Abstract
The sources of covariation among cognitive measures of Inspection Time, Choice Reaction Time, Delayed Response Speed and Accuracy, and IQ were examined in a classical twin design that included 245 monozygotic (MZ) and 298 dizygotic (DZ) twin pairs. Results indicated that a factor model comprising additive genetic and unique environmental effects was the most parsimonious. In this model, a general genetic cognitive factor emerged with factor loadings ranging from 0.28 to 0.64. Three other genetic factors explained the remaining genetic covariation between various speed and Delayed Response measures with IQ. However, a large proportion of the genetic variation in verbal (54%) and performance (25%) IQ was unrelated to these lower order cognitive measures. The independent genetic IQ variation may reflect information processes not captured by the elementary cognitive tasks, Inspection Time and Choice Reaction Time, nor our working memory task, Delayed Response. Unique environmental effects were mostly nonoverlapping, and partly represented test measurement error.
Modern theories of human cognition tend to view intelligence as a collection of varied abilities (including low, intermediate and high levels of cognitive processing), which can be organised hierarchically (Carroll, Citation1993; Kaufman & Lichtenberger, Citation1999; Roberts & Stankov, Citation1999). In this paper we report diverse measures of cognitive ability (information processing speed, working memory, and IQ) with the aim of understanding the genetic and environmental relationships among these correlated abilities. More specifically, we address whether the measures share the same underlying genetic factor, or whether they relate to each other through a number of different genetic factors. Findings on the significant phenotypic interrelationship among processing speed, working memory and IQ (Miller & Vernon, Citation1996; Salthouse, Citation1996) suggest that a single genetic factor will be the most predominant.
Although genetic studies (Alarcon, Plomin, Fulker, Corley, & DeFries, Citation1998; Petrill et al., Citation1998; Thompson, Detterman, & Plomin, Citation1991) have shown that there are significant genetic correlations between perceptual speed (psychometrically derived), memory and verbal/spatial abilities (factors derived from Cognitive Abilities Test), there have been few studies that directly assess the genetic relationship among processing speed, working memory and IQ. In our previous study of the genetic relations among Choice Reaction Time (CRT; 2-, 4-, and 8-choice), Delayed Response accuracy and Full Scale IQ, two genetic factors were found to mediate the covariance among these measures (Luciano et al., Citation2001a). In the present paper we have added Inspection Time and Delayed Response speed measures to the previously reported battery to establish whether further separation of genetic group factors is possible with a broadened range of cognitive measures.
Information processing speed as measured by elementary cognitive tasks is a conceptually distinct process from perceptual speed measured by psychometric tests. Processing speed and perceptual speed indices are correlated at only 0.16 (Kyllonen, Citation1993). Elementary cognitive task correlates of IQ include such measures as choice reaction time, speed of scanning in short term memory, visual and auditory discrimination ability, and speed of long-term memory retrieval. In general, the RT measures demonstrate correlations of around – 0.30 with IQ, while measures without an RT component, such as perceptual discrimination speed, are higher (e.g., – 0.50) (Deary & Stough, Citation1996; Jensen, Citation1993). In this study we sample two established processing speed measures, Inspection Time and CRT.
Multivariate studies of IQ or specific cognitive abilities have indicated that memory subtests (or factors) are influenced by a genetic general (g) factor, but even more so by a specific genetic factor (Cardon, Fulker, DeFries, & Plomin, Citation1992; Finkel, Pedersen, McGue, & McClearn, Citation1995; Luo, Petrill, & Thompson, Citation1994). There is a further report of a genetic relationship between processing speed and various short-term memory tests, including word recall, immediate and delayed text recall and figure memory (Finkel & McGue, Citation1993). These studies have either used short term memory tasks (which tap working memory storage) or a conglomerate measure of different memory types (e.g., short- and long-term) rather than a specific test of working memory.
To measure working memory, we use a visuospatial delayed response (DR) task, which involves both short-term storage and executive function. This task requires the participant to retain information over the course of a short time delay during which the response is withheld. It is more complex than the processing speed tasks because of the requirement of inhibition, timing the motor response as well as remembering the target position while ignoring distractors. Areas of prefrontal cortex have been implicated in executive function and are activated during task performance, indicating that the DR task does tap some process in the working memory system (Geffen et al., Citation1997; Goldman-Rakic, Citation1992). While accuracy on the DR task serves as an index of working memory, the DR speed measures also have relevance to our study. Delayed response speed is measured by a response initiation time and a movement time. Initiation time refers to the speed with which a participant reacts to a cue and may be akin to simple RT; another processing speed measure. Movement time is the speed with which a person completes a response following response initiation, and it may be associated with CRT because our CRT task uses a keyboard paradigm resulting in a combined decision and movement time response.
Using a classical twin design this study aimed to elucidate the proportion of variance contributed by genes and environment to the covariation among measures of Inspection Time, CRT (2-, 4-, and 8-choice conditions), DR speed and accuracy responses and IQ (verbal, performance). In doing so we seek to establish the genetic and environmental factor structure that best explains the covariation among these diverse cognitive abilities.
Method
Participants
This is an ongoing study of cognition in 16-year-old twins and their nontwin siblings (Wright et al., Citation2001). Here we analyse data from the first 543 twin pairs who participated (129 monozygotic [MZ] female, 116 MZ male, 75 dizygotic [DZ] female, 69 DZ male, 154 opposite sex). Zygosity was determined by ABO, MN and Rh blood groups and by nine independent polymorphic DNA markers. Twin pairs did not participate in the study if either one had a history of significant head injury, neurological or psychiatric illness, substance dependence or if they were currently taking long-term medications with central nervous system effects; approximately 6% of twin pairs were excluded from the study on this basis. Participants had normal or corrected-to-normal vision (better than 6/12 Snellen equivalent). The twins were mostly in their penultimate year of secondary school and aged between 15 and 18 years (16.22 years; SD = 0.32). To increase motivation they received a monetary reward based on their performance on the DR task. Written informed consent was obtained from the participant, as well as their parent/guardian, prior to testing.
Experimental protocol
The Inspection Time (IT), CRT task and IQ test were part of a psychometric battery, which also included two reading tests. The DR task was administered in a parallel testing session and involved the recording of behavioural responses and event-related potentials. Each session approximated 1.5 hr in length. One twin completed the psychometric session while the other completed the working memory session. The order of session testing was counterbalanced between twin pairs based on the birth order of the twins. A full description of the protocol is given in Wright et al. (Citation2001).
Tests
Multidimensional Aptitude Battery.
A shortened version of the MAB (Jackson, Citation1984, Citation1998) was used, which included three verbal subtests, (Information, Arithmetic, Vocabulary) and two Performance subtests (Spatial and Object Assembly). Each subtest had a multiple-choice format and was timed at 7 min. Participants were not penalised for guessing and were encouraged to answer every item within the time period. Administration and scoring were computerised. The IQ scores derived from computerised versus paper-and-pencil administration of the verbal subtests have been shown to be equivalent by Harrell et al. (Citation1987) and by MacLennan et al. (Citation1988). Test – retest correlations over a 3-month interval were calculated from a sample of 49 twin pairs and estimated at 0.87 for performance IQ and 0.89 for verbal IQ (Luciano et al., Citation2001b).
Inspection Time.
Inspection time was tested by a line discrimination task presented as a pseudocomputer game of choosing the longer of two worms to go fishing, as previously described (Luciano et al., Citation2001b). The two lines of comparison were described as worms that would quickly burrow into the ground (i.e., appearance of masking stimulus). The participant's task was to identify the longer worm in an effort to catch the most fish by pressing the corresponding left or right arrow key on the keyboard. Feedback in the form of a fish appeared at the lower left-hand side of the screen following every five correct judgements. The importance of accuracy and not reaction time was stressed verbally by the experimenter prior to beginning the task.
The number of trials and duration of the stimulus was determined by a parameter estimation by sequential testing (PEST) procedure (Findlay, Citation1978; Pentland, Citation1980). A staircase method was adopted in which the stimulus duration was altered on the basis of the participant's previous responses. This resulted in a sensitive tracking procedure influenced by random responses and lapses in attention/interest. To minimise bias from these factors, IT was estimated post hoc by fitting a cumulative normal curve (M = 0) to accuracy as a function of stimulus onset asynchrony (SOA). It is commonly estimated via curve extrapolation so that performance at any desired accuracy level can be attained (Nettelbeck, Citation1987). The statistic of interest is the standard deviation of the curve, which is the SOA at which 84% accuracy is achieved. The test – retest reliability of this measure is 0.69 (Luciano et al., Citation2001b). Participants whose data provided a poor fit to the cumulative normal curve (R2 < 0.95) were excluded (thirty-three participants, or 2.8% of the sample).
Choice Reaction Time task.
This task was presented to the participants in the visual form of dripping taps. The participant was instructed to quickly press the appropriate computer key to stop a tap from dripping (see Luciano et al., Citation2001a).
To minimise between-subject practice and order effects the sequence of choice conditions was fixed in the order of four, two, then eight (Smith & Stanley, Citation1983). The number of trials presented in each of the two, four, and eight choice conditions was 96, 48, and 96, respectively. For all conditions, eight taps appeared on the monitor; those taps in use for the 2- and 4-choice conditions were made salient by brightening their colour. The output measure was the mean of log transformed RT trials (in milliseconds) for correct responses in each choice condition. Raw RT trials < 150 ms or > 2000 ms were excluded from the calculation of the mean. Test – retest correlations for mean RT have been estimated at 0.61, 0.50 and 0.70 for the respective 2-, 4-, and 8-choice conditions (Luciano, Citation2002). Correlations between mean RT and accuracy showed evidence of a speed – accuracy trade-off effect in all choice conditions: 2-choice (r = 0.21), 4-choice (r = 0.11), and 8-choice (r = 0.39). Mean RT was thus adjusted for percentage of correct responses in the means model of the genetic analysis by inclusion of a regression term for accuracy.
Delayed Response task.
The apparatus and conditions of the DR task have been described in Luciano et al (Citation2001a). Briefly, the DR task requires the participant to make a response using a rubber-tipped pointer (5 mm in diameter) on a touch-sensitive computer screen. When not responding, participants rested their hand with the pointer on a touch-sensitive response pad situated approximately 10 cm outward from the base of the monitor.
Each trial began with the appearance of a filled black circle in the centre of the screen, which was presented for 250 ms plus the delay period of 1 s or 4 s. The target (chequered circle) measured 1.5° visual angle in diameter and appeared at pseudo-random positions 250 ms from trial commencement. On control trials, the target remained on the screen for the delay period, while on memory trials the target appeared for 150 ms. The disappearance of the fixation point was the signal for the participant to make a response: touching the target on control trials or the memorised position of the target on memory trials. A distractor that looked like the target was randomly presented on half of the trials following target onset. Participants were instructed to ignore the distractor. During the variable intertrial interval participants received feedback on the screen. Correct responses were rewarded with 2, 4, 8, or 10 cents, depending on how close the response was to the centre of the target. Fast responses ( < 200 ms), slow responses ( > 1500 ms) and incorrect position responses incurred a 5-cent penalty. Six blocks of 72 trials were administered in the experimental session.
The accuracy of a response was indexed by the overall amount won on control and memory trials, hence this measure encompassed the correctness of a response as well as the position displacement on a correct response. A negatively skewed distribution with a ceiling effect was observed for percentage of correct responses on control trials, hence total winnings was primarily an index of memory accuracy. Reaction time was measured by (1) initiation time: interval from fixation point offset to removal of the hand from the response pad; and (b) movement time: interval from removal of the hand from the response pad to the screen response. Reaction Time measures were collapsed into sensory and memory conditions rather than maintained as eight individual conditions to ensure increased internal reliability. Only memory conditions are reported in this study. The median estimate across each individual's trials was used in preference to the mean, because it is generally less sensitive to outliers. Test – retest correlations were 0.46 for winnings, 0.67 for initiation time, and 0.61 for movement time (Luciano, Citation2002).
Analysis
All hypotheses concerning means, variances, correlations and components of variation and covariation were tested within the framework of maximum likelihood (ML) analysis of raw data using Mx 1.50 (Neale, Citation2000). Models were fitted to the data, progressing from the most saturated to more restricted models. Means and variances were tested for equality across birth order, zygosity and sex. Other mean effects tested for were age and accuracy (CRT variables only). Once a means model was decided on, hypotheses concerning homogeneity of correlations between sexes within zygosity groups were tested to check for evidence of sex limitation (i.e., different effect of genes/environment in female and male subjects).
Genetic modelling began from the approach of Cholesky decomposition of additive genetic (A), common environmental (C), and unique environmental (E) covariance between the measures. This specifies as many factors as there are variables for each source of variance, each factor having one loading less than the previous one. Reduced models (i.e., with fewer parameters) are favoured if the likelihood ratio chi-square comparing the models is less than the critical value (α = .05) of the chi-square distribution, indicating that there is no significant difference between the saturated model and the reduced model. The most parsimonious Cholesky decomposition (i.e., the one with the fewest sources [A, C, E] of variation) was used as a baseline against which to compare submodels with fewer factors or loadings using the chi-square difference test (Neale & Cardon, Citation1992).
Results
Descriptive statistics
Computer and/or experimenter error resulted in the loss of IQ data from six participants (0.56%), and DR data from nine (0.84%) participants. Inspection Time was transformed by a logarithmic (base 10) function, while all other variables were normally distributed. Scores were considered univariate outliers (single distribution of Twin 1 and Twin 2 scores) if they exceeded ± 3 SD from the mean. The largest proportion of outliers was for IT (4.5%) followed by 8-choice RT variable (2.1%). Four multivariate outliers (across twin families and all variables) were identified using the %P function in Mx and removed.
Contrasts of means and variances across birth order and zygosity showed differences between groups for the IQ variables and DR initiation time. This difference was in equating means within sex in DZ opposite-sex groups (e.g., the first-born female mean could not be equated with the second-born female mean in DZ opposite-sex groups). However, when the birth order in DZ opposite-sex groups was ignored, the mean for female subjects and male subjects in this group equalled the respective mean for female and male subjects in the rest of the sample. The DZ same-sex and DZ opposite-sex co-twin correlations could be equated for all variables, and all variables showed that MZ co-twin correlations were larger than DZ co-twin correlations, suggesting the presence of genetic effects and the absence of sex limitation.
The ML estimates of means, standard deviations and effect size of regression coefficients of the variables are shown in , along with MZ and DZ co-twin correlations, which have been adjusted for any significant effects of age, sex, and CRT accuracy. The IQ scores ranged from 83 to 144 for verbal IQ and 64 to 151 for performance IQ. The mean percentages correct in the 2-, 4-, and 8-choice conditions were 92.2, 82.1, and 79.7, respectively.
Table I. Maximum likelihood estimates for IT, CRT and DR variables, and verbal and performance IQs
The size of the difference between MZ and DZ twin correlations was more consistent with common environment effects (or assortative mating) rather than dominance effects. Thus, we tested only for the presence of additive genetic, common environmental and individual (or unique) environmental effects. Phenotypic correlations (estimated by ML) among the variables are presented in . Within-task correlations for CRT, as would be expected, were high. The processing speed measures were more strongly correlated with IQ than with the DR measures.
Table II. Maximum likelihood estimates of phenotypic correlations
Genetic analysis
Results of the Cholesky analysis are shown in . An additive genetic (A) and unique environmental (E) model best fitted the data, as evidenced by the nonsignificant chi-square change of the nested model (χ2 = 14.81, Δdf = 45, p = 1.0). A common and unique environmental model showed a significant change in chi-square and was not investigated further. A simplified theoretical model for the additive genetic structure was tested by a comparison with the AE Cholesky decomposition (). The genetic structure of this model comprised a general factor (loading on all variables), a CRT factor (2-, 4, and 8-choice RT), a DR genetic factor (initiation time, movement time, winnings), a spatial memory factor (DR winnings, performance IQ), an IQ factor (verbal and performance); specific genetic influences were parameterised for all variables, except DR winnings and performance IQ (to ensure model identification). This submodel did not fit the data, indicating that a more complicated pattern of genetic covariance existed among the variables. Thus an approach was taken that involved dropping nonsignificant pathways in the AE Cholesky decomposition to arrive at a simplified solution. This procedure involved testing the significance of each path coefficient individually (i.e., on one degree of freedom).
Table III. Results of fitting multivariate models to the covariances of IT, CRT, DR and IQ measures
A reduced AE Cholesky decomposition in which nonsignificant loadings were fixed to zero showed the presence of nine genetic factors. The standardised path coefficients of this factor decomposition are shown in . The first genetic factor loaded on all measures, except DR movement time, with its strongest loading on IT. The second genetic factor loaded on all variables except IT. The third genetic factor loaded on 4- and 8-choice RT and verbal and performance IQs. The fourth genetic factor influenced 8-choice RT and DR initiation time. The fifth, sixth and ninth genetic factors showed independent loadings on DR initiation time, DR movement time, and performance IQ. The seventh genetic factor influenced DR winnings and performance IQ, while the eighth factor influenced verbal and performance IQs. Heritability estimates ranged from 0.23 for DR movement time to 0.82 for verbal IQ.
Figure 1. Path diagram depicting the standardised path coefficients for the reduced additive genetic and unique environmental Cholesky decomposition of covariance among Processing Speed, Delayed Response (DR) Speed and Accuracy, and IQ Variables. Italic = heritability of each measure.
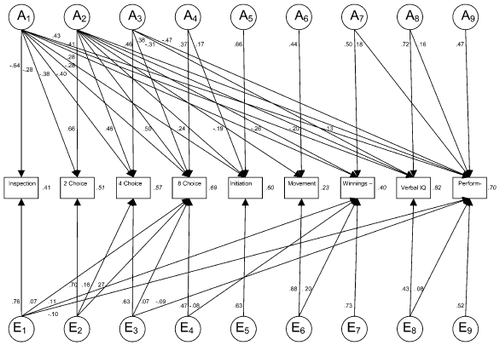
There was evidence for correlated unique environmental effects, with six factors having influence on multiple variables. In addition, specific environmental influences were observed for DR initiation time, DR winnings and performance IQ.
shows the relative influence from genes and environment on the phenotypic covariance between the varied cognitive measures. All relationships were mediated to a greater extent by genes, except for the association between DR winnings and DR movement time, where almost 80% of the covariance was environmental in origin. Also shown in are the genetic and environmental correlations estimated from the reduced AE Cholesky decomposition. The genetic correlations of the experimental cognitive measures with IQ were highest for 4-choice RT and lowest for DR movement time.
Table IV. Percentage of phenotypic covariance accounted for by genetic and unique environmental influences
Discussion
Although the relationship among processing speed, visuospatial working memory and IQ was primarily shown to be genetically mediated, a unitary factor was insufficient to explain the entire genetic covariance. A general genetic cognitive factor emerged, but several other genetic factors also accounted for large proportions of genetic covariation between varied cognitive measures.
The present results showed that, in general, processing speed was more strongly related to verbal and performance IQ (correlation range: – 0.22 to – 0.51) than was memory accuracy (correlations of 0.16 and 0.24 with respective verbal and performance IQs). Although the correlation between CRT and IQ was consistent with previous studies (perhaps slightly higher for 4- and 8-choice RT), the correlation between DR accuracy and IQ was lower than for other working memory tasks. In previous studies, working memory factors are typically shown to have a stronger correlation with IQ (range of ∼0.40 to 0.80) than with speed measures (e.g., Miller & Vernon, Citation1992, Citation1996). The lower correlation that we obtained may be because the DR task is less taxing on executive functioning than typical working memory tasks such as reading span. The phenotypic correlations between CRT measures and DR accuracy were similar to those reported for CRT and a spatial working memory task, that is, between 0.25 and 0.29 (Kyllonen, Citation1993). However, the phenotypic correlation between IT and DR accuracy was lower, indicating that processing speed could not be the only factor influencing the relationship between elementary cognitive tasks and memory performance. Perhaps a component process related to visual focus (which was larger in CRT and the DR task) or RT (also a factor in DR accuracy) increased the correlation between DR accuracy and CRT compared to IT.
Models in which additive genetic, common environmental and unique environmental components of variance were fitted to the data showed the best fitting model to be one without common environmental effects. Estimates of heritability agreed with previous findings of IQ, IT and CRT (Baker, Vernon, & Ho, Citation1991; Boomsma & Somsen, Citation1991; Luciano et al., Citation2001b; Posthuma, de Geus, & Boomsma, Citation2001) and the heritability of DR accuracy was consistent with studies of short-term memory (Finkel, Pedersen, & McGue, Citation1995; Thapar, Petrill, & Thompson, Citation1994).
The prediction of significant genetic covariance among processing speed, memory and IQ was supported, although this was not through a unitary genetic factor. The first genetic factor influenced all the measures (except DR movement time), with its largest loadings on IT, then IQ, 8-choice RT, 4-choice RT, and last, 2-choice RT, DR initiation time and DR winnings. Because IT is considered to be a purer measure of information processing speed than CRT, which is confounded with motor response, the first factor was theorised to represent a processing speed property of the brain that determines the efficiency or speed of very basic information processes, such as fine visual discrimination or encoding. This speed is also considered to increase the capacity of working memory, and hence IQ. This notion is further supported by the fact that DR movement time (a pure response time measure) was not influenced by this factor. A limitation of the Cholesky decomposition is that the first factor (genetic/environmental) must account for the entire variance (genetic/environmental) observed in the first variable entered into the model (i.e., IT). Although the Cholesky procedure indicated a single genetic factor influencing IT, we have shown in an alternative analysis that up to 21% of variance in IT is influenced by specific genes (Luciano et al., Citation2003).
The second genetic factor influenced all variables except IT and loaded most strongly on CRT variables. Because IT was the only task not requiring a speeded response, we theorised that this factor reflects those individual differences (e.g., anxiety, motivation, decision processes, strategy use) that influence performance under timed conditions. While the CRT task required fast responding, the DR and IQ tests both imposed time restrictions so that fast responding may have assisted performance. Delayed Response initiation time clocks the speed with which a person registers that the fixation point has disappeared (the cue to respond). A person with a faster initiation time will have more time to execute the manual response (movement time) before the lapse of the time limit; hence the negative relationship between these variables. Delayed Response accuracy indirectly taps speeded performance because responses that are too fast or too slow contribute to the variation in this measure, with the effect of reducing winnings. The Multidimensional Aptitude Battery combines speed and power (rewarding quantity and quality of performance), but it is not overly influenced by speed of responding and this is perhaps reflected in the comparatively lower factor loadings observed for IQ from the second genetic factor.
The third genetic factor influenced 4- and 8-choice RTs and IQ, so it is hypothesised to reflect processes related to choice decision. Because the direction of causation cannot be inferred from our data, it may be either that superior choice decision processing advantages higher order cognition or that people with higher IQs optimise their choice decision processes. Performance IQ actually showed the largest influence from this factor, so this factor may also relate more specifically to visuospatial content or alternatively, a speed (or ability) of learning function because performance subtests do not rely on prior knowledge. The fourth genetic factor influenced 8-choice RT and, to a lesser extent, DR initiation time and may capture motivational or persistence factors because the 8-choice condition was completed after the 2- and 4-choice conditions and IQ test. Because the DR task is lengthy (with six blocks of 72 trials), motivational factors may have also affected response speeds of this task, but the imposed time limit on each trial ensured that movement times were not affected. Delayed Response accuracy demonstrated a genetic relationship with performance IQ independent from the processing speed variables, signalling a unique relationship between memory processes and fluid ability that is not observed with crystallised ability. It may be that the genetic variance contributing to DR accuracy from this factor is related to the maintenance component of working memory, and it is the visuospatial nature of both the DR task and performance IQ subtests that gives rise to their specific genetic relationship. Our finding that memory accuracy is influenced to a greater extent by specific genetic factors rather than by a general factor is consistent with previous research (Cardon et al., Citation1992; Finkel, Pedersen, McGue et al., Citation1995; Luo et al., Citation1994).
Genetic factors emerged that showed specific influences on DR initiation time, DR movement time and performance IQ. Delayed Response initiation time was primarily influenced by specific genes (44% of variance). These genes may have been related to an automaticity response function because initiation was always in response to the same stimulus (fixation point disappearing) and could be paced for the 4-s delay condition. Participants also received twice the number of trials in the DR task than the CRT task and this may have additionally fostered automaticity of responding in the DR task. The genetic factor specific to DR movement time may reflect some physical attribute of the arm/hand, or alternatively it may relate to some perceptual timing mechanism because a time limit was imposed on responses.
Genetic correlations were especially strong between variables within the CRT task and between verbal and performance IQ. The genetic correlation between 4- and 8-choice RT was 0.89, indicating that the processes tapped by each measure rely virtually on the same gene (or set of genes), although environmental effects differ considerably (6% overlap). Moderate to high genetic correlations were observed between the IQ variables with the processing speed measures and DR accuracy. The strong genetic relationship between CRT and IQ was in accord with findings of Rijsdijk, Vernon, and Boomsma (Citation1998) who used a 2-choice RT task in a similar aged sample. They obtained a genetic correlation with the Raven Progressive Matrices of – 0.36, we found correlations ranging from – 0.38 (2-choice) to – 0.57 (4-choice) for verbal IQ and from – 0.34 (2-choice) to – 0.70 (4-choice) for performance IQ. Moderate genetic correlations were also observed between the processing speed measures and DR accuracy (in fact they were larger than the correlations between DR speed and DR accuracy). The genetic correlations between processing speed measures and DR accuracy were similar in magnitude to those reported (0.44 and 0.50) by Finkel and McGue (Citation1993), who used a speed factor and a host of short-term memory tasks.
Fifty-four per cent of the genetic variance in verbal IQ and 25% in performance IQ was actually independent of that determining the lower level processes sampled by the processing speed and DR tasks, suggesting that although speed and memory processes may contribute to higher order functioning, they are not sufficient to explain the entire genetic variation in IQ. The larger amount of independent genetic variance observed for verbal IQ compared to performance IQ is probably due to the visuospatial nature of the speed and DR tasks. For parsimony, common environmental effects were excluded from our final model, but point estimates from the additive genetic – common environmental – unique environmental (ACE) model indicated moderate common environmental effects on verbal IQ, and hence part of the observed independent genetic variance for verbal IQ may actually be environmental in origin.
We hypothesise that the independent genetic variance in IQ may be reflective of the overall efficiency of information processing abilities not tapped by the current tasks (e.g., access to long-term memory). For instance, Detterman (Citation2000) espouses a multifaceted model of general intelligence where g consists of many varied cognitive processes, which he further suggests independently contribute to brain processing to form an integrated system. Such an interpretation fits well with the pleiotropic perspective of gene function in intelligence, that is, the same genes influencing many diverse abilities. For a complete understanding of the individual variation in cognition, noncognitive factors such as personality characteristics (e.g., conscientiousness) must also be investigated. Without an increased number of informative variables in this analysis, such interpretation can only be speculative. However, information of this nature has been collected within the context of our study, and these questions will be addressed in the future.
Although there was some correlation of unique environmental effects between variables, the size of the first factor loading for every factor was disproportionately larger than any other factor loading, indicating that environmental effects were mostly variable specific. The largest environmental associations were among CRT measures and between DR movement time and DR winnings. Because tests were not perfectly reliable (Luciano et al., Citation2001a; Luciano et al., Citation2001b), some of the unique environmental variance may be composed of test measurement error.
In summary, the present study demonstrated that genetic factors influenced the relationship among processing speed, visuospatial memory and IQ variables, with further genetic factors unique to DR speed and performance IQ. There was support for multiple genetic factors influencing the covariation among speed, memory and IQ, indicating that there may be cognitive processes associated specifically with choice decision and with short-term memory storage that relate to IQ but not to each other. This view is in line with the notion of Detterman (Citation2000) that intelligence consists of varied independent information processes that combine to form an integrated cognitive network. We theorise that this integrated cognitive system can also manifest itself as a general factor, as reflected by the genetic general cognitive factor observed in this study.
The inclusiveness of the cognitive measures in this study will be informative for the next stage of this project, which involves the hunt for specific genes influencing cognitive ability. Our approach will be to search for genes affecting the component cognitive processes, which are arguably closer to the biological brain function than a complex (or higher-order) cognitive measure such as IQ. For instance, 4-choice RT is largely heritable (h2 = 0.57), shares 26% of variance with performance IQ (of which 86% is genetically mediated) and has a genetic correlation of – 0.70 with performance IQ. Identification of a gene for 4-choice RT may show that this same gene also influences IQ.
Acknowledgements
This project was supported by grants from the Australian Research Council (Grant Numbers: A79600334, A79906588, A79801419) and the Human Frontier Science Program (Grant Number RG0154/1998-B). We express our gratitude to Ann Eldridge and Marlene Grace for data collection and to the twins and their parents for their cooperation.
- Alarcon, M, Plomin, R, Fulker, DW, Corley, R, and DeFries, JC, 1998. Molarity not modularity: Multivariate genetic analysis of specific cognitive abilities in parents and their 16-year-old children in the Colorado Adoption Project, Cognitive Development 14 (1) (1998), pp. 175–193.
- Baker, LA, Vernon, PA, and Ho, H, 1991. The genetic correlation between intelligence and speed of information processing, Behavior Genetics 21 (4) (1991), pp. 351–367.
- Boomsma, DI, and Somsen, RJM, 1991. Reaction times measured in a choice reaction time and a double task condition: A small twin study, Personality and Individual Differences 12 (6) (1991), pp. 519–522.
- Cardon, LR, Fulker, DW, DeFries, JC, and Plomin, R, 1992. Multivariate genetic analysis of specific cognitive abilities in the Colorado Adoption Project at age 7, Intelligence 16 (1992), pp. 383–400.
- Carroll, JB, 1993. "Human cognitive abilities: A survey of factor-analytic studies". 1993, Cambridge: Cambridge University Press.
- Deary, IJ, and Stough, C, 1996. Intelligence and inspection time, American Psychologist 51 (6) (1996), pp. 599–608.
- Detterman, DK, 2000. General intelligence and the definition of phenotypes, 233 (2000), p. pp. 136 – 144, In G. R. Bock, J. A. Goode, & K. Webb (Eds.), The nature of intelligence, Chichester, UK: John Wiley & Sons.
- Findlay, JM, 1978. Estimates on probability functions: A more virulent PEST, Perception and Psychophysics 23 (2) (1978), pp. 181–185.
- Finkel, D, and McGue, M, 1993. The origins of individual differences in memory among the elderly: A behavior genetic analysis, Psychology and Aging 8 (4) (1993), pp. 527–537.
- Finkel, D, Pedersen, N, and McGue, M, 1995. Genetic influences on memory performance in adulthood: Comparison of Minnesota and Swedish twin data, Psychology and Aging 10 (1995), pp. 437–446.
- Finkel, D, Pedersen, NL, McGue, M, and McClearn, GE, 1995. Heritability of cognitive abilities in adult twins: Comparison of Minnesota and Swedish data, Behavior Genetics 25 (1995), pp. 421–431.
- Geffen, GM, Wright, MJ, Green, HJ, Gillespie, NA, Smyth, DC, Evans, DM, et al., 1997. Effects of memory load and distraction on performance and event-related slow potentials in a visuospatial working memory task, Journal of Cognitive Neuroscience 9 (1997), pp. 743–757.
- Goldman-Rakic, PS, 1992. Working memory and the mind, Scientific American 267 (1992), pp. 73–79.
- Harrell, TH, Honaker, M, Hetu, M, and Oberwager, J, 1987. Computerized versus traditional administration of the Multidimensional Aptitude Battery – Verbal Scale: An examination of reliability and validity, Computers in Human Behavior 3 (1987), pp. 129–137.
- Jackson, DN, 1984. "Manual for the Multidimensional Aptitude Battery". 1984, Port Huron, MI: Research Psychologists Press.
- Jackson, DN, 1998. "Multidimensional Aptitude Battery II". 1998, Port Huron, MI: Sigma Assessment Systems.
- Jensen, AR, 1993. Why is reaction time correlated with psychometric g, Current Directions in Psychological Science 2 (1993), pp. 53–56.
- Kaufman, AS, and Lichtenberger, EO, 1999. "Essentials of WAIS-III assessment". 1999, New York: John Wiley & Sons.
- Kyllonen, PC, 1993. Aptitude testing inspired by information processing: A test of the Four-sources Model, The Journal of General Psychology 120 (1993), pp. 375–405.
- Luciano, M, 2002. "An information processing approach to the genetic study of psychometric intelligence: Links between processing speed, working memory and IQ". 2002, Unpublished doctoral thesis, University of Queensland, Brisbane, Australia.
- Luciano, M, Smith, GA, Wright, MJ, Geffen, GM, Geffen, LB, and Martin, NG, 2001a. Genetic covariance among measures of information processing speed, working memory, and IQ, Behavior Genetics 31 (2001a), pp. 581–592.
- Luciano, M, Smith, GA, Wright, MJ, Geffen, GM, Geffen, LB, and Martin, NG, 2001b. On the heritability of inspection time and its covariance with IQ: A twin study, Intelligence 29 (2001b), pp. 443–457.
- Luciano, M, Smith, GA, Wright, MJ, Geffen, GM, Geffen, LB, and Martin, NG, 2003. "Genetic covariance between processing speed and IQ". 2003. p. (pp. 163 – 182), In R. Plomin, J. DeFries, P. McGuffin, & I. Craig (Eds.), Behavior genetics in the postgenomic era, Washington, DC: APA Books.
- Luo, D, Petrill, SA, and Thompson, LA, 1994. An exploration of genetic g: Hierarchical factor analysis of cognitive data from the Western Reserve Twin Project, Intelligence 18 (1994), pp. 335–347.
- MacLennan, RN, Jackson, DN, and Bellantino, N, 1988. Response latencies and the computerized assessment of intelligence, Personality and Individual Differences 9 (1988), pp. 811–816.
- Miller, LT, and Vernon, PA, 1992. The general factor in short-term memory, intelligence, and reaction time, Intelligence 16 (1992), pp. 5–29.
- Miller, LT, and Vernon, PA, 1996. Intelligence, reaction time, and working memory in 4- to 6-year-old children, Intelligence 22 (1996), pp. 155–190.
- Neale, MC, 2000. "Statistical Modeling with Mx". 2000, Richmond, VA 23298: Department of Psychiatry, Box 126 MCV.
- Neale, MC, and Cardon, LR, 1992. "Methodology for genetic studies of twins and families". 1992, Dordrecht, Netherlands: Kluwer Academic Publishers.
- Nettelbeck, T, 1987. "Inspection time and intelligence". 1987. p. (pp. 295 – 346), In P. A. Vernon (Ed.), Speed of information-processing and intelligence, Norwood, NJ: Ablex Publishing Corporation.
- Pentland, A, 1980. Maximum likelihood estimation: The best PEST, Perception and Psychophysics 28 (1980), pp. 377–379.
- Petrill, SA, Plomin, R, Berg, S, Johansson, B, Pedersen, NL, Ahern, F, et al., 1998. The genetic and environmental relationship between general and specific cognitive abilities in twins age 80 and older, Psychological Science 9 (1998), pp. 183–189.
- Posthuma, D, de Geus, EJC, and Boomsma, DI, 2001. Perceptual speed and IQ are associated through common genetic factors, Behavior Genetics 31 (2001), pp. 593–602.
- Rijsdijk, FV, Vernon, PA, and Boomsma, DI, 1998. The genetic basis of the relation between speed-of-information-processing and IQ, Behavioural Brain Research 95 (1998), pp. 77–84.
- Roberts, RD, and Stankov, L, 1999. Individual differences in speed of mental processing and human cognitive abilities: Toward a taxonomic model, Learning and Individual Differences 11 (1999), pp. 1–120.
- Salthouse, TA, 1996. The processing-speed theory of adult age differences in cognition, Psychological Review 103 (1996), pp. 403–428.
- Smith, GA, and Stanley, G, 1983. Clocking g: Relating intelligence and measures of timed performance, Intelligence 7 (1983), pp. 353–368.
- Thapar, A, Petrill, SA, and Thompson, LA, 1994. The heritability of memory in the Western Reserve twin project, Behavior Genetics 24 (1994), pp. 155–160.
- Thompson, LA, Detterman, DK, and Plomin, R, 1991. Associations between cognitive abilities and scholastic achievement: Genetic overlap but environmental differences, Psychological Science 2 (1991), pp. 158–165.
- Wright, MJ, De Geus, E, Ando, J, Luciano, M, Posthuma, D, Ono, Y, et al., 2001. Genetics of cognition: Outline of collaborative twin study, Twin Research 4 (2001), pp. 48–56.