Abstract
Quantification of landscape-based vegetation structural variation and pattern is a significant goal for a variety of ecological, monitoring and biodiversity studies. Vegetation structural metrics, derived from airborne laser scanning (ALS or aerial light detection and ranging—LiDAR) and QuickBird satellite imagery, were used to establish the degree of plot-based vegetation variation at a hillslope scale. Topographic position is an indicator of energy and water availability, and was quantified using DEM-based insolation and topographic wetness, respectively, stratifying areas into hot-warm-cold and wet-moist-dry topographic classes. A range of vegetation metrics—maximum and modal canopy height, crown cover, foliage cover, NDVI and semivariance—were compared among randomly selected plots from each topographic class. NDVI increases with increasing landscape wetness, whereas ALS-derived foliage cover decreases with increasing insolation. Foliage cover is well correlated with crown cover (R 2 =0.65), and since foliage cover is readily calculable for whole-of-landscape application, it will provide valuable and complementary information to NDVI. Between-plot heterogeneity increases with increasing wetness and decreasing insolation, indicating that more sampling is required in these locations to capture the full range of landscape-based variability. Pattern analysis in landscape ecology is one of the fundamental requirements of landscape ecology, and the methods described here offer statistically significant, quantifiable and repeatable means to realise that goal at a fine spatial grain.
Introduction
One key driver of vegetation variation is the underlying landform (Swanson et al. Citation1988; Franklin Citation1995, Citation1998). Vegetation structure is known to vary with landscape position, but is rarely quantified on a systematic basis, because traditional methods are site restricted and so are costly and time consuming to extend, or are prohibited by lack of access. Yet vegetation structure is a key component of vegetation condition assessment (Parkes et al. Citation2003; Gibbons & Freudenberger Citation2006), and structural variation and complexity can be used as a biodiversity surrogate (McElhinny et al. Citation2005). If structure varies with location, site-based assessment is unlikely to encapsulate the structural heterogeneity of a vegetation community. Australian field studies have shown that aspect can affect vegetation structure (Bale et al. Citation1998), in a manner analogous to the often-recognised difference in riparian and upslope vegetation form (Austin et al. Citation1996; Catterall et al. Citation2001). In addition, crown and open-canopy size can vary across the landscape at a number of scales (Hobbs & Cramer Citation2003). Remote sensing offers methods for landscape-based vegetation assessment, since it allows a synoptic overview of the landscape, and information can be determined by a variety of means at different scales so that a range of vegetation characteristics can be captured.
Recent advances in high spatial resolution remote sensing, such as airborne laser scanning (ALS: also known as LiDAR—light detection and ranging), allow assessment of vegetation structure from a site to individual tree scales. The key importance of ALS is the ability to measure metrics normally determined by ground observation, such as foliage cover, canopy cover, and tree height, and correlate them with ground-based observations with a high degree of confidence (Lefsky et al. Citation2002; Hyyppã et al. Citation2008). Because ALS measures the 3D distribution of earth surface features as well as the arrangement of vegetation canopies, it can also be used to model the geomorphic landscape on which vegetation depends. Digital elevation models (DEMs) are gridded representations of the earth's surface, and DEM-based topographic derivatives, such as slope, aspect, drainage and catchment boundaries, can be readily and repeatedly defined using standard GIS procedures (Wilson & Gallant Citation2000). From a vegetation perspective, the key landscape variables are the availability of energy and water (Stephenson Citation1998; Wright et al. Citation2004), which may be modelled by DEM-based insolation and topographic wetness (Burrough & McDonald Citation1998; Urban et al. Citation2000).
To date, the majority of remote-sensing studies involving vegetation pattern and landscape position have focused on moderate spatial resolution imagery with a resolution of 30 m and gridded elevation data from c.1:25k topographic mapping (Burrough & McDonald Citation1998; Franklin Citation1998; Deng et al. Citation2007) principally due to data availability. High-resolution studies, with or without a landscape context, are commonly applied in temperate or boreal regions (Johansen & Phinn Citation2007; Hyyppã et al. Citation2008; Thompson et al. Citation2008), but analysis in tropical (e.g. Clark et al. Citation2004) or sub-tropical areas is less common. Compared to temperate coniferous–deciduous forests, Australian sub-tropical eucalypt forest and woodland have significantly different structural characteristics, particularly in open, asymmetric crown configuration and low foliage density (Specht & Specht Citation1999); also, leaf angles are different from those found in deciduous forests, which have a more horizontal structure. Since both multi-spectral satellite imaging and laser scanning technologies depend on reflectance and transmittance characteristics of foliar structure, techniques developed for cool temperate or boreal zones may not be directly transferable. Similarly, climate-landscape controls on vegetation pattern may differ, for example the higher solar azimuth will tend to reduce the effect of topographic aspect, compared to the greater degree of shadowing experienced in higher-latitude zones.
Semivariance (Goovaerts Citation1997; Kent et al. Citation2006) has found increasing use for pattern analysis in various remote-sensing applications. Semivariance measures spatial autocorrelation as a function of the distance between multiple point pairs and thus the average structural dimension, or range, represented in the data (Cressie Citation1991). Semivariance is often used to detect the scale at which an ecological pattern occurs (Meisel & Turner Citation1998), and has been used in a variety of applications including catena and vegetation pattern (Bedford & Small Citation2008), canopy height prediction (Hudak et al. Citation2002) and landslide analysis (Glenn et al. Citation2006). Semivariance is mathematically related to Moran's I and Geary's C measures of spatial pattern (Dale et al. Citation2002), and is the preferred measure owing to its speed of application and available software (Geostatistical Analyst: ESRI Inc.). A small number of studies have used semivariance for high-resolution vegetation analysis of crown, canopy gap (open canopy) and stand dimension (Levesque & King Citation2003; Johansen et al. Citation2007). At the data resolution of the current study, horizontally autocorrelated variables include open canopy to crown and inter-crown elevation, and spectral variation within and between canopy and open-canopy areas. For discrimination of open-canopy–canopy scale, the elevation or spectral variables must have a greater between-feature rather than within-feature response.
In this study we discriminate a range of fine-scale vegetation metrics using high spatial resolution QuickBird satellite imagery and airborne ALS data in eucalypt woodland-forest in south-eastern Australia, and compare the metrics with DEM-derived insolation and topographic wetness. While not establishing causation, any correlation between forest metrics and topographic variables can be used for inference about growth condition and limitations. For example, there is a well-established linear relationship between growth rate and radiant energy (Bell & Williams Citation1997), so that an inverse relationship implies some other limiting factor. Previous field assessment in the study area suggests that a range of vegetation types are associated with particular landforms (NPWS Citation2003); however, those associations have not been rigorously quantified and tested. The requirement, therefore, is to quantify vegetation structure according to its landscape position, and to determine whether a consistent relationship emerges between the two.
The key questions investigated are: (1) how does fine-scale vegetation structure vary with landscape position, where position refers to solar orientation as well as downslope moisture accumulation; and (2) does ALS data and QuickBird multi-spectral imaging provide spatially distinct, and hence complementary, vegetation metrics at landscape scale?
Methods
Study area
The study area comprises approximately 40 km2 of mixed eucalyptus woodland, associated with tall open forest, gully rainforest, heathland and headwater swamps, located in elevated coastal hinterland, approximately 70 km south-west of Sydney, Australia (see ). The study area forms part of the approximately 1000 km2 Woronora Plateau (NPWS Citation2003) and falls within the Sydney water-supply catchment (SCA Citation2007). The underlying lithology is dominated by relatively impervious quartzose sandstone, producing shallow watertables with subsurface drainage controlled by joints and seepage planes (Hazleton & Tille Citation1990). Landform consists of dissected sandstone plateaux, with an elevated easterly escarpment draining towards the west (Young & Young Citation1988). Evaporation generally exceeds precipitation in the area with similar topography and vegetation to the north, except at the seaward edge of the plateau, where orographic rainfall effects are most pronounced (Keith et al. Citation2006). While a pronounced east to west rainfall gradient is present over the 20 km width of the plateau (NPWS Citation2003), the current study area was chosen to avoid significant rainfall gradient, in order to focus on vegetation variation within local topography. Given the presence of shallow groundwater and evaporative demand, seasonal drought conditions may apply to the study site. Vegetation variation at regional (approximately 10 km) and local, hillslope scale is believed to be associated principally with rainfall and topographic variation (NPWS Citation2003). Local, edaphic controls on plant species distribution in wet heathland (upland swamps) are well established (Keith & Myerscough Citation1993). Moisture-related variations seem to be more significant for heath–woodland transitions in the Blue Mountains, which is in a similar physiographic setting but west of the current study area, where control by shallow soils, related to slope length and inclination, rather than aspect, is dominant (Wilkinson & Humphreys Citation2006). Temporal and spatial variation also occurs in response to fire, with several major fires since 1960 affecting heathland floristic succession (Keith et al. Citation2007), and lower intensity burns occurring on southern slopes after the 2001 wildfire (Chafer et al. Citation2004). The vegetation is relatively undisturbed, with agricultural development historically inhibited by low-nutrient soils typical of the underlying Permian–Triassic lithologies (Hazleton & Tille Citation1990) and rugged topography (see ). Localised disturbance includes powerline corridors, water supply reservoirs and fire trails. Recent disturbance includes temporary track construction for seismic survey access (Illawarra Coal Holdings Pty Ltd Citation2007).
FIGURE 1. Location map.
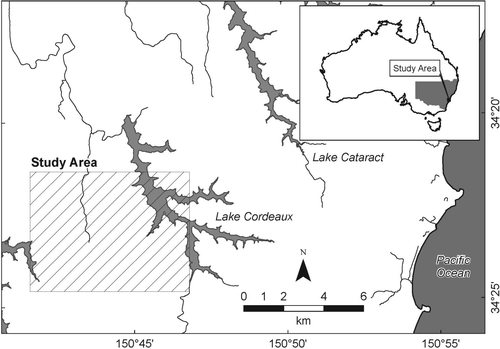
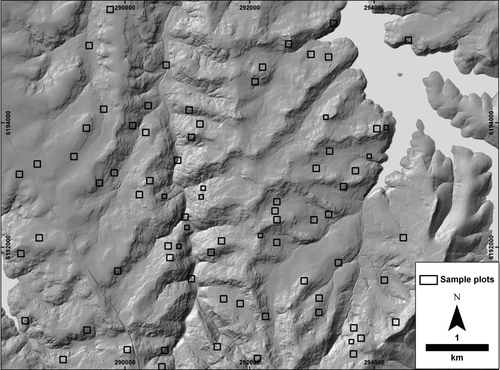
PLATE 1. Topography and sample plots. Hillshade solar angle 45°; azimuth 315°. See Jenkins (Citation2009) for additional field site data.
Data
Field data
The dominant vegetation type in the study area consists of woodland dominated by Eucalyptus sieberi, E. piperita, Corymbia gummifera and E. racemosa/E. haemastoma. This association is widespread in the coastal Sydney Basin, with E. sieberi and E. piperita extending into the montane hinterland (Keith Citation2004). Woodland understorey is typically dense shrub or heath, with high species diversity. Dominant understorey heathland genera include Leptospernum, Banksia and Hakea. Eucalyptus haemastoma is dominant in shallow watertable areas adjacent to wet heathlands, and E. piperita becomes dominant in tall open forest, where its growth form changes from woodland (short trunk with irregular branches) to forest form (long trunk with spreading crown). Detailed floristic inventory is available elsewhere (NPWS Citation2003).
Eight field sites, averaging 4000 m2, and 2 km of line transect were established as part of an ongoing field program. Site data collected included tree location, species, canopy height and closure, and understorey strata equivalent to Floristic Association specification (Hnatiuk et al. Citation2009). Trees were located either by differential GPS (dGPS) or by laser rangefinder from a central location within the site. Linear transects used a stem-to-stem zigzag method (Hnatiuk et al. Citation2009) along 100 m baselines, with dGPS used to record stem and crown edge locations. Crown height class, diameter at breast height and dominant tree species were also recorded. Results of the field study are presented in Jenkins (Citation2009).
Imagery and ALS survey
Orthorectified colour aerial photography was flown in March 2002 and October 2007 (0.2 m and 0.25 m resolution, respectively). Two QuickBird images (DigitalGlobe Citation2008) were acquired on 30 January 2006 and 20 January 2007; however, only one image (2007) was used in the analysis. Image values are supplied by the vendor as top-of-atmosphere radiance. Small-footprint, discrete pulse ALS survey of the study area was conducted in January 2006. The initial ALS point spacing was approximately 1 m and the point cloud was separated into ground and non-ground points by the vendor. Both digital elevation (DEM) and canopy elevation (CEM) models were constructed from ALS ground and non-ground data respectively using natural neighbour interpolation at 1 m resolution. Potential CEM interpolation artefacts—a consequence of data gaps in open-canopy areas—were reduced by incorporating spatially selected ground points that fell closer to non-ground points than the nominal point spacing (1 m) to fill the gaps. A canopy height model (CHM) was derived by subtracting the DEM from the CEM. A 3×3 median filter was applied to the CHM to preserve edges while reducing the effect of inter-canopy variations (Popescu et al. Citation2003), which should enhance canopy–open-canopy discrimination. Vegetation metrics require local height of non-ground points, rather than height above sea level. This was derived by subtraction of DEM height from the discrete non-ground data points. QuickBird imagery orthorectification was performed using a 3D method (Toutin Citation2004) with vendor-supplied rational polynomial coefficient files, ground control registration points from aerial imagery, and the 1 m ALS-derived DEM.
Plot sites refer to those areas chosen for remote-sensing sampling (see ‘Plot selection’ section) and are distinct from field sites, which were ground surveyed. Plot-averaged metrics include ALS-derived foliage cover, crown cover, maximum canopy height, and modal canopy height. Multi-spectral metrics include plot-averaged Normalised Difference Vegetation Index (NDVI). NDVI compares the amount of near-infra red to red light reflected, and is a good indicator of chlorophyll abundance; hence plant vigour (Jensen Citation2005). Canopy NDVI was also determined by extracting NDVI values from ALS-derived crown-cover areas, in order to separate the soil-understorey effect in open-canopy areas. Maximum canopy height was based on the 99th height percentile, to reduce outlier effects (Riano et al. Citation2003). Modal canopy height was determined after inspection of the height frequency histogram, with the value selected from within the canopy portion of the profile. Foliage cover was based on the ratio of non-ground to total count points per plot (Holmgren et al. Citation2003; Riano et al. Citation2003). Crown cover was based on CHM stratification at 10 m, from the field-based height used to differentiate understorey and canopy. Geostatistics were used to establish the autocorrelated range for ALS point elevations or gridded NDVI values extracted to points. The range was then used to infer the predominant size of tree crowns or crown clusters and open-canopy gaps.
Landscape position classification
Landscape position was stratified into classes to provide the independent variables to evaluate vegetation variation over the landscape. Position classification was by insolation and topographic wetness index (TWI), as proxies for vegetative energy and water requirements, respectively. Insolation (Kumar et al. Citation1997) was calculated as incoming direct radiation at monthly intervals, using the ALS DEM (Spatial Analyst: ESRI Inc.). TWI is defined in terms of the natural logarithm of upslope contributing area (α) and slope (β) as ln(a/tanβ) (Sørensen & Seibert Citation2007), so that locations with larger upslope contributing areas receive greater moisture, with moisture residency controlled by slope. The ALS-based DEM was used for landform classification, and was resampled to 10 m spatial resolution to reduce microtopographic artefacts, such as rough ground due to blocky sandstone substrate and reflections from tree bases and trash accumulation. Computation of TWI (Spatial Analyst: ESRI Inc.) at fine scale also introduces artefacts of alternating wet and dry zones, particularly in valley bottom areas, where a more consistent moisture profile would be expected (Burrough et al. Citation2000). Using the 10 m DEM reduced the topographic and TWI artefacts to an acceptable level, producing landform classes of suitable width for subsequent within-class sampling plots.
The insolation and TWI surfaces were stratified into three classes of equal area (cool-warm-hot and wet-moist-dry). The insolation classification thresholds were 420 938–1 406 220 (cool), 1 406 220–1 466 355 (warm) and 1 466 355–1 600 501 (hot). The TWI classification thresholds were 0.519–2.592 (dry), 2.592–3.672 (moist) and 3.672—11.532 (wet). A low-pass filter and boundary clean commands were used to aggregate the TWI classes (Spatial Analyst: ESRI Inc.). Classification effectively identified ridge, midslope and valley areas (dry–wet TWI) as well different aspect (insolation), especially along linear areas with even slope. More complex, interspersed landform was seen on convex shoulders and nose slopes, especially for east or west aspects, where insolation transition is highest. Similar complexity was apparent in convex headslopes, where wetness is expected to change over short distances. The interspersed distance variation meant that sample plots (below) were more easily located on plateaux and broader valleys (typical landform class widths c.100–300 m), but less easily located in transitional concave–convex areas (typical landform class widths c.10–50 m).
Plot selection
Remote-sensing sample plots were located randomly within each insolation and TWI class. Approximately 100 potential plots were initially identified, and plots near roads and other anthropogenic disturbances excluded, based on visual image interpretation. A total of 65 100×100 m (1 ha) plots, with an additional 11 71×71 m (0.5 ha) plots, to cover narrow landform zones, were established, yielding approximately 30 plots per landscape position class (see ). Owing to plot randomisation and landscape stratification effects, remote-sensing plots did not coincide with established field sites.
Vegetation pattern analysis
Semivariance of NDVI and CHM data was used in order to assess canopy spatial pattern. At the data scale of this study, the semivariance range is expected to relate to canopy–open-canopy spacing, as a result of systematic height variation from canopy to open-canopy areas in ALS data or NDVI values for spectral data. An increase in semivariance range will be associated with an increase in crown and open-canopy dimension. Gridded point data were used, with lag width based on the ALS and multi-spectral pixel size (1 m and 2.4 m), with a 12 lags search distance (12 m and 28.8 m, respectively). The widely used spherical model was used for semivariogram modelling (Geostatistical Analyst: ESRI Inc.), and since frequency distribution of plot data for height and NDVI was approximately normal, no data transformation was undertaken.
Statistical methods
To establish how vegetation varies with landscape position (Question 1), one-way Analysis of Variance (ANOVA) was used to extract significant difference of means for the vegetation metric-landscape position classes, using a Shapiro–Wilk's W statistic to determine whether the metrics were normally distributed (Shapiro et al. Citation1968). Statistical significance was assessed at α<0.05. For interpretation, data were classed as normal (p>0.05), sub-optimal (p ≥ 0.01), or non-normal (p<0.01). Post hoc assessment of ANOVA with Fischer's Least Significant Difference (LSD) and Scheffé tests was applied to establish which metric-landform classes were significantly different. Both tests were accepted for normally distributed data, but only the more conservative Scheffé test was accepted for sub-optimal data. Ad hoc assessment was applied for non-normal data, where apparently significant post hoc separation was encountered.
Significance tests also established whether a range of metrics from both multi-spectral and ALS sources were required to describe landscape-based variation (Question 2). To gain an appreciation of metric redundancy, NDVI was used as the predictor variable for regression with foliage cover and for spatial pattern assessment with foliage height values (1 m lag semivariance), with Pearson's R 2 values used for assessment. NDVI values were used as predictor variables as the values were found to be normally distributed, compared to the sub-optimally distributed foliage cover and foliage height values. Regression was also calculated for foliage cover and crown cover, as these metrics appeared to be consistently related. Qualitative assessment of variance range was used to evaluate metric consistency between different landscape position classes (Question 1).
Results
Vegetation-landscape comparison
A number of metrics showed significant difference for insolation and TWI classes (see ). In particular, mean NDVI (averaged over the entire plot) increased from dry to wet plots. A similar but weaker trend was observed for canopy-only NDVI, indicating that NDVI variation also occurs in the understorey of open-canopy areas. Foliage cover shows significant increase from hot to cool plots, as does canopy semivariance range (1 m lags). Crown cover, maximum height and canopy mode height increase both from dry to wet and hot to cool plots, although the relationship is not as strong and crown cover has a very high level of variance. Non-normal values for crown cover, maximum height and broader-scale semivariance (12 lags at 2.4 m) are unsurprising, given the heterogeneous canopy–open-canopy pattern seen in CHM mapping and imagery. The overall trend is for taller, greener vegetation in cool-wet plots, and that different metrics capture different, significant vegetative characteristics. For example, the relationship between foliage cover (as proxy for above-ground biomass) and NDVI (photosynthetically active vegetation) is decoupled, with foliage cover showing an inverse-energy response as opposed to NDVI increasing with water. A consistent relationship is that a higher degree of variability is evident for cool and wet plots.
TABLE 1. Plot means, standard error and ANOVA significance α<0.05
Correlation—regression
For redundancy assessment of multi-spectral vs ALS data, a comparison between spectral (NDVI) and structural (ALS) properties on a plot-by-plot basis was made (see ). This showed that a generally weak to moderate positive relationship exists between NDVI and foliage cover for dry and moist plots, as well as for cool and hot plots. The relationship did not extend, however, to warm or wet plots and the overall relationships do not hold if the plots are pooled. NDVI values have been shown to saturate at high Leaf Area Index (LAI) levels (Turner et al. Citation1999). Since LAI is a direct measure of foliage density (Watson Citation1947) this may explain the lack of correlation between NDVI and foliage cover. However, a study of mixed moist Eucalyptus forest in the NSW south coast showed a linear relationship between LAI and NDVI (Coops et al. Citation1997), and the LAI for the current study is expected to be lower, so saturation effects are not expected to be of concern. Other comparisons (NDVI vs crown cover and maximum height) showed no consistent relationship. In contrast, a strong relationship for pooled plots was found between foliage cover and crown cover (R 2=0.65; standard error=16 per cent cover; p<0.001), indicating a reasonable level of redundancy between these two ALS-derived metrics.
TABLE 2. Between-plot correlation of QuickBird NDVI vs ALS-derived metrics. The lack of correlation overall shows the lack of redundancy between different metrical and sensor types
Spatial pattern—geostatistics
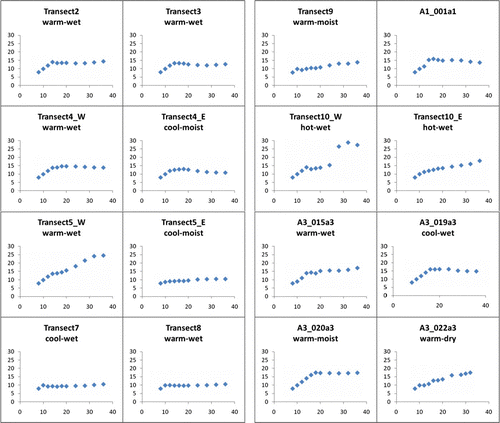
Semivariance ranges were taken over a series of lag widths for selected field sites (see ). Ranges for one metre ALS data semivariance lags (12 m search distance) typically do not plateau, whereas maximum ranges are typically met at 2.4 m lags (28.8 m search distance), indicating that the latter distance gives a good indication of dominant feature size within the plots. The canopy–open-canopy dimension range for TWI class shows a significant increase from dry to wet plots for ALS height values, whereas a similar but statistically insignificant trend is apparent for NDVI values (see ). Hot plots have smaller ranges than warm or cool plots. In other words, crown and inter-canopy gap dimensions are smaller in sites with high insolation values. Greater heterogeneity (higher variance) is apparent for 2.4 m lags, with higher overall canopy–open-canopy dimensions (c.10–16 m). While the 2.4 m lag data are non-normally distributed, the consistent pattern of dry plot smaller dimension and cool plot larger dimension agrees well with the significant results observed in the 1 m lag data.
FIGURE 2. Semivariance range plots for ALS height data. Semivariance range (vertical axis) for a series of lag widths (horizontal axis) for selected field sites. The range typically plateaus at a certain lag width, which indicates the size of dominant features within the field sites. Distances in metres. Site A1_001 was outside the area of landscape stratification. See Jenkins (Citation2009) for additional site details.
If inspection of scatter plots for NDVI vs CHM semivariance is made (not shown), a weak scatter of data beyond 15 m towards the maximum search distance (28.8 m) is demonstrated, which indicates that the range of canopy–open-canopy dimension has been adequately explored. A weak but consistent plot-by-plot correlation for cool and wet plots exists for ALS height vs NDVI at 1 m lag semivariance ranges (see ). These relationships diminish considerably at 2.4 m lag distances.
TABLE 3. Correlation of NDVI vs CHM semivariance ranges. The lack of correlation overall shows the lack of redundancy between different metrical and sensor types
The general trends from the results are shown in . Critically, note that NDVI and horizontal dimension (semivariance range) respond to landscape moisture variation, whereas foliage and crown cover respond to energy, demonstrating both the separability of landform types, as well as the complementary nature of ALS and multi-spectral analyses.
PLATE 2. Vegetation responses to energy and water availability (scale bar 50 m). Representative illustrations from sample plots (a) NDVI increases with increasing water; (b) foliage cover decreases with increasing energy (field photographs); (c) canopy-gap dimension increases with increasing water (from CHM 10 m stratification).

Discussion
Quantification of vegetation structure and its relationship to landscape position represents a significant advance in vegetation ecology, particularly for Australian, eucalypt-dominated environments. While other Australian studies have examined low-rainfall woodland areas (Lee & Lucas Citation2007), to our knowledge this is the first such study in higher-rainfall woodlands with scrubby understorey, which are a distinct regional vegetation type in SE Australia (Keith Citation2004). The current study shows that remote-sensing vegetation metrics show significant differences for different landform classes, and that no particular metric encapsulates all variation.
Question 1: how does fine-scale vegetation structure vary with landscape position?
The partitioning of vegetation structure via both insolation and TWI confirms the necessity to separate these components for landscape vegetation studies, where different ‘dry’ effects can be related to high evapotranspiration demand on sunlit slopes vs areas with low overall water availability (Stephenson Citation1998). Typically, vegetation structural dimensions—NDVI and foliage cover, and, to a more uncertain extent, tree height and crown cover—increase in cool or wet locations. Subtle trends in horizontal pattern—the average canopy–open-canopy dimension—are also evident: warmer and drier plots typically have smaller crowns and open-canopy gaps compared to cool and wet plots, evident from ALS-based semivariance range. The less distinct NDVI pattern at 1 m lags is not surprising, both because QuickBird multi-spectral pixel size (2.4 m) may not sufficiently resolve feature dimensions of c.8–10 m and because relatively high NDVI levels in the shrubby understorey confounds the metric. The semivariance range variation is less than expected, given the visible trends in canopy–open-canopy dimension apparent on image and CHM data. This requires further investigation, since presumably either the metric or its application scale was not able to capture the full pattern variation. Metrics for hot and dry landform classes show the least variance, in general, indicating that vegetation structure is more consistent and predictable in these locations. Cool and wet plots, by comparison, have higher levels of variance, suggesting greater structural heterogeneity. The presence of upland swamps and fringing woodland in wet areas, which have substantially reduced canopy cover, contributes to heterogeneity in these landscape areas. These results are important for landscape-based assessment, such as mapping or field sampling, as it demonstrates that more attention must be paid to cool-wet locations in order to capture the full range of vegetation characteristics present within a study area. While an ALS-based study of SE Australian forests also found less predictability for forests with greater height and canopy closure (Lee & Lucas Citation2007), good results have been attained in northern NSW tall open forests (Goodwin et al. Citation2007), suggesting further work is necessary to clarify the applicability of ALS and high-resolution imagery metrics for dense Australian forests.
Question 2: can ALS and QuickBird data provide complementary assessment for a range of vegetation metrics?
Foliage cover (ALS) and NDVI (QuickBird) are shown here to be both significant and separate measures of vegetation structure and their variation within the landscape, thus demonstrating the data source complementarities. Foliage cover is closely related to crown cover, and the importance of crown-cover mapping for forest ecology is well established (Stone & Haywood Citation2006; Goodwin et al. Citation2007). Foliage cover is also less variable than crown cover for particular landscape classes, so should allow better vegetation discrimination. Also, since foliage cover is a simple ratio metric it is more readily calculated for whole-of-landscape mapping than spatially disjunctive crown cover, although crown-cover mapping is still important since it allows separation of plot-level crown and understorey NDVI, which further allows evaluation of soil-shrub abundance in open-canopy areas. NDVI is being used for landscape-scale fire-intensity prediction (Chafer et al. Citation2004; Hammill & Bradstock Citation2006), so foliage cover may provide complementary information for fine-scale fire risk assessment.
Quantitative vegetation metrics derived from ALS and multi-spectral data can complement and extend field-based structural assessment, and can distinguish vegetation pattern variation at landscape scale, even where relatively subtle structural variation occurs. The potential applications for these methods include landform-based fire-fuel modelling, interpretation of ecological variation and gradients within a landscape-based context, enhanced vegetation mapping, vegetation monitoring using quantitative assessment, and incorporation of structural assessment for biodiversity studies.
Acknowledgements
Funding for this project was through Australian Coal Association Research Project ACARP 15013 and a University of New England Keith and Dorothy Mackay Travel Scholarship to Ross Jenkins. We thank BHP Billiton – Illawarra Coal for access to ALS data and orthophotography. Logistical support in the field by BHPBIC and Biosis Pty Ltd is gratefully acknowledged. Ross Jenkins thanks Paul Frazier for research supervision, and we thank Sarah Gergel for preliminary review and two anonymous journal reviewers whose comments significantly contributed to the quality and clarity of the final version.
Related Research Data
REFERENCES
- Austin , M.P. , Pausas , J.G. and Nicholls , A.O. 1996 . ‘Patterns of tree species richness in relation to environment in southeastern New South Wales, Australia’ . Australian Journal of Ecology , 21 : 154 – 64 .
- Bale , C.L. , Williams , J.B. and Charley , J.L. 1998 . ‘The impact of aspect on forest structure and floristics in some Eastern Australian sites’ . Forest Ecology and Management , 110 : 363 – 77 .
- Bedford , D.R. and Small , E.E. 2008 . ‘Spatial patterns of ecohydrologic properties on a hillslope-alluvial fan transect, central New Mexico’ . Catena , 73 : 34 – 48 .
- Bell , D.T. & Williams , J.E. 1997 ‘ Eucalypt ecophysiology ’, in Williams , J.E. & Woinarski , J.C.Z. Eucalypt ecology: individuals to ecosystems , Cambridge University Press , Cambridge .
- Burrough , P.A. and McDonald , R.A. 1998 . Principles of geographical information systems , Oxford University Press, New York .
- Burrough , P.A. , van Gaans , P.F.M. and MacMillan , R.A. 2000 . ‘High-resolution landform classification using fuzzy k-means’ . Fuzzy Sets and Systems , 113 : 37 – 52 .
- Catterall , C.P. , Pipe , S.D. , Bunn , S.E. and Michael , A.J. 2001 . ‘Flora and fauna assemblages vary with local topography in a subtropical eucalypt forest’ . Australian Journal of Ecology , 26 : 56 – 69 .
- Chafer , C. , Noonan , M. and Macnaught , E. 2004 . ‘The post-fire measurement of fire severity and intensity in the Christmas 2001 Sydney wildfires’ . International Journal of Wildland Fire , 13 : 227 – 40 .
- Clark , M.L. , Clark , D.B. and Roberts , D.A. 2004 . ‘Small-footprint lidar estimation of sub-canopy elevation and tree height in a tropical rain forest landscape’ . Remote Sensing of Environment , 91 : 68 – 89 .
- Coops , N. , Delahaye , A. and Pook , E. 1997 . ‘Estimation of eucalypt forest Leaf Area Index on the south coast of New South Wales using Landsat MSS data’ . Australian Journal of Botany , 45 : 757 – 69 .
- Cressie , N. 1991 . Statistics for spatial data , John Wiley, New York .
- Dale , M.R.T. , Dixon , P. , Fortin , M.-J. , Legendre , P. , Myers , D.E. and Rosenberg , M.S. 2002 . ‘Conceptual and mathematical relationships among methods for spatial analysis’ . Ecography , 25 : 558 – 77 .
- Deng , Y. , Chen , X. , Chuvieco , E. , Warner , T. and Wilson , J.P. 2007 . ‘Multi-scale linkages between topographic attributes and vegetation indices in a mountainous landscape’ . Remote Sensing of Environment , 111 : 122 – 34 .
- DigitalGlobe 2008 QuickBird imagery products product guide revision 5 , DigitalGlobe Inc., Longmont, CO .
- Franklin , J. 1995 . ‘Predictive vegetation mapping: geographic modelling of biospatial patterns in relation to environmental gradients’ . Progress in Physical Geography , 19 : 474 – 99 .
- Franklin , J. 1998 . ‘Predicting the distribution of shrub species in southern California from climate and terrain-derived variables’ . Journal of Vegetation Science , 9 : 733 – 48 .
- Gibbons , P. and Freudenberger , D. 2006 . ‘An overview of methods used to assess vegetation condition at the scale of the site’ . Ecological Management & Restoration , 7 : S10 – S17 .
- Glenn , N.F. , Streutker , D.R. , Chadwick , D.J. , Thackray , G.D. and Dorsch , S.J. 2006 . ‘Analysis of LiDAR-derived topographic information for characterizing and differentiating landslide morphology and activity’ . Geomorphology , 73 : 131 – 48 .
- Goodwin , N.R. , Coops , N.C. and Culvenor , D.S. 2007 . ‘Development of a simulation model to predict LiDAR interception in forested environments’ . Remote Sensing of Environment , 111 : 481 – 92 .
- Goovaerts , P. 1997 . Geostatistics for natural resources evaluation , Oxford University Press, New York .
- Hammill , K.A. and Bradstock , R.A. 2006 . ‘Remote sensing of fire severity in the Blue Mountains: influence of vegetation type and inferring fire intensity’ . International Journal of Wildland Fire , 15 : 213 – 26 .
- Hazleton , P.A. and Tille , P.J. 1990 . Soil landscapes of the Wollongong-Port Hacking 1:100k sheet , Soil Conservation Service of NSW, Sydney .
- Hnatiuk , R.J. , Thackway , R. & Walker , J. 2009 ‘ Vegetation ’, in National Committee on Soil and Terrain Australian soil and land survey handbook , CSIRO Publishing , Melbourne .
- Hobbs , R.J. and Cramer , V.A. 2003 . ‘Natural ecosystems: pattern and process in relation to local and landscape diversity in southwestern Australian woodlands’ . Plant and Soil , 257 : 371 – 8 .
- Holmgren , J. , Nilsson , M. and Olsson , H. 2003 . ‘Simulating the effects of lidar scanning angle for estimation of mean tree height and canopy closure’ . Canadian Journal of Remote Sensing , 29 : 623 – 32 .
- Hudak , A.T. , Lefsky , M.A. , Cohen , W.B. and Berterretche , M. 2002 . ‘Integration of lidar and Landsat ETM+ data for estimating and mapping forest canopy height’ . Remote Sensing of Environment , 82 : 397 – 416 .
- Hyyppã , J. , Hyyppã , H. , Leckie , D. , Gougeon , F. , Yu , X. and Maltamo , M. 2008 . ‘Review of methods of small-footprint airborne laser scanning for extracting forest inventory data in boreal forests’ . International Journal of Remote Sensing , 29 : 1339 – 66 .
- Illawarra Coal Holdings Pty Ltd 2007 Dendrobium mine area 3A SMP , BHP Billiton Group , Wollongong .
- Jenkins , R.B. 2009 High resolution remote sensing for native vegetation assessment and monitoring: an impact assessment approach , University of New England .
- Jensen , J.R. 2005 . Introductory digital image processing: a remote sensing perspective , Prentice Hall, Upper Saddle River, NJ .
- Johansen , K. , Coops , N.C. , Gergel , S.E. and Stange , Y. 2007 . ‘Application of high spatial resolution satellite imagery for riparian and forest ecosystem classification’ . Remote Sensing of Environment , 110 : 29 – 44 .
- Johansen , K. and Phinn , S. 2007 . ‘Linking riparian vegetation spatial structure in Australian tropical savannas to ecosystem health indicators: semi-variogram analysis of high spatial resolution satellite imagery’ . Canadian Journal of Remote Sensing , 32 : 228 – 43 .
- Keith , D.A. 2004 . Ocean shores to desert dunes: the native vegetation of New South Wales and the ACT , Department of Environment and Conservation, Hurstville .
- Keith , D.A. , Holman , L. , Rodoreda , S. , Lemmon , J. and Bedward , M. 2007 . ‘Plant functional types can predict decade-scale changes in fire-prone vegetation’ . Journal of Ecology , 95 : 1324 – 37 .
- Keith , D.A. and Myerscough , P.J. 1993 . ‘Floristics and soil relations of upland swamp vegetation near Sydney’ . Australian Journal of Ecology , 18 : 325 – 44 .
- Keith , D. , Rodoreda , S. , Holman , L. and Lemmon , J. 2006 . Monitoring change in upland swamps in Sydney's water catchments: the roles of fire and rain , Department of Environment and Conservation, Sydney .
- Kent , M. , Moyeed , R.A. , Reid , C.L. , Pakeman , R. and Weaver , R. 2006 . ‘Geostatistics, spatial rate of change analysis and boundary detection in plant ecology and biogeography’ . Progress in Physical Geography , 30 : 201 – 31 .
- Kumar , L. , Skidmore , A.K. and Knowles , E. 1997 . ‘Modelling topographic variation in solar radiation in a GIS environment’ . International Journal of Geographical Information Science , 11 : 475 – 97 .
- Lee , A.C. and Lucas , R.M. 2007 . ‘A LiDAR-derived canopy density model for tree stem and crown mapping in Australian forests’ . Remote Sensing of Environment , 111 : 493 – 518 .
- Lefsky , M.A. , Cohen , W.B. , Parker , G.G. and Harding , D.J. 2002 . ‘Lidar remote sensing for ecosystem studies’ . BioScience , 52 : 19 – 30 .
- Levesque , J. and King , D.J. 2003 . ‘Spatial analysis of radiometric fractions from high-resolution multispectral imagery for modelling individual tree crown and forest canopy structure and health’ . Remote Sensing of Environment , 84 : 589 – 602 .
- McElhinny , C. , Gibbons , P. , Brack , C. and Bauhus , J. 2005 . ‘Forest and woodland stand structural complexity: its definition and measurement’ . Forest Ecology and Management , 218 : 1 – 24 .
- Meisel , J.E. and Turner , M.G. 1998 . ‘Scale detection in real and artificial landscapes using semivariance analysis’ . Landscape Ecology , 13 : 347 – 62 .
- NPWS 2003 Native vegetation of the Woronora, O'Hares and Metropolitan catchments , Conservation Assessment and Data Unit , NSW National Parks and Wildlife Service , Hurstville .
- Parkes , D. , Newell , G. and Cheal , D. 2003 . ‘Assessing the quality of native vegetation: the “habitat hectares” approach’ . Ecological Management & Restoration , 4 : S29 – S38 .
- Popescu , S.C. , Wynne , R.H. and Nelson , R.F. 2003 . ‘Measuring individual tree crown diameter with lidar and assessing its influence on estimating forest volume and biomass’ . Canadian Journal of Remote Sensing , 29 : 564 – 77 .
- Riano , D. , Meier , E. , Allgower , B. , Chuvieco , E. and Ustin , S.L. 2003 . ‘Modeling airborne laser scanning data for the spatial generation of critical forest parameters in fire behavior modeling’ . Remote Sensing of Environment , 86 : 177 – 86 .
- SCA 2007 Special Areas Strategic Plan of Management , available from: http://www.sca.nsw.gov.au/the-catchments/special-areas ( accessed 11 June 2009 ).
- Shapiro , S.S. , Wilk , M.B. and Chen , H.J. 1968 . ‘A comparative study of various tests for normality’ . Journal of the American Statistical Association , 63 : 1343 – 72 .
- Sørensen , R. and Seibert , J. 2007 . ‘Effects of DEM resolution on the calculation of topographical indices: TWI and its components’ . Journal of Hydrology , 347 : 79 – 89 .
- Specht , R.L. and Specht , A. 1999 . Australian plant communities: dynamics of structure, growth and biodiversity , Oxford University Press, South Melbourne .
- Stephenson , N. 1998 . ‘Actual evapotranspiration and deficit: biologically meaningful correlates of vegetation distribution across spatial scales’ . Journal of Biogeography , 25 : 855 – 70 .
- Stone , C. and Haywood , A. 2006 . ‘Assessing canopy health of native eucalypt forests’ . Ecological Management & Restoration , 7 : S24 – S30 .
- Swanson , F.J. , Kratz , T.K. , Caine , N. and Woodmansee , R.G. 1988 . ‘Landform effects on ecosystem patterns and processes’ . BioScience , 38 : 92 – 8 .
- Thompson , S.D. , Gergel , S.E. and Coops , N.C. 2008 . ‘Classification of late seral coastal temperate rainforests with high spatial resolution QuickBird imagery’ . Canadian Journal of Remote Sensing , 34 : S460 – S470 .
- Toutin , T. 2004 . ‘Review article: geometric processing of remote sensing images: models, algorithms and methods’ . International Journal of Remote Sensing , 25 : 1893 – 1924 .
- Turner , D.P. , Cohen , W.B. , Kennedy , R.E. , Fassnacht , K.S. and Briggs , J.M. 1999 . ‘Relationships between Leaf Area Index and Landsat TM Spectral Vegetation Indices across three temperate zone sites’ . Remote Sensing of Environment , 70 : 52 – 68 .
- Urban , D.L. , Miller , C. , Halpin , P.N. and Stephenson , N.L. 2000 . ‘Forest gradient response in Sierran landscapes: the physical template’ . Landscape Ecology , 15 : 603 – 20 .
- Watson , D.J. 1947 . ‘Comparative physiological studies on the growth of field crops. 1. Variation of net assimilation rate and leaf area between species and varieties, and within and between years’ . Annals of Botany , 11 : 41 – 76 .
- Wilkinson , M.T. and Humphreys , G.S. 2006 . ‘Slope aspect, slope length and slope inclination controls of shallow soils vegetated by sclerophyllous heath—links to long-term landscape evolution’ . Geomorphology , 76 : 347 – 62 .
- Wilson , J.P. & Gallant , J.C. 2000 Terrain analysis: principles and applications , John Wiley , New York
- Wright , I.J. , Reich , P.B. , Westoby , M. , Ackerly , D.D. , Baruch , Z. , Bongers , F. , Cavender-Bares , J. , Chapin , T. , Cornelissen , J.H.C. , Diemer , M. , Flexas , J. , Garnier , E. , Groom , P.K. , Gulias , J. , Hikosaka , K. , Lamont , B.B. , Lee , T. , Lee , W. , Lusk , C. , Midgley , J.J. , Navas , M.-L. , Niinemets , U. , Oleksyn , J. , Osada , N. , Poorter , H. , Poot , P. , Prior , L. , Pyankov , V.I. , Roumet , C. , Thomas , S.C. , Tjoelker , M.G. , Veneklaas , E.J. and Villar , R. 2004 . ‘The worldwide leaf economics spectrum’ . Nature , 428 : 821 – 7 .
- Young , A.R.M. and Young , R.M. 1988 . ‘Altogether barren, peculiarly romantic’: the sandstone lands around Sydney’ . Australian Geographer , 19 : 9 – 25 .