp120 by glycan removal and immune complex formation.Vaccine2011; 29:9064–9074.
- KumarR,TuenM,LiuJet al.Elicitation of broadly reactive antibodies against glycan-modulated neutralizing V3 epitopes of HIV-1 by immune complex vaccines.Vaccine2013; 31:5413–5421.
- KruijsenD,EinarsdottirHK,SchijfMAet al.Intranasal administration of antibody-bound respiratory syncytial virus particles efficiently primes virus-specific immune responses in mice.J Virol2013; 87:7550–7557.
- RandallRE,YoungDF,SouthernJA.Immunization with solid matrix-antibody-antigen complexes containing surface or internal virus structural proteins protects mice from infection with the paramyxovirus, simian virus 5.J Gen Virol1988; 69:2517–2526.
- RandallRE,YoungDF.Solid matrix-antibody-antigen complexes induce antigen-specific CD8+ cells that clear a persistent paramyxovirus infection.J Virol1991; 65:719–726.
- XuDZ,WangXY,ShenXLet al.Results of a phase III clinical trial with an HBsAg-HBIG immunogenic complex therapeutic vaccine for chronic hepatitis B patients: experiences and findings.J Hepatol2013; 59:450–456.
- BerzofskyJA,BensussanA,CeaseKBet al.Antigenic peptides recognized by T lymphocytes from AIDS viral envelope-immune humans.Nature1988; 334:706–708.
- KumarR,ViscianoML,LiH,HioeC.Targeting a neutralizing epitope of HIV envelope Gp120 by immune complex vaccine.J AIDS Clin Res2012; S8:pii: 5512.
- TsouchnikasG,ZlatkovicJ,JarmerJet al.Immunization with immune complexes modulates the fine specificity of antibody responses to a flavivirus antigen.J Virol2015; 89:7970–7978.
- RivalC,SamyE,SetiadyY,TungK.Cutting edge: Ly49C/I(-) neonatal NK cells predispose newborns to autoimmune ovarian disease induced by maternal autoantibody.J Immunol2013; 191:2865–2869.
- RivalC,SetiadyY,SamyET,HarakalJ,TungKS.The unique neonatal NK cells: a critical component required for neonatal autoimmune disease induction by maternal autoantibody.Front Immunol2014; 5:242.
- WielandA,ShashidharamurthyR,KamphorstAOet al.Antibody effector functions mediated by Fcgamma-receptors are compromised during persistent viral infection.Immunity2015; 42:367–378.
- YamadaDH,ElsaesserH,LuxAet al.Suppression of Fcgamma-receptor-mediated antibody effector function during persistent viral infection.Immunity2015; 42:379–390.
- MoirS,FauciAS.B cells in HIV infection and disease.Nat Rev Immunol2009; 9:235–245.
- LiuP,OvermanRG,YatesNLet al.Dynamic antibody specificities and virion concentrations in circulating immune complexes in acute to chronic HIV-1 infection.J Virol2011; 85:11196–11207.
- RivaE,MaggiF,AbbruzzeseFet al.Immune complexed (IC) hepatitis C virus (HCV) in chronically and acutely HCV-infected patients.Med Microbiol Immunol2009; 198:13–18.
- AcostaEG,BartenschlagerR.Paradoxical role of antibodies in dengue virus infections: considerations for prophylactic vaccine development.Expert Rev Vaccines2016; 15:467–482.
- HalsteadSB.Dengue antibody-dependent enhancement: knowns and unknowns.Microbiol Spectr2014 doi:10.1128/microbiolspec.AID-0022-2014.
- HalsteadSB,MahalingamS,MarovichMA,UbolS,MosserDM.Intrinsic antibody-dependent enhancement of microbial infection in macrophages: disease regulation by immune complexes.Lancet Infect Dis2010; 10:712–722.
- UbolS,HalsteadSB.How innate immune mechanisms contribute to antibody-enhanced viral infections.Clin Vaccine Immunol2010; 17:1829–1835.
- TsaiTT,ChuangYJ,LinYSet al.Antibody-dependent enhancement infection facilitates dengue virus-regulated signaling of IL-10 production in monocytes.PLoS Negl Trop Dis2014; 8:e3320.
- PerreauM,PantaleoG,KremerEJ.Activation of a dendritic cell-T cell axis by Ad5 immune complexes creates an improved environment for replication of HIV in T cells.J Exp Med2008; 205:2717–2725.
- BengschB,MartinB,ThimmeR.Restoration of HBV-specific CD8+ T cell function by PD-1 blockade in inactive carrier patients is linked to T cell differentiation.J Hepatol2014; 61:1212–1219.
- GardinerD,LalezariJ,LawitzEet al.A randomized, double-blind, placebo-controlled assessment of BMS-936558, a fully human monoclonal antibody to programmed death-1 (PD-1), in patients with chronic hepatitis C virus infection.PLoS One2013; 8:e63818.
- SharmaP,AllisonJP.Immune checkpoint targeting in cancer therapy: toward combination strategies with curative potential.Cell2015; 161:205–214.
- ShermanAC,TrehanpatiN,DaucherMet al.Augmentation of hepatitis B virus-specific cellular immunity with programmed death receptor-1/programmed death receptor-L1 blockade in hepatitis B virus and HIV/hepatitis B virus coinfected patients treated with adefovir.AIDS Res Hum Retroviruses2013; 29:665–672.