
Abstract
Middle East respiratory syndrome coronavirus (MERS-CoV) has caused fatal infections, some through hospital-acquired transmission, in affected regions since its emergence in 2012. Although the virus is not pandemic among humans, it poses a great threat to public health due to its zoonotic origin. Thus, both preventative and therapeutic countermeasures are urgently needed. In this study, we discovered a panel of neutralizing monoclonal antibodies (mAbs) against MERS-CoV, which mapped to a wide range of regions on the spike (S) protein of the virus. In addition to mAbs with neutralizing epitopes located on the receptor-binding domain, one mAb, 5F9, which binds to the N-terminal domain (NTD) of the MERS-CoV S1 subunit, showed efficient neutralizing activity against the wild-type MERS-CoV strain EMC/2012, with a half maximal inhibitory concentration of 0.2 μg/mL. We concluded that a novel neutralizing epitope for MERS-CoV also resides on the NTD of the S protein, indicating that the NTD might be important during the viral infection process. Our findings have significant implications for further vaccine design and for the development of prophylactic and therapeutic monoclonal immunotherapies against MERS-CoV infection.
Emerging Microbes & Infections (2017) 6, e37; doi:10.1038/emi.2017.18; published online 24 May 2017
Keywords:
Introduction
Middle East respiratory syndrome coronavirus (MERS-CoV), a novel lethal human pathogen, has led to 1879 laboratory-confirmed cases of infection with an approximate fatality rate of 36% since its identification in Saudi Arabia in 2012.Citation1 The symptoms caused by MERS-CoV are similar to the symptoms of severe acute respiratory syndrome coronavirus (SARS-CoV), manifesting as an acute and severe lower respiratory illness with extrapulmonary involvement, including vomiting, diarrhea and renal failure.Citation2, Citation3 Infections have been confirmed in 27 countries, with the most cases occurring in the Middle East, followed by the recent outbreak in the Republic of Korea.Citation4, Citation5 Serological and virological surveys have indicated that dromedary camels are likely the major reservoir of MERS-CoV.Citation6, Citation7, Citation8 Although some human-to-human transmission cases, including contact with health care workers and family members, have been reported,Citation9, Citation10, Citation11, Citation12 it remains possible that the virus could acquire an adaptive mutation during repeated interspecies transmission events. Due to its potential threat, MERS-CoV has been listed as a Category C Priority Pathogen by the National Institute of Allergy and Infectious Diseases. However, neither licensed vaccines nor antiviral drugs against MERS-CoV have been approved for clinical use. Efficient countermeasures against this virus are urgently needed.
MERS-CoV is an enveloped positive-sense single-stranded RNA virus that belongs to the lineage C β-CoV genus. The MERS-CoV genome is ~30 kb and encodes the 5′-replicase structural protein (spike-envelope-membrane-nucleocapsid)-poly (A)-3′.Citation13, Citation14, Citation15 The highly glycosylated spike (S) protein mediates viral infections and is a primary determinant of cell tropism and pathogenesis. It assembles as a trimer on the viral particle surface and contains two functional subunits. The S1 subunit (residues 1–751) is mainly responsible for mediating viral particle attachment to the cell surface and is dependent on the dipeptidyl peptidase 4 (DPP4) receptor.Citation16, Citation17 The S2 subunit (residues 752–1353) facilitates the subsequent fusion of the virus with the host cell membrane ( and ). To deliver the viral nucleic acid into the host cell, the S1 subunit binds to the cellular receptor and triggers conformational changes in the S2 subunit, which then inserts its fusion peptide into the target cell membrane to form a six-helix bundle fusion core that prepares the viral and cell membranes for fusion.Citation19
Figure 1 Schematic diagram and analogous three-dimensional (3D) structure of the MERS-CoV S protein. (A) Amino acid sequences of the recombinant proteins (rNTD, rRBD, rS1, rS2 and rS) evaluated in this study. The NTD region of focus in this study is indicated by a red triangle. (B) The 3D structure of the MERS-CoV S protein was predicted using PyMOL, and the side view or transverse view is shown based on the trimeric S structure of HKU1.Citation18 N-terminal domain (NTD), RBD, S2 and the 367–606 and 606–751 aa regions are colored in light blue, dark blue, orange, red and green, respectively.
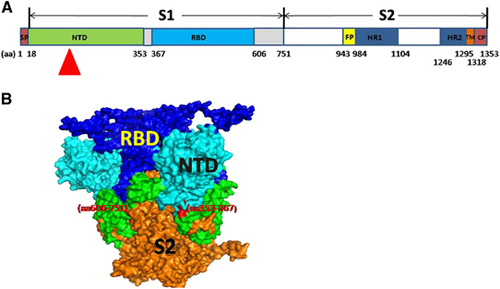
The receptor binding domain (RBD) of the S1 subunit serves as a determinant region for the production of MERS-CoV neutralizing antibodiesCitation20, Citation21, Citation22, Citation23, Citation24, Citation25, Citation26, Citation27 and has been the target for the development of a number of promising MERS-CoV vaccine candidates.Citation27, Citation28, Citation29, Citation30, Citation31, Citation32 The MERS-CoV RBD maps to a 200–300 residue region spanning residues 358–588, 367–588, 367–606, 377–588 and 377–662, which are located in the S1 subunit C-terminal domain.Citation16, Citation17, Citation33, Citation34, Citation35, Citation36, Citation37, Citation38, Citation39, Citation40 The specific interaction between MERS-CoV RBD and its receptor DPP4 (also known as CD26) has also been revealed at the atomic level by structural analysis.Citation16, Citation17 Accumulating evidence shows that all the neutralizing antibodies with therapeutic potential against MERS-CoV interfere with the association of RBD with DPP4.Citation20, Citation21, Citation22, Citation23, Citation24, Citation25, Citation26, Citation27 In addition to the immunogenic RBD, the undefined N-terminal domain (NTD) accounting for a large portion of S1 may serve as a functional component of the MERS-CoV S1 subunit. NTD has been recognized as the receptor-binding site in several CoVs, including murine hepatitis virus.Citation41 In this study, we discovered a neutralizing mAb that specifically recognizes the MERS-CoV S protein NTD. This study highlights the importance of the region in the viral infection process, which may enable us to better understand the underlying neutralizing mechanism of natural MERS-CoV infection. These findings pave the way for MERS-CoV vaccine and immunotherapy development.
Materials and methods
Ethics statement
Female BALB/c mice aged six to eight weeks were used for mAb production. The animal studies were performed in strict compliance with the Guide for the Care and Use of Laboratory Animals of the People’s Republic of China. The study protocol was approved by the Committee on the Ethics of Animal Experiments of the Chinese Center for Disease Control and Prevention.
Cell lines, virus and reagents
Vero E6, 293FT and Huh7.5 cell lines were cultured in Dulbecco’s Modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin-streptomycin (P/S), 1% l-glutamine and 1% HEPES. Non-essential amino acids (1%) were added to Huh7.5 cell cultures. SP2/0 cells were grown in RPMI 1640 containing 20% FBS, 1% P/S, 1% l-glutamine and 1% HEPES. After fusion with immunized mouse spleen cells, the hybridomas were cultured in DMEM with 20% FBS, 2% hypoxanthine aminopterin thymidine (HAT) and 1% P/S. All cells were cultured at 37 °C with 5% CO2. All reagents were purchased from Gibco (Life Technologies, Grand Island, NY, USA), except for DMEM, which was obtained from HyClone (Life Technologies, South Logan, UT, USA).
The MERS-CoV (HCoV-EMC/2012) strain was kindly provided by Professor Ron Fouchier (Erasmus Medical Centre, Rotterdam, Netherlands). The plasmids PNL4-3.luc.R-E- (a plasmid encoding an envelope-defective and luciferase-expressing HIV-1 genome) and pVRC-MERS-S (a plasmid encoding the spike protein of MERS-CoV) used for the MERS-CoV pseudovirus package were developed in our laboratory.
The baculovirus-insect cell sf9-derived recombinant MERS-CoV (HCoV-EMC/2012) S protein including rS (amino acids (aa) 1–1297) (40069–V08B), rS2 (aa 726–1296) (40070–V08B) and rS1 (40069–V08H) (aa 1–725) was purchased from SinoBiological Inc. (Beijing, China). The recombinant RBD (rRBD) (aa 367–606) and NTD (rNTD) (aa 18–353) expressed in a baculovirus system were kindly provided by Professors Jinghua Yan and George F Gao (Institute of Microbiology, Chinese Academy of Science, Beijing, China). The mouse mAb Isotyping Kit was purchased from Pierce (Life Technologies). The HiTrap protein G columns, Protein A sensor chip, 10 × PBS-P buffer (pH 7.4) and 10 mM glycine-HCl (pH 1.5) were purchased from GE Healthcare (Pittsburgh, PA, USA).
Mouse immunization and mAb generation
Mice were immunized with 35 μg MERS-CoV rS combined with 150 μL Freund’s complete adjuvant (Sigma, St Louis, CA, USA) via subcutaneous immunization and boosted twice at 2-week intervals beginning three weeks after the initial immunization. The mice were killed 3 days after the last immunization, and their splenocytes were fused with mouse myeloma cells.
The mAbs were generated as previously described.Citation42 Cells collected from the spleens of killed animals were fused with cultured SP2/0 cells at a 10:1 ratio in the presence of PEG1450 (Sigma). HAT selection medium was used for the fused hybridoma cultures. After a 2-week incubation, the positive hybridomas were selected using rS-coated ELISA, and the positive clones were subjected to limited dilutions and downstream validation. For large-scale mAb production, ascites fluid from mice inoculated with the hybridomas was collected and purified using a caprylic acid-ammonium sulfate precipitation assay and HiTrap Protein G columns with the AKTA system (GE Healthcare). Isotype classification of the purified mAbs was performed using the Pierce Rapid ELISA Mouse mAb Isotyping Kit (Pierce, Rockford, IL, USA) according to the manufacturer’s instructions.
ELISA assay
To precisely determine the binding regions of the mAbs targeting the S protein of MERS-CoV, truncated S fragments (rS1, rS2, rRBD and rNTD) and a panel of 43 peptides (Supplementary Table S1) spanning the entire MERS-CoV NTD were used as coating antigens in an indirect ELISA. Briefly, 96-well ELISA plates (Corning, Shanghai, China, Asia) were coated with the recombinant proteins (1 μg/mL) or 18 aa peptides (2.5 μg/mL) overnight at 4 °C. After blocking, the mAbs were added to the wells and incubated for 1.5 h at 37 °C. The plates were then incubated with HRP-conjugated secondary antibody for 1 h at 37 °C, and the binding activity was determined at 450 nm using a plate reader (Multiskan MK3). An anti-influenza virus N9-specific mAb was used in all assays as an unrelated negative control, and pre-fusion mouse serum was used as a positive control.
Affinity measurement by Biacore
The binding kinetics and affinity of the mAb 5F9 to purified rNTD were analyzed by surface plasmon resonance (SPR) (Biacore T200, GE Healthcare). The mAb 5F9 was immobilized to a Protein A sensor chip via binding to the Fc region in 1 × PBS-P+buffer (pH 7.4). Different concentrations of purified rNTD (60, 30, 15, 7.5 and 3.25 nM) were run at a flow rate of 10 μL/min in PBS-P+buffer. The surface was regenerated with 10 mM glycine-HCl (pH 1.5). Sensorgrams were fit with a 1:1 binding model using BIA Evaluation software (GE Healthcare).
Western blot analysis
Purified MERS-CoV rS and rNTD were analyzed on 12% SDS–polyacrylamide gels supplemented with 2 × SDS after boiling for 15 min. The denatured proteins were then transferred to a nitrocellulose membrane, blocked with 5% non-fat milk in PBST for 1 h at room temperature, and probed overnight with 1 μg/mL mAb and pre-fusion mouse serum (1:10 000 dilution) at 4°C. The membranes were incubated with DyLight 800 IgG secondary goat anti-mouse IgG (LI-COR Biosciences, NE, USA) and scanned using the Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA).
Neutralization test
The neutralizing activity of the selected mAbs was initially determined using pseudoviruses as described previously.Citation28 Briefly, 293FT cells were co-transfected with the plasmids PNL4-3.luc.R-E- and pVRC-MERS-S, and culture supernatant containing sufficient pseudotyped MERS-CoV was collected 48–72 h post-transfection. Subsequently, DPP4-expressing Huh7.5 cells, upon reaching a density of 5 × 103/well in a 96-well plate, were infected with 200 TCID50 (50% tissue culture infective dose) pseudovirus MERS-CoV in the presence or absence of mAbs. The culture medium was renewed with fresh medium containing 2% FBS 18–20 h post-infection, and luciferase activity was determined after an additional 48 h incubation using an Infinite M1000 illuminometer.
The neutralizing activity of the mAbs against live MERS-CoV was also determined in DPP4-expressing Vero E6 cells. Upon reaching a density of 5 × 104/well in a 12-well plate, cell monolayers were infected with 30–35 plaque-forming units (PFU) of live virus in the presence or absence of the mAb. After three days of incubation at 37°C, the inhibitory capacity of the mAbs was assessed by determining the numbers of PFU compared with the potent MERS-CoV anti-RBD and anti-N9 mAbs. All experiments associated with live MERS-CoV were conducted in a BSL-3 laboratory at the National Institute of Viral Diseases Control and Prevention, China CDC.
Results
Characterization of the mAbs
To identify novel neutralizing epitopes on the MERS-CoV S protein, a panel of mAbs targeting the S protein (aa 1–1297) was generated using a traditional hybridoma fusion protocol.Citation42 Nine rS-ELISA binding mAbs covering the RBD, the NTD and outside these two regions were selected from 37 clones. As shown in and , all mAbs bound strongly to sf9-derived rS (aa 1–1297) at a concentration of 1 μg/mL. Five of these mAbs (1F4, 3A2, 4A6, 4F8 and 4C5) bound strongly to rRBD (aa 367–606), while one (1B5) and three mAbs (5F9, 2B4 and 7D7) bound to rS2 (aa 753–1353) and rNTD (aa 18–353), respectively.
Figure 2 The isolated mAbs exhibited different binding affinities and neutralizing potencies to MERS-CoV. (A) These mAbs (5F9, 2B4, 7D7, 1F4, 3A2, 4A6, 4F8, 4C5 and 1B5) were mapped to different regions of the MERS-CoV S protein using ELISA. rS, rS1, rS2, rRBD and rNTD were used at a concentration of 1 μg/mL. (B) Inhibition of pseudovirus MERS-CoV entry into Huh7.5 cells by the isolated mAbs. Each of the selected mAbs (1 μg/mL), anti-N9 mAb (1 μg/mL) and anti-serum (1:10 000) were evaluated for their neutralizing activity against pseudotyped MERS-CoV after incubation with 293FT cell surface-expressing MERS-CoV S proteins. Neutralizing activity was defined as occurring when the inhibition percentage exceeded 50%. The results shown are representative of three independent experiments. N-terminal domain, NTD.
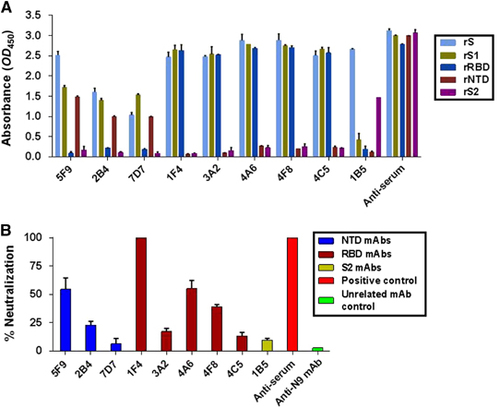
Next, we assessed the neutralizing activities of the nine selected mAbs against 200 TCID50 MERS-CoV pseudoviruses. As shown in , only mAbs 5F9, 1F4, 4A6 and 4F8 exhibited greater than 50% neutralization potency at 1 μg/mL compared to pre-fusion mouse serum (1:10 000 dilution). The rRBD-binding mAb 1F4 showed the strongest neutralizing activity (~100%); another two mAbs (4A6 and 4F8) and one novel rNTD-binding mAb (5F9) exhibited efficient neutralizing activity (over 50%). The other five mAbs, including 1B5 (rS2-binding mAb), exhibited less than 20% neutralization activity. The mAb 5F9 was therefore selected for further characterization and evaluation.
Binding efficiency of 5F9 to rNTD of MERS-CoV
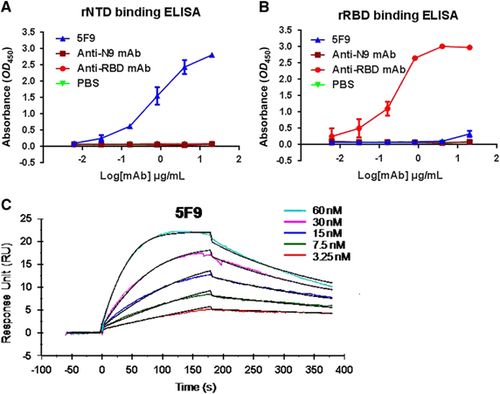
To further determine the binding efficiency of mAb 5F9, samples were diluted five-fold and detected using an MERS-CoV rNTD-coated ELISA. The results showed that 5F9 had a high binding affinity for rNTD, with an EC50 of approximately 0.85 μg/mL (). Meanwhile, 5F9 exhibited no cross-reaction with rRBD, even at a concentration of 4 μg/mL (). The anti-N9 mAb was evaluated in parallel as an unrelated negative control.
Next, the binding rate constant of the mAb 5F9 to rNTD was determined by SPR. As shown in , 5F9 showed nanomolar affinity for the rNTD (equilibrium dissociation constant (KD) equivalent to 5.42 nM).
Figure 3 The 5F9 antibody bound to the rNTD with high affinity. Recombinant proteins at 1 μg/mL were used to coat plates overnight at 4 °C, and each of the mAbs was serially diluted in PBS and assessed for binding affinity and specificity to MERS-CoV rNTD (A) and rRBD (B). (C) The binding kinetics between 5F9 and rNTD using SPR. The Ka and KD values were calculated from sensograms using five rNTD concentrations (60, 30, 15, 7.5 and 3.25 nM). The results shown are representative of three independent experiments. Anti-RBD, anti-N9 mAb and PBS were used as the positive, unrelated and blank controls, respectively. N-terminal domain, NTD.
Neutralizing efficiency of 5F9 to MERS-CoV
Previous results showed that 5F9 exhibited approximately 60% neutralizing activity toward pseudovirus MERS-CoV at a concentration of 1 μg/mL. We determined the lowest concentration of the mAb that can effectively inhibit pseudovirus MERS-CoV entry using two-fold dilutions of the mAbs. The results showed that the neutralizing activity of 5F9 was dose dependent, and the IC50 was approximately 0.24 μg/mL. The IC50 of anti-RBD mAb (0.02 μg/mL) was 10-fold lower than the IC50 of 5F9 (). Therefore, 5F9 showed efficient neutralizing activity and was further characterized.
Figure 4 Potent in vitro neutralization of MERS-CoV by 5F9 mAb. (A) Neutralization of pseudotyped MERS-CoV. DPP4-expressing Huh7.5 cells were cultured with 200 TCID50 pseudotyped MERS-CoV in the presence of serially diluted mAbs. The neutralizing percentage was calculated by measuring luciferase expression compared to the pseudovirus-infected cell control. (B) Neutralization of live viruses. Different concentrations of the mAbs were pre-cultured with the live viruses (HCoV-EMC) in Vero E6 cell monolayers. The neutralization percentage was evaluated by calculating the decrease in PFU number compared with the virus-infected control. (C) PFU images of viral infection in the presence of the mAbs on day 3. The images correspond to the neutralizing percentages in B. Approximately 30–35 PFU virus stocks (HCoV-EMC) were used to infect Vero E6 cells in a 12-well plate with or without mAbs. The results shown are representative of three independent experiments. Anti-RBD and anti-N9 mAbs were used as positive and unrelated controls, respectively.
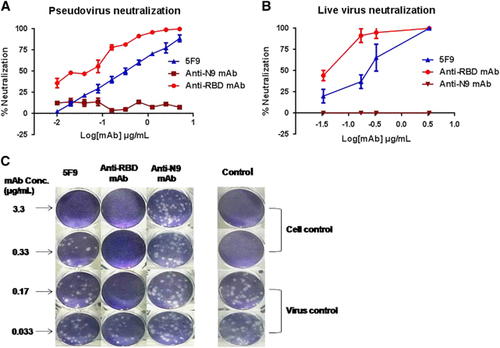
The neutralizing potency against live MERS-CoV (strain HCoV-EMC/2012) was also determined. As shown in , mAb 5F9 neutralized the live MERS-CoV infection, with the same IC50 of 0.2 μg/mL. Images of the reduced PFU formation, corresponding to the live MERS-CoV neutralizing percentages, are shown in . Together, the neutralization data indicate that MERS-CoV NTD may play an important role in the viral infection process.
Preliminary epitope analysis of the mAb 5F9
To further characterize the binding abilities of mAb 5F9 to the MERS-CoV NTD (aa 18–353), we employed a Western blot assay in which the rNTD was denatured to determine if the epitope was conformational or linear. The results showed that mAb 5F9 efficiently bound both the MERS-CoV rNTD (aa 18–353) and rS (). An 18 aa peptide ELISA was performed to locate the binding site of 5F9. The results showed that 5F9 bound very weakly to these peptides (OD450<0.5) (, Supplementary Table S1). These results suggest a possible linear binding epitope between 5F9 and the NTD, although the specific region was not clearly determined.
Figure 5 Binding abilities of 5F9 to denatured MERS-CoV rS, rNTD and synthetic peptides spanning the NTD. (A) Western blot analysis of mAb 5F9 bound to denatured rS and rNTD. Lane rS, 1 μg denatured rS protein (aa 1–1297); Lane rNTD, 1 μg denatured rNTD (aa 18–353). Pre-fusion mouse serum (anti-serum) was used as a positive control. (B) Binding of 5F9 to overlapping 18 aa peptides. P2 to P44 (18 aa peptides) correspond to numbers 2 to 44 in Supplementary Table S1. Each peptide was coated onto a 96-well plate at 250 ng/well, and the mAb was evaluated at a concentration of 1 μg/mL. The results shown are representative of three independent experiments. N-terminal domain, NTD.
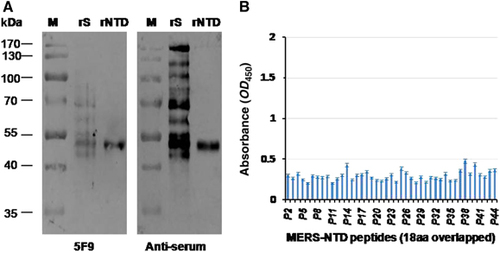
Table 1 Characterization of the mAbs against MERS-CoV in this study
Discussion
Several studies have demonstrated that most of the potent neutralizing mAbs against MERS-CoV derived from both human and murine origins target the RBD region of the S protein.Citation20, Citation21, Citation22, Citation23, Citation24, Citation25, Citation26, Citation27 In addition, these mAbs have shown promising therapeutic value in animal models.Citation23, Citation24, Citation26, Citation27 RBD-based vaccines have also elicited strong humoral immune responses using different immunization strategies.Citation27, Citation28, Citation29, Citation30, Citation31, Citation32 Our findings demonstrate that the mAb 5F9 antibody specifically targets the MERS-CoV NTD and exhibits efficient neutralizing activity against MERS-CoV, although its in vitro neutralizing potency (IC50 0.2 μg/mL) was approximately 10-fold lower than the potency of the RBD-targeting mAbs. Thus, this mAb warrants further development and evaluation in animal models to provide alternatives for MERS-CoV immunotherapy should the virus mutate and no longer remain susceptible to RBD-specific mAb treatment.
The surface S segment is the major CoV structural protein, and maintaining the quaternary nature of the S1–S2 complex is vital for viral infections. Recently, one study showed that SARS-CoV infection is affected by the inactive-to-active state transition of the S glycoprotein trimer. In addition, the loops connecting the NTD to the C-terminal domain (CTD) 1 and CTD1 to CTD2 were found to play key roles in the conformational switch.Citation43 Lassa virus (LASV) envelope glycoprotein complex (GPC) is another class I viral fusion protein that initiates infection by processing into GP1, GP2 and an unusual stable signal peptide (SSP). GP1 is responsible for receptor binding, while GP2 mediates the fusion of the virion with a cell membrane. Previous studies demonstrated that gaps in the glycan shield of LASV GPC permitted neutralizing antibodies to interact simultaneously with the N-terminal extension of GP1 and either the fusion loop or the T-loop of GP2, inhibiting the structural alterations required for fusion.Citation44 These discoveries showed that maintaining glycoprotein structural integrity during viral infection is necessary and that neutralizing mAbs targeting different surface glycoprotein epitopes can interfere with structural state switching. As shown in , structural docking of the MERS-CoV S protein revealed the NTD and C-terminal RBD of the S1 subunit, which form extensive quaternary interactions with each other. These features may imply that certain epitopes on the NTD of MERS-CoV S1 are structurally or functionally close to the ones on the RBD and that the mAbs targeting these epitopes can interfere with receptor binding or the post-fusion stage.
The glycan composition of class I envelope glycoproteins can shield a virus against the neutralizing effect of a mAb. A neutralizing mAb against the HIV-1 envelope glycoprotein was found to bind to the residues and glycans on gp41 and gp120, thereby precluding the CD4- and/or co-receptor-induced conformational changes required for membrane fusion.Citation45 The S proteins of the Coronaviridae family, consisting of four genera (α-, β-, γ- and δ-CoVs), are also class I envelope glycoproteins, and all use the S1 protein RBD or NTD to interact with cellular receptors or co-receptors for viral attachment.Citation46 To the best of our knowledge, most α-CoVs and some β-CoVs, including SARS-CoV, MERS-CoV and HCoV-HKU1, use protein moieties as their recognition receptors, and their main functional RBDs are located in the CTD of S1. In contrast, the RBDs of bovine CoV, human CoV OC43 and one avian γ-CoV are located in the NTD, and sugar moieties are the main receptors. More notably, the NTD and CTD of two α-CoVs, porcine epidemic diarrhea CoV and transmissible gastroenteritis, might both play vital roles in viral infection.Citation47, Citation48 The NTD of MERS-CoV is also glycosylated, and the neutralizing mAb 5F9 may bind to the glycan or other residues of the NTD, stabilizing the pre-fusion conformation of the S protein and the S1–S2 interaction, thereby precluding the DPP4- and/or co-receptor-induced conformational changes required for membrane fusion.
Continuing human infection with MERS-CoV is a global public health concern that highlights the need for efficient countermeasures. Currently, various vaccines and therapeutic antibodies against MERS-CoV are under development, and most have targeted the MERS-CoV RBD because it contains the most variable regions. Multiple evaluations of the full length S-based or truncated S1-based vaccines indicate that an increase in epitope range would be important for future MERS-CoV vaccine designs.Citation49, Citation50 Our study suggested that the NTD, which contains the neutralizing epitope, should be considered an immunogenic component. Thus, we propose that combining the most promising vaccine candidate with the NTD would not only improve the protective effects but also reduce the likelihood of possible resistance development via mutation.
Supplementary Table 1
Download MS Excel (24 KB)Acknowledgments
This work was supported by grants from the National Key Research and Development Program of China (2016YFD0500301, 2016YFC1200901, and 2016YFC1200200), the Megaproject for Infectious Disease Research of China (2016ZX10004001-003) and the General Program of National Natural Science Foundation of China 31570162. The funders played no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. The authors thank Dr. Jinghua Yan and Dr George F Gao (CAS Key Laboratory of Pathogenic Microbiology and Immunology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China) for providing reagents (rRBD and rNTD), and Dr Bart L Haagmans and Dr Ron A.M. Fouchier (Department of Viroscience, Erasmus Medical Center, Rotterdam, The Netherlands) for providing the HCoV-EMC/2012 strain.
Supplementary Information for this article can be found on the Emerging Microbes & Infections website (http://www.nature.com/emi)
Related Research Data
References
- Zaki AM, van Boheemen S, Bestebroer TMet al.Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 2012;367: 1814–1820.
- Chan JF, Lau SK, Woo PC.The emerging novel Middle East respiratory syndrome coronavirus: the 'knowns' and 'unknowns'. J Formos Med Assoc 2013;112: 372–381.
- Chan JF, Li KS, To KKet al.Is the discovery of the novel human betacoronavirus 2c EMC/2012 (HCoV-EMC) the beginning of another SARS-like pandemic? J Infect 2012;65: 477–489.
- Ki M.2015 MERS outbreak in Korea: hospital-to-hospital transmission. Epidemiol Health 2015;37: e2015033.
- Lee SS, Wong NS.Probable transmission chains of Middle East respiratory syndrome coronavirus and the multiple generations of secondary infection in South Korea. Int J Infect Dis 2015;38: 65–67.
- Briese T, Mishra N, Jain Ket al.Middle East respiratory syndrome coronavirus quasispecies that include homologues of human isolates revealed through whole-genome analysis and virus cultured from dromedary camels in Saudi Arabia. MBio 2014;5: e01146–01114.
- Haagmans BL, Al Dhahiry SH, Reusken CBet al.Middle East respiratory syndrome coronavirus in dromedary camels: an outbreak investigation. Lancet Infect Dis 2014;14: 140–145.
- Yusof MF, Eltahir YM, Serhan WSet al.Prevalence of Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels in Abu Dhabi Emirate, United Arab Emirates. Virus Genes 2015;50: 509–513.
- Health Protection Agency (HPA) UK Novel Coronavirus Investigation team.Evidence of person-to-person transmission within a family cluster of novel coronavirus infections, United Kingdom, February 2013. Euro Surveill 2013;18: 20427.
- Memish ZA, Zumla AI, Assiri A.Middle East respiratory syndrome coronavirus infections in health care workers. N Engl J Med 2013;369: 884–886.
- Assiri A, McGeer A, Perl TMet al.Hospital outbreak of Middle East respiratory syndrome coronavirus. N Engl J Med 2013;369: 407–416.
- Seo YB, Heo JY, Song M-Set al.Environmental contamination and viral shedding in MERS patients during MERS-CoV outbreak in South Korea. Clin Infect Dis 2015;62: 755–760.
- Cotten M, Lam TT, Watson SJet al.Full-genome deep sequencing and phylogenetic analysis of novel human betacoronavirus. Emerg Infect Dis 2013;19: 736–742B.
- Chan JF, Lau SK, To KKet al.Middle East respiratory syndrome coronavirus: another zoonotic betacoronavirus causing SARS-like disease. Clin Microbiol Rev 2015;28: 465–522.
- van Boheemen S, de Graaf M, Lauber Cet al.Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. MBio 2012;3: e00473–00412.
- Lu G, Hu Y, Wang Qet al.Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 2013;500: 227–231.
- Wang N, Shi X, Jiang Let al.Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res 2013;23: 986–993.
- Kirchdoerfer RN, Cottrell CA, Wang Net al.Pre-fusion structure of a human coronavirus spike protein. Nature 2016;531: 118–121.
- Millet JK, Whittaker GR.Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc Natl Acad Sci USA 2014;111: 15214–15219.
- Jiang L, Wang N, Zuo Tet al.Potent neutralization of MERS-CoV by human neutralizing monoclonal antibodies to the viral spike glycoprotein. Sci Transl Med 2014;6: 234ra259–234ra259.
- Ying T, Du L, Ju TWet al.Exceptionally potent neutralization of Middle East respiratory syndrome coronavirus by human monoclonal antibodies. J Virol 2014;88: 7796–7805.
- Tang X-C, Agnihothram SS, Jiao Yet al.Identification of human neutralizing antibodies against MERS-CoV and their role in virus adaptive evolution. Proc Natl Acad Sci USA 2014;111: E2018–E2026.
- Li Y, Wan Y, Liu Pet al.A humanized neutralizing antibody against MERS-CoV targeting the receptor-binding domain of the spike protein. Cell Res 2015;25: 1237–1249.
- Corti D, Zhao J, Pedotti Met al.Prophylactic and postexposure efficacy of a potent human monoclonal antibody against MERS coronavirus. Proc Natl Acad Sci USA 2015;112: 10473–10478.
- Du L, Zhao G, Yang Yet al.A conformation-dependent neutralizing monoclonal antibody specifically targeting receptor-binding domain in Middle East respiratory syndrome coronavirus spike protein. J Virol 2014;88: 7045–7053.
- Pascal KE, Coleman CM, Mujica AOet al.Pre-and postexposure efficacy of fully human antibodies against Spike protein in a novel humanized mouse model of MERS-CoV infection. Proc Natl Acad Sci USA 2015;112: 8738–8743.
- Wang L, Shi W, Joyce MGet al.Evaluation of candidate vaccine approaches for MERS-CoV. Nat Commun 2015;6: 7712.
- Lan J, Yao Y, Deng Yet al.Recombinant receptor binding domain protein induces partial protective immunity in rhesus macaques against Middle East respiratory syndrome coronavirus challenge. EBioMedicine 2015;2: 1438–1446.
- Du L, Kou Z, Ma Cet al.A truncated receptor-binding domain of MERS-CoV spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: implication for developing therapeutics and vaccines. PloS One 2013;8: e81587.
- Ma C, Li Y, Wang Let al.Intranasal vaccination with recombinant receptor-binding domain of MERS-CoV spike protein induces much stronger local mucosal immune responses than subcutaneous immunization: Implication for designing novel mucosal MERS vaccines. Vaccine 2014;32: 2100–2108.
- Tang J, Zhang N, Tao Xet al.Optimization of antigen dose for a receptor-binding domain-based subunit vaccine against MERS coronavirus. Hum Vaccin Immunother 2015;11: 1244–1250.
- Zhang N, Tang J, Lu Let al.Receptor-binding domain-based subunit vaccines against MERS-CoV. Virus Res 2015;202: 151–159.
- Du L, Zhao G, Kou Zet al.Identification of a receptor-binding domain in the S protein of the novel human coronavirus Middle East respiratory syndrome coronavirus as an essential target for vaccine development. J Virol 2013;87: 9939–9942.
- Jiang S, Lu L, Du Let al.A predicted receptor-binding and critical neutralizing domain in S protein of the novel human coronavirus HCoV-EMC. J Infect 2013;66: 464–466.
- Mou H, Raj VS, van Kuppeveld FJet al.The receptor binding domain of the new Middle East respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J Virol 2013;87: 9379–9383.
- Yang Y, Du L, Liu Cet al.Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus. Proc Natl Acad Sci USA 2014;111: 12516–12521.
- Raj VS, Mou H, Smits SLet al.Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013;495: 251–254.
- Ma C, Wang L, Tao Xet al.Searching for an ideal vaccine candidate among different MERS coronavirus receptor-binding fragments—the importance of immunofocusing in subunit vaccine design. Vaccine 2014;32: 6170–6176.
- Jiang S, Lu L, Du Let al.Putative conformations of the receptor-binding domain in S protein of hCoV-EMC in complex with its receptor dipeptidyl peptidase-4. J Infect 2013;67: 156.
- Chen Y, Rajashankar KR, Yang Yet al.Crystal structure of the receptor-binding domain from newly emerged Middle East respiratory syndrome coronavirus. J Virol 2013;87: 10777–10783.
- Peng G, Sun D, Rajashankar KRet al.Crystal structure of mouse coronavirus receptor-binding domain complexed with its murine receptor. Proc Natl Acad Sci USA 2011;108: 10696–10701.
- He Y, Lu H, Siddiqui Pet al.Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J Immunol 2005;174: 4908–4915.
- Gui M, Song W, Zhou Het al.Cryo-electron microscopy structures of the SARS-CoV spike glycoprotein reveal a prerequisite conformational state for receptor binding. Cell Res 2017;27: 119–129.
- Robinson JE, Hastie KM, Cross RWet al.Most neutralizing human monoclonal antibodies target novel epitopes requiring both Lassa virus glycoprotein subunits. Nat Commun 2016;7: 11544.
- Blattner C, Lee JH, Sliepen Ket al.Structural delineation of a quaternary, cleavage-dependent epitope at the gp41-gp120 interface on intact HIV-1 Env trimers. Immunity 2014;40: 669–680.
- King AM, Adams MJ, Lefkowitz EJ. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses.Elsevier, Netherland.2011;9: 193–210.
- Deng F, Ye G, Liu Qet al.Identification and comparison of receptor binding characteristics of the spike protein of two porcine epidemic diarrhea virus strains. Viruses 2016;8: 55.
- Schultze B, Krempl C, Ballesteros MLet al.Transmissible gastroenteritis coronavirus, but not the related porcine respiratory coronavirus, has a sialic acid (N-glycolylneuraminic acid) binding activity. J Virol 1996;70: 5634–5637.
- Modjarrad K.MERS-CoV vaccine candidates in development: The current landscape. Vaccine 2016;34: 2982–2987.
- Du L, Jiang S.Middle East respiratory syndrome: current status and future prospects for vaccine development. Expert Opin Biol Ther 2015;15: 1647–1651.