
Abstract
Previous findings of Middle East Respiratory Syndrome coronavirus (MERS-CoV)-related viruses in bats, and the ability of Tylonycteris-BatCoV HKU4 spike protein to utilize MERS-CoV receptor, human dipeptidyl peptidase 4 hDPP4, suggest a bat ancestral origin of MERS-CoV. We developed 12 primary bat cell lines from seven bat species, including Tylonycteris pachypus, Pipistrellus abramus and Rhinolophus sinicus (hosts of Tylonycteris-BatCoV HKU4, Pipistrellus-BatCoV HKU5, and SARS-related-CoV respectively), and tested their susceptibilities to MERS-CoVs, SARS-CoV, and human coronavirus 229E (HCoV-229E). Five cell lines, including P. abramus and R. sinicus but not T. pachypus cells, were susceptible to human MERS-CoV EMC/2012. However, three tested camel MERS-CoV strains showed different infectivities, with only two strains capable of infecting three and one cell lines respectively. SARS-CoV can only replicate in R. sinicus cells, while HCoV-229E cannot replicate in any bat cells. Bat dipeptidyl peptidase 4 (DPP4) sequences were closely related to those of human and non-human primates but distinct from dromedary DPP4 sequence. Critical residues for binding to MERS-CoV spike protein were mostly conserved in bat DPP4. DPP4 was expressed in the five bat cells susceptible to MERS-CoV, with significantly higher mRNA expression levels than those in non-susceptible cells (P = 0.0174), supporting that DPP4 expression is critical for MERS-CoV infection in bats. However, overexpression of T. pachypus DPP4 failed to confer MERS-CoV susceptibility in T. pachypus cells, suggesting other cellular factors in determining viral replication. The broad cellular tropism of MERS-CoV should prompt further exploration of host diversity of related viruses to identify its ancestral origin.
These authors contributed equally: Susanna K. P. Lau, Rachel Y. Y. Fan, Hayes K. H. Luk
Introduction
Coronaviruses (CoVs) are important pathogens in animals and humans, responsible for a variety of respiratory, enteric, hepatic, and neurological diseases. They are now classified into four genera, Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus, with Betacoronavirus further divided into lineages A to DCitation1–Citation4. Humans are infected by six CoVs, including human CoV 229E (HCoV-229E) and human CoV NL63 (HCoV-NL63) belonging to Alphacoronavirus; human CoV OC43 (HCoV-OC43) and human CoV HKU1 (HCoV HKU1) belonging to Betacoronavirus lineage A; Severe Acute Respiratory Syndrome-related CoV (SARSr-CoV) belonging to Betacoronavirus lineage B; and Middle East Respiratory Syndrome CoV (MERS-CoV) belonging to Betacoronavirus lineage CCitation5–Citation12. The emergence potential of CoVs is believed to be related to their tendency for mutation and recombination, allowing the generation of new viruses being able to adapt to new hostsCitation3,Citation13–Citation18.
Bats are an important reservoir of alphacoronaviruses and betacoronaviruses, which may jump to other animals or humans to cause new epidemicsCitation2,Citation19. For example, SARS-CoV is likely a recombinant virus originated from horseshoe bats as the primary reservoir and palm civet as the intermediate hostCitation16,Citation20–Citation25. Since the SARS epidemic, numerous other novel CoVs from humans or animals have been discoveredCitation2,Citation26–Citation30, allowing a better understanding of the evolutionary origin of emerging CoVs.
Although dromedary camels are now known to be the immediate animal source of the recent MERS epidemic, the evolutionary origin of MERS-CoV remains obscureCitation31,Citation32. When the virus was first discovered, it was found to be closely related to Tylonycteris bat CoV HKU4 (Ty-BatCoV HKU4) and Pipistrellus bat CoV HKU5 (Pi-BatCoV HKU5) previously discovered in lesser bamboo bat (Tylonycteris pachypus) and Japanese pipistrelle (Pipistrellus abramus) respectively in Hong KongCitation3,Citation11,Citation33,Citation34. Five other lineage C betacoronaviruses closely related to MERS-CoV were subsequently detected in bats, including BtVs-BetaCoV/SC2013, Hypsugo BatCoV HKU25 from China bats, and coronavirus Neoromicia/PML-PHE1/RSA/2011 (NeoCoV), BtCoVNeo5038/KZN/RSA/2015, and BatCoV PREDICT/PDF-2180 from African batsCitation35–Citation39. Besides bats, a lineage C betacoronavirus, Erinaceus CoV VMC/DEU, subsequently defined as a novel species, Hedgehog coronavirus 1, has also been discovered in European hedgehogs, a group of animals being phylogenetically closely related to batsCitation40. While none of these animal viruses represent the immediate ancestor of MERS-CoV, the spike protein of Ty-BatCoV HKU4 was most closely related to that of MERS-CoV, and was shown to utilize the MERS-CoV receptor, human dipeptidyl peptidase 4 (hDPP4) or CD26, for cell entryCitation41,Citation42. Previous studies have also shown that MERS-CoV was able to infect bat cell lines and Jamaican fruit batsCitation43,Citation44. These findings suggest that bats may be the primary host of the ancestor of MERS-CoV.
Although MERS-CoV has been shown to replicate in various animal cell lines including bat cellsCitation43–Citation48, the broad tissue tropism was mainly demonstrated using type strain EMC/2012, and no comparison was made between different MERS-CoV strains. Moreover, cells from the bat hosts of MERS-CoV-related viruses, such as T. pachypus and P. abramus which harbor Ty-BatCoV HKU4 and Pi-BatCoV HKU5, respectively, were not included in previous studies, which may be due to the geographical limitation of these bat species. To better understand the replicative ability of MERS-CoV in bat cells, which may provide clues on the origin of MERS-CoV, we developed diverse primary bat cell lines from different bat species, including Rhinolophus sinicus (the host of SARSr-BatCoV) and T. pachypus (the host of Ty-BatCoV HKU4), and tested their susceptibilities to infection by different strains of MERS-CoV, SARS-CoV, and HCoV-229E. The DPP4 mRNA sequences of six bat species and their expression in bat cells were determined to correlate with viral replication results. Our findings showed differential cell tropism between different strains of MERS-CoV, SARS-CoV, and HCoV-229E, which offers insights to the origin of MERS-CoV.
Results
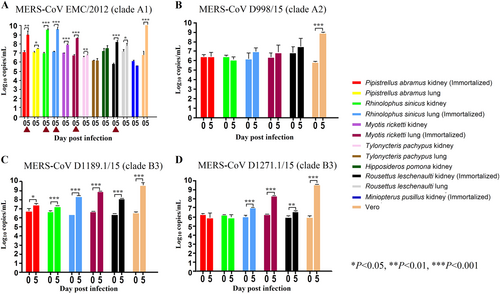
Five of 12 tested bat cell lines are susceptible to MERS-CoV EMC/2012 infection
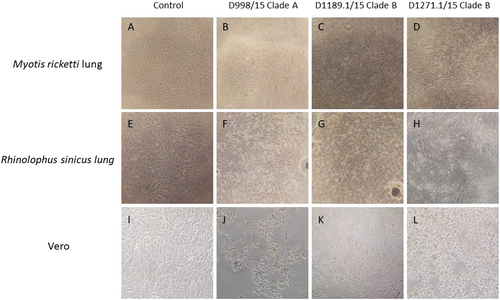
Since lineage C betacoronaviruses closely related to MERS-CoV were detected in bats, we developed 12 diverse primary bat cell lines from seven different bat species, including Hipposideros pomona, Miniopterus pusillus, Myotis ricketti, Pipistrellus abramus (the host of Pi-BatCoV HKU5), Rhinolophus sinicus (the host of SARSr-BatCoV and Rs-BatCoV HKU2), Tylonycteris pachypus (the host of Ty-BatCoV HKU4), Rousettus leschenaultii (the host of many viruses including Ro-BatCoV HKU9), which were subjected to infection with MERS-CoV at multiplicity of infection (MOI) of 1. Viral titers were determined by RT-qPCR on day 5 p.i. Five of the 12 cell lines (M. ricketti lung, P. abramus kidney, R. sinicus kidney and lung, and R. leschenaulti kidney cells) and Vero cells propagated MERS-CoV with at least one log10 increase in viral load. The highest increase in viral load was observed in R. sinicus kidney and lung cells, which was comparable with that observed in Vero cells (Fig. ). Cytopathic effects (CPEs) were observed in infected M. ricketti lung and R. sinicus lung cells with rounding of cells (Fig. ). The infectivities of the viruses from culture supernatants were confirmed by passage in Vero cells with CPE. H. pomona kidney, M. pusillus kidney, M. ricketti kidney, P. abramus lung, R. leschenaulti lung, and T. pachypus kidney and lung cells did not support MERS-CoV infection.
The 12 bat cell lines (PAK: Pipistrellus abramus kidney, PAL Pipistrellus abramus lung, RSK: Rhinolophus sinicus kidney, RSL: Rhinolophus sinicus lung, MRK: Myotis ricketti kidney, MRL: Myotis ricketti lung, TPK: Tylonycteris pachypus kidney, TPL: Tylonycteris pachypus lung, HPK: Hipposideros pomona kidney, RLK: Rousettus leschenaultii kidney, RLL: Rousettus leschenaultii lung, MPK: Miniopterus pusillus kidney) and Vero cells were subject to infection by MERS-CoV EMC/2012 (belonging to clade A1) with MOI of 1 (a). Culture supernatants were harvested at day 0 and 5 postinfection. Viral titers were determined by real-time quantitative RT-PCR. Viral load was expressed as log10 copies/mL. Error bars indicate the standard deviation of triplicate samples. The five bat cell lines susceptible to MERS-CoV EMC/2012 infection with ≥ 1 log10 increase in viral load at day 5 were marked with red triangles. They were subject to infection by three other MERS-CoV strains isolated from camels in Dubai, D998/15 (belonging to clade A2) (b), D1189.1/15 (c) and D1271.1/15 (d) (belonging to clade B3) (b). Different MERS-CoV strains displayed different infectivities in these five bat cells. (*P < 0.05; **P < 0.01; ***P < 0.001)
CPE was compared between Myotis ricketti lung (immortalized) cells that were uninfected (control) (a), and infected with Dubai camel MERS strains D998/15 (b), D1189.1/15 (c) & D1271.1/15 (d). CPE was compared between Rhinolophus sinicus lung (immortalized) cells that were uninfected (control) (e), and infected with Dubai camel MERS strains D998/15 (f), D1189.1/15 (g), and D1271.1/15 (h). CPE was compared between Vero cells that were uninfected (control) (i), and infected with Dubai camel MERS strains D998/15 (g), D1189.1/15 (k), and D1271.1/15 (l)
Different MERS-CoV strains displayed different infectivities on bat cells
MERS-CoVs are currently classified into three major clades, clade A, B, and C, which were further divided into subclades A1-A2, B1-B6, C1, and non-C1Citation49–Citation52. To test if different MERS-CoV strains may show similar infectivities on bat cells, the five bat cells (M. ricketti lung, P. abramus kidney, R. sinicus kidney and lung, and R. leschenaulti kidney cells), which were susceptible to MERS-CoV EMC/2012 (belonging to clade A1), were subjected to infection by three other MERS-CoV strains isolated from camels in Dubai, D998/15 (belonging to clade A2, but with spike gene belonging to clade C), and D1189.1/15 and D1271.1/15 (both belonging to clade B3)Citation51. Among the five challenged bat cells, M. ricketti lung supported infection by both D1189.1/15 and D1271.1/15; while R. sinicus lung and R. leschenaulti kidney supported infection by D1189.1/15, with at least one log10 increase in viral load. None of the cells supported infection by D998/15 (Fig. ).
SARS-CoV can replicate in R. sinicus cells
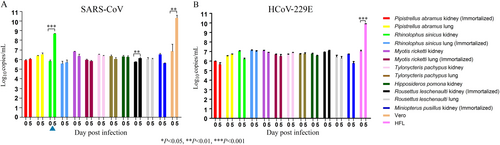
Since Chinese horseshoe bats are the major reservoir of SARS-related-CoVs, we also tested the 12 bat cell lines, including R. sinicus cells, for susceptibility to a clinical strain of SARS-CoV. SARS-CoV strain HKU-39849 can replicate in R. sinicus kidney but not lung cells, with at least one log10 increase in viral load (Fig. ). The other bat cells did not support SARS-CoV infection.
The 12 bat cell lines (PAK: Pipistrellus abramus kidney, PAL Pipistrellus abramus lung, RSK: Rhinolophus sinicus kidney, RSL: Rhinolophus sinicus lung, MRK: Myotis ricketti kidney, MRL: Myotis ricketti lung, TPK: Tylonycteris pachypus kidney, TPL: Tylonycteris pachypus lung, HPK: Hipposideros pomona kidney, RLK: Rousettus leschenaultii kidney, RLL: Rousettus leschenaultii lung, MPK: Miniopterus pusillus kidney) and Vero/HFL cells were subject to infection by SARS-CoV with MOI of 1 (a) and HCoV-229E with MOI of 0.01(b). Culture supernatants were harvested at day 0 and 5 postinfection. Viral titers were determined by real-time quantitative RT-PCR. Viral load was expressed as log10 copies/mL. Error bars indicate the standard deviation of triplicate samples. Only RSK cells can support SARS-CoV infection with ≥ 1 log10 increase in viral load at day 5 (blue triangle) and none of the 12 bat cell lines support HCoV-229E infection. (*P < 0.05; **P < 0.01; ***P < 0.001)
HCoV-229E cannot replicate in bat cells
The 12 bat cells were also tested for susceptible to HCoV-229E infection. HCoV-229E strain ATCC VR-740, previously isolated from a man with upper respiratory illness, cannot replicate in any of the tested bat cells (Fig. ).
mRNA transcript sequence analysis and expression of DPP4 in bat cells
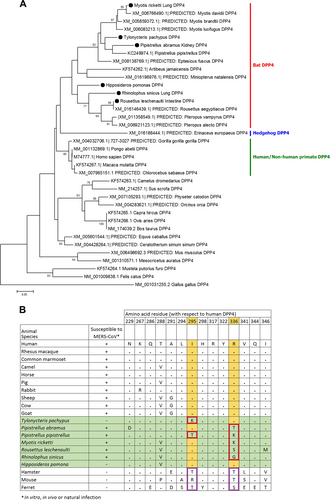
Partial DPP4 mRNA transcript sequences (corresponding to nt 688–1040 of hDPP4 which includes residue 229–346, where the critical residues for MERS-CoV spike protein binding are present) were determined for six of the seven bat species from which the 12 tested bat cells were developed. The sequence of M. pusillus was not determined, as RT-PCR for the DPP4 mRNA from bat cells was negative. Phylogenetic analysis showed that the bat DPP4 mRNA sequences formed a distinct cluster being closely related to sequences from human and non-human primates; while the dromedary camel DPP4 sequence was most closely related to that of wild boar (Fig. ). Previous studies have identified 14 critical residues in hDPP4 for binding of MERS-CoV spike proteinCitation53,Citation54. Upon multiple sequence alignment of the corresponding regions that contain these critical residues, most of the residues are conserved in the six bat DPP4 sequences in this study (Fig. ). Notably, T. pachypus DPP4 (Tp-DPP4) contains an I → K substitution at position 295 compared with hDPP4. On the other hand, residue R336 of hDPP4 was only conserved in T. pachypus and H. pomona, both not susceptible to MERS-CoV infection.
(a) The trees were constructed by Neighbor-Joining method using JTT substitution models and bootstrap values calculated from 1000 trees. Only bootstrap values >70% are shown. One hundred and twelve aa positions were included in the analyses. The scale bars represent 20 substitutions per site. Bat DPP4s that are sequenced in this study are labeled with black circles. Comparison of critical amino acid residues in DPP4 from different animal host for receptor binding in the region of residues 229–346 with respect to human DPP4 (b)
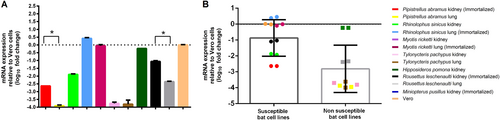
RT-qPCR of bat DPP4 mRNA in the corresponding bat cells was performed to determine the mRNA expression levels. Results showed that DPP4 mRNA was expressed in all the five bat cells that were susceptible to MERS-CoV infection, while it was also expressed in H. pomona kidney and R. leschenaulti lung cells which were not susceptible to MERS-CoV infection. The mRNA expression level in P. abramus kidney (susceptible to MERS-CoV) was significantly higher than that in its lung (non-susceptible) cells (P = 0.0185 by Student’s t test). Similarly, the mRNA expression level in R. leschenaulti kidney (susceptible to MERS-CoV) was significantly higher than that in its lung (non-susceptible) cells (P = 0.0009 by Student’s t test). Compared with bat cells that are non-susceptible, bat cells that are susceptible to MERS-CoV showed a significantly higher mRNA expression level (P = 0.0174 by Student’s t test) (Fig. ).
(a) mRNA levels of DPP4 were measured in various bat cells extracts by RT-qPCR and plotted relative to Vero cells, normalized by β-actin mRNA levels. The mRNA levels of DPP4 were compared between susceptible and non-susceptible bat cell lines (b). Statistical significance was assessed by Student’s t test P < 0.05
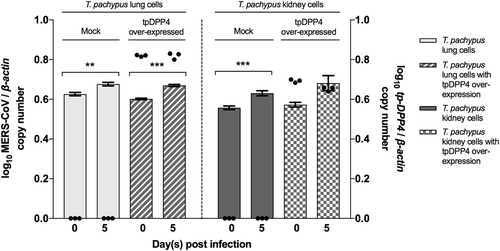
Overexpression of Tp-DPP4 does not confer MERS-CoV EMC/2012 susceptibility to T. pachypus cells
Since DPP4 was not expressed in T. pachypus cells while this bat species hosts Ty-BatCoV HKU4, a close relative of MERS-CoV which can utilize hDPP4 for cell entryCitation41,Citation42, we overexpressed Tp-DPP4 in T. pachypus lung and kidney cells for infection by MERS-CoV. While a slight increase (less than one log10) in viral replication was noted in both mock and Tp-DPP4-overexpressed cells on day 5 p.i., no significant difference was noted between mock and Tp-DPP4-overexpressed cells (Fig. ), suggesting that overexpression of Tp-DPP4 does not confer MERS-CoV susceptibility to T. pachypus cells.
Cells were infected with MERS-CoV at a multiplicity of infection (MOI) of 1 for 5 days. Determination of MERS-CoV viral load in supernatant (n = 3) by RT-qPCR with normalization to β-actin (represented by bar). Determination of tpDPP4 expression in cell lysates (n = 3) by RT-qPCR with normalization to beta-actin (represented by dot). (*P < 0.05, **P < 0.01, ***P < 0.001)
Discussion
The present study provides further support to the bat origin of MERS-CoV. Previous studies have shown that MERS-CoV can infect cell lines from different animals, including cells from various bat speciesCitation43–Citation47. However, bat cells from host species of MERS-CoV-related viruses, such as T. pachypus and P. abramus which harbor Ty-BatCoV HKU4 and Pi-BatCoV HKU5, respectively, were not included in these studies. In this study, MERS-CoV EMC/2012 was able to replicate in cells from four different bat genera/species belonging to three different bat families including Pteropodidae, Rhinolophidae, and Vespertilionidae. None of these bat cells were previously reported to support MERS-CoV replication. While lineage C betacoronaviruses have not been detected in bats outside the Vespertilionidae family, such diverse cellular tropism should prompt further studies to explore the host diversity of this group of CoVs and hence the possible evolutionary origin of MERS-CoV. In particular, the ability of MERS-CoV to replicate in cells from P. abramus, the host of Pi-BatCoV HKU5, may suggest that the ancestor of MERS-CoV could have originated from Pipistrellus-related bats. Interestingly, MERS-CoV showed the highest replicative ability in cells from R. sinicus. This suggests that MERS-CoV may potentially infect this bat species which is also the natural reservoir of SARSr-CoVs and animal origin of SARS-CoVCitation21. Nevertheless, three other MERS-CoV strains belonging to either clade A2 or B3 showed differential replicative abilities in the five bat cell lines that were susceptible to EMC/2012 strain (clade A1), with strain D998/15 (clade A2) unable to replicate in all five tested cell lines. Interestingly, it has been shown in a recent study that some clade C MERS-CoV strains from Nigeria and Burkina Faso replicated at a lower titre in Calu-3 cellsCitation52. This is in line with the present study that our clade A2 strain D998/15 (with its spike gene belonging to clade C), conferred less broad cellular tropism. Our findings suggest that different clades or strains of MERS-CoV may possess different cellular tropism and host range, while all MERS-CoVs can replicate well in the immune-deficient Vero cells.
The broader cellular tropism of MERS-CoV than SARS-CoV and HCoV-229E in bat cells may reflect the host diversity of lineage C betacoronaviruses in bats. Bats are now known to be the recent origin of at least two human CoVs, including SARS-CoV and HCoV-229E. SARS-CoV is likely a recombinant virus arising from horseshoe bats before it jumped to civet as the intermediate host and then humanCitation16,Citation24. SARSr-CoVs in bats were shown to utilize the SARS-CoV receptor, human angiotensin-converting enzyme 2 (hACE2), for cell entryCitation55. Similar to SARS-CoV, HCoV-229E is likely to have originated from bats. CoVs closely related to HCoV-229E have been detected in bats of the genus Hipposideros in AfricaCitation56. More recently, CoVs even closer to HCoV-229E were identified in dromedary camels in the Middle East, which were able to utilize the receptor of HCoV-229E, human aminopeptidase N (hAPN) for cell entryCitation57. This suggests that dromedary camels may have served as intermediate hosts for bat-to-human transmission of HCoV-229E, which may be analogous to the evolution of MERS-CoV. Recently, a CoV, closely related to HCoV-NL63 in most genome regions except the spike protein, was detected from a bat of the genus Triaenops in KenyaCitation58. However, further studies are required to identify even closer relatives of HCoV-NL63 to ascertain its possible bat origin. In contrast to SARSr-CoVs and HCoV-229E which were mainly found in horseshoe and roundleaf bats, respectively, a more diverse host range was observed in lineage C betacoronaviruses. Yet, the different bat species harboring lineage C betacoronaviruses all belonged to the family Vespertilionidae. Specifically, Ty-BatCoV HKU4, Pi-BatCoV HKU5, Hp-BatCoV HKU25, and BtVs-BetaCoV/SC2013 were detected in bats belonging to the genera, Tylonycteris, Pipistrellus, Hypsugo, and Verspetilio, respectively, in China, whereas NeoCoV and BatCoV PREDICT/PDF-2180 were detected in bats belonging to Neoromicia and Pipistrellus in AfricaCitation36,Citation37. In this study, SARS-CoV could only replicate in R. sinicus kidney cells but not other bat cells, which reflects its evolutionary origin from horseshoe bats. On the other hand, HCoV-229E was unable to replicate in any tested bat cells including cells from Hipposideros. This may suggest a relatively narrow host range of SARS-CoV and HCoV-229E in bats compared with lineage C betacoronaviruses including MERS-CoV.
The close phylogenetic relationship between bat and primate DPP4 sequences may reflect the replicative ability of MERS-CoV in bat cells. Moreover, cells that supported MERS-CoV replication showed significantly higher DPP4 mRNA expression than those that were non-susceptible to MERS-CoV, suggesting that cellular DPP4 expression is critical for viral infection. Since Ty-BatCoV HKU4 from T. pachypus was previously shown to be able to utilize hDPP4 for cell entryCitation41,Citation42, we expected MERS-CoV to be able to utilize Tp-DPP4 for receptor binding and infect T. pachypus cells. While the inability of MERS-CoV to replicate in T. pachypus cells may be partly explained by the lack of DPP4 expression, overexpression of Tp-DPP4 was unable to confer viral susceptibility. Since mRNA expression may not always correlate with cell surface expression of the receptor, further studies are required to study the receptor-binding interphase between MERS-CoV and Tp-DPP4 and whether other cellular factors may play a role in determining viral replicative abilities in T. pachypus cells, which may offer further insights into the evolutionary origin and mechanisms of interspecies transmission of MERS-CoV. Interestingly, one of the critical residues for binding to MERS-CoV spike protein, R336, was only conserved in T. pachypus and H. pomona DPP4 (cells from these two species did not support MERS-CoV replication), but not in the other four sequenced bat DPP4 (cells from these four species supported MERS-CoV replication). This suggests that this residue may not be critical for receptor binding. Binding and mutagenesis studies may help better understand the role of receptor-binding-interphase during viral evolution and interspecies jumping.
Materials and methods
Ethics statement
The collection of bat samples for developing bat cell lines was approved by the Committee on the Use of Live Animals for Teaching and Research, the University of Hong Kong, Hong Kong, China.
Cell lines
The bat cells, Vero (African green monkey kidney) cells, and HFL (human embryonic lung fibroblast) cells used in this study are described in Table . For development of primary bat cell lines, bats were captured in Hong Kong and euthanized before dissection for recovery of cells from organs aseptically. Briefly, the kidney and lung tissue were rinsed with cold PBS and cut into small pieces. Cold 0.25% trypsin-EDTA was added to the tissues and incubated at 4 °C overnight. Tissues were then incubated at 37 °C on shaking platform for 30 min. Supernatants were filtered through cell strainers to remove large pieces of tissuesCitation59. Bat cells were harvested by spinning down the supernatant at 1200 rpm for 8 min and were grown in DMEM/F12 supplemented with 15% FBS. Vero and HFL cells were grown in MEM supplemented with 10% FBS. All cells were incubated at 37 °C with 5% CO2.
Cell lines used in the present study
Virus isolates
MERS-CoV strain EMC/2012 was provided by Fouchier and Zaki et al.Citation11. MERS-CoV strains D998/15, D1189.1/15, and D1271.1/15 were isolated from dromedary camels in DubaiCitation51. SARS-CoV strain HKU-39849 was isolated from the brother-in-law of the index patient in Hong Kong during the SARS epidemicCitation8. The HCoV-229E strain ATCC VR-740 was used. MERS-CoV isolates and SARS-CoV were propagated in Vero cells at MOI of 0.01 in MEM supplemented with 1% FBS. HCoV-229E was propagated in HFL.
Infection of bat cell lines
Viral titers were determined as median tissue culture infective dose (TCID50) per ml in confluent cells in 96-well microtiter plates. Cells were seeded onto 24-well tissue culture plates, at 2 × 105 cells per well with the respective medium and incubated at 37 °C and 5% CO2 for 24 h prior to experiment. Cells were washed once with PBS and inoculated with 1 MOI of MERS-CoV or SARS-CoV, or 0.01 MOI of 229E for 1 h. After 1 h of viral adsorption, the supernatant was removed and cells were washed twice with PBS. The cells were maintained in MEM supplemented with 1% FBS for Vero and HFL cells and DMEM/F12 supplemented with 1% FBS for bat cells, before further incubation for 5 days.
Viral replication studies
To study viral replication efficiency, progeny viruses from cell culture supernatants collected at 5 days postinfection (p.i.) were subjected to reverse transcription-quantitative PCR (RT-qPCR) according to our previous protocolsCitation60. Briefly, total RNA extracted from cell culture supernatants with QIAsymphony DSP Virus/Pathogen Mini Kit (Qiagen, Hilden, Germany) was reverse transcribed and amplified with MERS-CoV primers (forward primer 5′-CAAAACCTTCCCTAAGAAGGAAAAG-3′ reverse primer 5′-GCTCCTTTGGAGGTTCAGACAT-3′), SARS-CoV primers (forward 5′- ACCAGAATGGAGGACGCAATG-3′ reverse 5′-GCTGTGAACCAAGACGCAGTATTAT-3′) and HCoV-229E primers (forward 5’-CAGTCAAATGGGCTGATGCA-3′ reverse 5′-AAAGGGCTATAAAGAGAATAAGGTATTCT-3′) using real-time one-step quantitative RT-PCR assay as described previously with modificationsCitation45,Citation60. Probes for MERS-CoV [5′-(FAM)ACAAAAGGCACCAAAAGAAGAATCAACAGACC(BHQ1)-3′], SARS-CoV [5′-(FAM)ACCCCAAGGTTTACCC(NFQ)-3′] and HCoV-229E [5′-(FAM)CCCTGACGACCACGTTGTGGTTCA(BHQ1)-3′] were used (Table ). Reactions were first incubated at 50 °C for 30 min, followed by 95 °C for 2 min, and were then thermal cycled for 50 cycles (95 °C for 15 s, 55 °C for 30 s). A series of 10 log10 dilutions equivalent to 1 × 101 to 1 × 1010 copies per reaction mixture were prepared to generate calibration curves and were run in parallel with the test samples. All experiments were performed in triplicate.
Primers used for RT-qPCR in this study
Amplification and sequencing of partial bat DPP4 mRNA transcripts
Total RNA was extracted from bat cell lysates using RNeasy Mini Spin Column (QIAgen). cDNA was PCR amplified with primers, 5′-GTCACCAGAGGGTCATAAA-3′ and 5′-CCACTTCCTCTGCCATCAAA-3′. The PCR mixture (25 μl) contained cDNA, PCR buffer, 200 μM (each) dNTPs, and 1.0 U Iproof Polymerase (Bio-Rad, Hercules, CA, USA). The mixtures were amplified for 40 cycles of 98 °C for 10 s, 55 °C for 30 s, and 72 °C for 72 s and a final extension at 72 °C for 10 min in an automated thermal cycler (Applied Biosystems, Foster City, CA, USA). RT-PCR products were gel purified using QIAquick gel extraction kit (Qiagen), and sequenced with an ABI Prism 3700 DNA Analyzer (Applied Biosystems). The sequences obtained were compared with sequences of DPP4 genes in GenBank database. Phylogenetic tree construction was performed based on an amino acid alignment of partial DPP4 sequences (corresponding to residue 229–346 of hDPP4) using the Neighbor-Joining method with JTT model by MEGA 6.0, with bootstrap values calculated from 1000 trees.
DPP4 expression analysis
To study DPP4 expression profiles in different bat cell lines, cell lysates were collected for total RNA extraction using RNeasy Mini Spin Column (QIAgen). RNA was eluted in 50 μl of RNase-free water and was used as a template for one-step RT-qPCR with SuperScript III platinum One-step qRT-PCR system (Invitrogen, Carlsbad, CA, USA). RT-qPCR assays were performed using conserved primers designed by multiple alignment of available bat DPP4 gene sequences (Table ), using β-actin for normalization. cDNA was amplified in a LightCycler 480 (Roche, Basel, Switzerland) with 25 μl reaction mixtures containing 2 × reaction mix, 5 μl RNA, 25 μM ROX reference dye, 50 μM primers, and 10 μM probe at 50 °C for 30 min, then 95 °C for 2 min followed by 50 cycles of denaturation, annealing and extension. Experiments were performed in triplicates, and results were expressed as the mean expression level of DPP4/β-actin. The relative expression between different bat cells was then calculated by the ΔΔCt method.
tpDPP4 overexpression in T. pachypus cells
T. pachypus DPP4 (tpDPP4) sequence was cloned into pLenti7.3/V5-TOPO vector (Invitrogen). The construct was transfected into 293FT cells together with ViraPower Packaging Mix (Invitrogen) using Lipofectamine 2000 (Life Technologies, Carlsbad, CA, USA). Lentivirus was harvested from the supernatant and concentrated using PEG-it (System Biosciences, Palo Alto, USA). T. pachypus cells resistant to MERS-CoV were transduced using the concentrated lentivirus for tpDPP4 overexpression and were subsequently subjected to inoculation with 1 MOI of MERS-CoV.
Nucleotide sequence accession numbers
The nucleotide sequences of bat DPP4 obtained from this study have been deposited in the GenBank sequence database under accession numbers MH345671-MH345676.
Acknowledgements
We thank C.T. Shek, Agriculture, Fisheries, and Conservation Department, the Government of the HKSAR, for capture of bats for bat cell development and expert opinion. This work is partly supported by Theme-Based Research Scheme (T11-707/15-R) and Research Grant Council Grant, University Grant Council; Health and Medical Research Fund of the Food and Health Bureau of HKSAR; University Development Fund and Special Research Achievement Award, The University of Hong Kong; Consultancy Service for Enhancing Laboratory Surveillance of Emerging Infectious Disease for the HKSAR Department of Health; The Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, the Ministry of Education of China; National Science and Technology Major Project of China (grant number 2012ZX10004213). Views expressed in this paper are those of the authors only, and may not represent the opinion of the AFCD or the Government of the HKSAR.
Author contributions
S.K.P.L. and P.C.Y.W. conceived the study and designed the experiments. R.Y.Y.F., H.K.H.L., L.C., J.F., K.S.M.L., E.Y.M.W. and S.S.A. performed the experiments. S.K.P.L., R.F.F.Y., H.K.H.L., J.F., K.S.M.L., J.F.W.C., R.K.H.K., K.H.C., U.W., K.Y.Y. and P.C.Y.W. analyzed the data. S.K.P.L., R.F.F.Y., H.K.H.L., K.S.M.L., L.C. and P.C.Y.W. wrote the paper.
Conflict of interest
The authors declare that they have no conflict of interest.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Related Research Data
References
- de Groot R. J. et al. Coronaviridae. In: Virus Taxonomy, Classification and Nomenclature of Viruses. Ninth Report of the International Committee on Taxonomy of Viruses, International Union of Microbiological Societies, Virology Division, 806–828 (2011).
- WooPCet al.Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirusJ. Virol.201286 3995 400810.1128/JVI.06540-1122278237
- WooPCet al.Comparative analysis of twelve genomes of three novel group 2c and group 2d coronaviruses reveals unique group and subgroup featuresJ. Virol.2007811574158510.1128/JVI.02182-0617121802
- GorbalenyaAESnijderEJSpaanWJMSevere acute respiratory syndrome coronavirus phylogeny: toward consensusJ. Virol.2004787863786610.1128/JVI.78.15.7863-7866.200415254158
- DrostenCet al.Identification of a novel coronavirus in patients with severe acute respiratory syndromeNew Engl. J. Med.20033481967197610.1056/NEJMoa03074712690091
- FouchierRAMet al.A previously undescribed coronavirus associated with respiratory disease in humansProc. Natl. Acad. Sci.20041016212621610.1073/pnas.040076210115073334
- KsiazekTGet al.A novel coronavirus associated with severe acute respiratory syndromeNew Engl. J. Med.20033481953196610.1056/NEJMoa03078112690092
- PeirisJSMet al.Coronavirus as a possible cause of severe acute respiratory syndromeLancet20033611319132510.1016/S0140-6736(03)13077-212711465
- van der HoekLet al.Identification of a new human coronavirusNat. Med.20041036837310.1038/nm102415034574
- WooPCet al.Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumoniaJ. Virol.20057988489510.1128/JVI.79.2.884-895.2005538593
- ZakiAMvan BoheemenSBestebroerTMOsterhausADFouchierRAIsolation of a novel coronavirus from a man with pneumonia in Saudi ArabiaNew Engl. J. Med.20123671814182010.1056/NEJMoa1211721
- de GrootRJet al.Middle East respiratory syndrome coronavirus (MERS-CoV): announcement of the Coronavirus Study GroupJ. Virol.2013877790779210.1128/JVI.01244-1323678167
- WooPCYet al.Comparative analysis of 22 coronavirus HKU1 genomes reveals a novel genotype and evidence of natural recombination in coronavirus HKU1J. Virol.2006807136714510.1128/JVI.00509-061489027
- KottierSACavanaghDBrittonPExperimentalVirology199521356958010.1006/viro.1995.0029
- LauSKPet al.Isolation and characterization of a novel betacoronavirus subgroup a coronavirus, rabbit coronavirus HKU14, fromJ. Virol.2012865481549610.1128/JVI.06927-113347282
- LauSKPet al.Ecoepidemiology and complete genome comparison of different strains of severe acute respiratory syndrome-related rhinolophus bat coronavirus in china reveal bats as a reservoir for acute, self-limiting infection that allows recombination eventsJ. Virol.2010842808281910.1128/JVI.02219-0920071579
- LauSKPet al.Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombinationJ. Virol.201185113251133710.1128/JVI.05512-113194943
- LauSKPet al.Coexistence of different genotypes in the same bat and serological characterization of rousettus bat coronavirus HKU9 belonging to a novel betacoronavirus subgroupJ. Virol.201084113851139410.1128/JVI.01121-102953156
- LauSKPet al.Discovery of a novel coronavirus, China Rattus Coronavirus HKU24, from Norway rats supports the murine origin of betacoronavirus 1 and has implications for the ancestor of betacoronavirus lineage AJ. Virol.2014893076309210.1128/JVI.02420-144337523
- GuanYIsolation and characterization of viruses related to the SARS coronavirus from animals in southern ChinaScience200330227627810.1126/science.108713912958366
- LauSKet al.Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe batsProc. Natl. Acad. Sci. USA2005102140401404510.1073/pnas.0506735102
- LiWBats are natural Reservoirs of SARS-like coronavirusesScience200531067667910.1126/science.111839116195424
- LauSKet al.Severe acute respiratory syndrome (SARS) coronavirus ORF8 protein is acquired from SARS-related coronavirus from greater horseshoe bats through recombinationJ. Virol.201589105321054710.1128/JVI.01048-154580176
- SongHDet al.Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and humanProc. Natl. Acad. Sci.20051022430243510.1073/pnas.0409608102
- ChineseSMECMolecular evolution of the SARS coronavirus during the course of the SARS epidemic in ChinaScience20043031666166910.1126/science.1092002
- LauSKPet al.Recent transmission of a novel alphacoronavirus, bat coronavirus HKU10, from Leschenault’s Rousettes to Pomona Leaf-Nosed Bats: first evidence of interspecies transmission of coronavirus between bats of different subordersJ. Virol.201286119061191810.1128/JVI.01305-123486284
- WooPCet al.Discovery of a novel bottlenose dolphin coronavirus reveals a distinct species of marine mammal coronavirus in GammacoronavirusJ. Virol.2014881318133110.1128/JVI.02351-133911666
- ZhangJet al.Genomic characterization of equine coronavirusVirology20073699210410.1016/j.virol.2007.06.035
- HasoksuzMet al.Biologic, antigenic, and full-length genomic characterization of a bovine-like coronavirus isolated from a giraffeJ. Virol.2007814981499010.1128/JVI.02361-061900194
- JinLet al.Analysis of the genome sequence of an alpaca coronavirusVirology200736519820310.1016/j.virol.2007.03.03517459444
- ReuskenCBet al.Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: a comparative serological studyLancet Infect. Dis.20131385986610.1016/S1473-3099(13)70164-623933067
- ChanJFWet al.Middle East respiratory syndrome coronavirus: another zoonotic betacoronavirus causing SARS-like diseaseClin. Microbiol. Rev.20152846552210.1128/CMR.00102-1425810418
- WooPCYet al.Molecular diversity of coronaviruses in batsVirology200635118018710.1016/j.virol.2006.02.041
- LauSKet al.Genetic characterization of betacoronavirus Lineage C viruses in bats reveals marked sequence divergence in the spike protein of pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: implications for the origin of the novel Middle East respiratory syndrome coronavirusJ. Virol.2013878638865010.1128/JVI.01055-133719811
- YangLet al.MERS-related betacoronavirus in Vespertilio superans bats, ChinaEmerg. Infect. Dis.2014201260126210.3201/eid2007.1403184073873
- CormanVMet al.Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African batJ. Virol.201488112971130310.1128/JVI.01498-144178802
- Anthony, S. J. et al. Further evidence for bats as the evolutionary source of Middle East respiratory syndrome coronavirus. mBio8 (2017).
- GeldenhuysMet al.A metagenomic viral discovery approach identifies potential zoonotic and novel mammalian viruses in Neoromicia bats within South AfricaPloS One201813e019452710.1371/journal.pone.01945275868816
- Lau, S. K. P. et al. Receptor usage of a novel bat lineage C betacoronavirus reveals evolution of MERS-related coronavirus spike proteins for human DPP4 binding. J. Infect. Dis.218,197–207 (2018).
- CormanVMet al.Characterization of a novel betacoronavirus related to middle East respiratory syndrome coronavirus in European hedgehogsJ. Virol.20148871772410.1128/JVI.01600-133911734
- YangYet al.Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirusProc. Natl. Acad. Sci. USA2014111125161252110.1073/pnas.1405889111
- WangQet al.Bat origins of MERS-CoV supported by bat coronavirus HKU4 usage of human receptor CD26Cell host & Microbe20141632833710.1016/j.chom.2014.08.009
- CaiYet al.CD26/DPP4 cell-surface expression in bat cells correlates with bat cell susceptibility to Middle East respiratory syndrome coronavirus (MERS-CoV) infection and evolution of persistent infectionPloS One20149e11206010.1371/journal.pone.01120604237331
- MunsterVJet al.Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis)Sci. Rep.2016610.1038/srep218784761889
- ChanJFet al.Differential cell line susceptibility to the emerging novel human betacoronavirus 2c EMC/2012: implications for disease pathogenesis and clinical manifestationJ. Infect. Dis.20132071743175210.1093/infdis/jit123
- EckerleIet al.Replicative capacity of MERS coronavirus in livestock cell linesEmerg. Infect. Dis.20142027627910.3201/eid2002.1311823901466
- Vergara-AlertJet al.Livestock susceptibility to infection with Middle EastEmerg. Infect. Dis.20172323224010.3201/eid2302.1612395324816
- BanerjeeAet al.Generation and characterization of eptesicus fuscus (Big brown bat) kidney cell lines immortalized using the myotis polyomavirus large T-antigenJ. Virol. Methods201623716617310.1016/j.jviromet.2016.09.00827639955
- Lau, S. K. P., Wong, A. C. P., Lau, T. C. K. & Woo, P. C. Y. Molecular evolution of MERS coronavirus: dromedaries as a recent intermediate host or long-time animal reservoir? Int. J. Mol. Sci.18 (2017).
- CottenMet al.Transmission and evolution of the Middle East respiratory syndrome coronavirus in Saudi Arabia: a descriptive genomic studyLancet20133821993200210.1016/S0140-6736(13)61887-53898949
- LauSKet al.Polyphyletic origin of MERS coronaviruses and isolation of a novel clade A strain from dromedary camels in the United Arab EmiratesEmerg. Microbes Infect.2016510.1038/emi.2016.1295180373
- ChuDKWet al.MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversityProc. Natl. Acad. Sci. USA20181153144314910.1073/pnas.1718769115
- LuGet al.Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26Nature201350022723110.1038/nature1232823831647
- WangNet al.Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4Cell Res.20132398699310.1038/cr.2013.9223835475
- LiWet al.Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirusNature200342645045410.1038/nature0214514647384
- CormanVMet al.Evidence for an ancestral association of human coronavirus 229E withJ. Virol.201589118581187010.1128/JVI.01755-154645311
- CormanVMet al.Link of a ubiquitous human coronavirus to dromedary camelsProc. Natl. Acad. Sci. USA20161139864986910.1073/pnas.1604472113
- Tao, Y. et al. Surveillance of bat coronaviruses in Kenya identifies relatives of human coronaviruses NL63 and 229E and their recombination history. J. Virol. 91 (2017).
- CrameriGet al.Establishment, immortalisation and characterisation of pteropid bat cell linesPloS One20094e826610.1371/journal.pone.00082662788226
- Lau, S. K. et al. Delayed induction of proinflammatory cytokines and suppression of innate antiviral response by the novel Middle East respiratory syndrome coronavirus: implications for pathogenesis and treatment. J. Gen. Virol. 94, (Pt 12) 2679–90 (2013).