Abstract
This study examined visual search for animal fear stimuli and whether high fear levels influence children's visual search. Experiment 1 was conducted with adults to provide a control for the effects observed in Experiments 2 and 3 with children. Both adults and children were faster to locate snakes and spiders among flowers and mushrooms than vice versa in arrays of nine but not of four pictures. Both groups were also faster to determine target absence from arrays of snakes and spiders than flowers and mushrooms regardless of array size. Experiment 3 showed that compared with low-fearful children, those who feared snakes and spiders did not show a search advantage for determining target absence from arrays containing snakes and spiders compared with flowers and mushrooms. These results support preferential search for animal fear stimuli in children and suggest that high fearfulness affects children's ability to disengage attention from feared stimuli.
Keywords:
From an evolutionary perspective, the human fear response observed in situations of impending danger and threat is part of an evolved defensive behavioural system that motivates organisms to escape and avoid sources of danger (e.g., Blanchard & Blanchard, Citation1988; Bolles, Citation1970; Fanselow & Lester, Citation1988). The evolutionary basis of fear activation have more recently been incorporated into information processing models of fear activation (e.g., Öhman, Citation1993). The central premise of these approaches is that for organisms to cope with the influx of information encountered in the environment, certain types of stimuli representing danger and harm are detected and processed preferentially over less important information (Mineka & Öhman, Citation2002). Mineka and Öhman propose the concept of an evolved “fear module” that selectively processes phylogenetically based fear stimuli, such as snakes, spiders and angry faces – stimuli that have long been associated with danger and threat to humans. Although fear activation can be triggered by ontogenetic fear stimuli, the effects have been strongest for phylogenetically based fear stimuli (Öhman & Mineka, Citation2001). Moreover, preferential selection of fear-relevant stimuli is activated automatically, such that conscious analysis of a fear stimulus is not necessary for the elicitation of a fear response.
A considerable theoretical literature has also developed proposing that stimuli depicting threat to the safety and wellbeing of humans are more likely to differentially capture the attention of highly anxious and fearful individuals in comparison with non-anxious controls (e.g., Mogg & Bradley, Citation1998; Williams, Watts, MacLeod, & Mathews, Citation1997). That anxious individuals disproportionately allocate attention to threat stimuli is considered a component of a maladaptive attempt to regulate negative emotion because it may interfere with habituation, thereby contributing to the maintenance of anxiety in the long term (Mogg & Bradley, Citation1998).
Support for the contention that phylogenetic fear stimuli, such as snakes and spiders and angry faces are preferentially processed by all individuals and attract greater attention in high anxious individuals comes from diverse sets of data, with strong support coming from studies using visual search tasks (e.g., Fox et al., Citation2000; Öhman, Flykt, & Esteves, Citation2001; Öhman, Lundqvist, & Esteves, Citation2001). Öhman, Flykt, et al. (Citation2001) conducted three experiments using an adapted version of the face-in-the-crowd methodology (e.g., Hansen & Hansen, Citation1988) to examine whether attention is drawn selectively to phylogenetically based fear stimuli. In Experiment 1 non-selected adults were faster to detect fear targets (snakes and spiders) among non-fear backgrounds (mushrooms and flowers) than non-fear targets among fear backgrounds in 3×3 arrays of nine pictures. They were also faster to determine that backgrounds of snakes and spiders contained no targets in comparison with backgrounds of mushrooms and flowers. In Experiment 2 in which the visual display varied from 2×2 to 3×3 arrays, the same pattern of results was obtained. Moreover, search times for fear targets did not increase but did so for non-fear targets when the number of distracting stimuli increased from three to eight. In Experiment 3 the effect observed in Experiment 2 was replicated: faster identification of fear targets irrespective of the number of distracting stimuli and faster determinations that targets were absent from snake and spider backgrounds in comparison with flower and mushroom backgrounds. In combination, these findings were proposed as suggestive of a genuine search asymmetry between fear and non-fear stimuli. Moreover, a major finding of Experiment 3 was that participants fearful of either snakes or spiders showed facilitated search for their feared stimuli but did not differ from controls in search for non-feared fear-relevant or fear-irrelevant targets.
These results provide support for evolutionary models of fear activation, which propose that humans are capable of preferentially processing phylogenetically based fear stimuli as part of an adaptive fear response (Mineka & Öhman, Citation2002). The faster identification of snakes and spiders in comparison with mushrooms and flowers is said to reflect that the fear relevance of these stimuli captured attention either as a result of a top-down strategy or as the result of a bottom-up stimulus-driven attentional process triggered by characteristic features of the particular stimuli (Wolfe, Citation1998). By contrast, the observed slower identification of a non-fear target among fear stimuli is suggestive of the utilisation of a serial search strategy. These findings have alternatively been argued to reflect that perceptual characteristics of snake and spider stimuli might make them easier to detect than mushrooms and flowers within a visual search paradigm (e.g., Pashler, Citation1987; Purcell, Stewart, & Skov, Citation1996; Wolfe, Citation1998). However, other studies that have used line drawings with face stimuli and have better controlled for perceptual features of stimuli have shown strong evidence of preferential search for phylogenetically based fear stimuli (e.g., Fox et al., Citation2000; Hadwin et al., Citation2003).
Furthermore, that participants highly fearful of either snakes or spiders in the Öhman, Flykt, et al. (Citation2001) study showed facilitated search for their specifically feared stimulus but did not differ from controls in search for non-feared fear-relevant or fear-irrelevant stimuli accords with cognitive models of anxiety that propose that high levels of fear and anxiety selectively drive attention to feared stimuli (e.g., Mogg & Bradley, Citation1998; Williams et al., Citation1997). These fear-related differences in search for snakes and spiders versus non-feared stimuli accord with similar findings in adults with social phobia who were faster to detect angry faces than happy faces presented in neutral backgrounds in comparison with controls (e.g., Gilboa-Schechtman, Foa, & Amir, Citation1999).
In an extension of the theoretical and empirical work with adults, researchers have begun to consider the influence of fear and anxiety upon attention allocation to fear and non-fear stimuli in children and adolescents. Such investigations have sought to examine the extent to which biased attention processes associated with anxiety disorders are stable across development and therefore may play a role in the causation and maintenance of anxiety over time. These studies have primarily utilised threat and neutral stimuli in either word or picture formats in visual-probe tasks and emotional Stroop tasks and the results have not been consistent. Several studies have shown a processing bias toward threat stimuli in anxious youths (e.g., Moradi, Taghavi, Neshat-Doost, Yule, & Dalgleish, Citation1999; Taghavi, Dalgleish, Moradi, Neshat-Doost, & Yule, Citation2003; Vasey, Daleiden, Williams, & Brown, Citation1995), whereas other studies have found that a bias toward threat stimuli was common to children in general (e.g., Ehrenreich & Gross, Citation2002; Kindt, Bierman, & Brosschot, Citation1997; Kindt, Brosschot, & Everaerd, Citation1997; Waters, Lipp, & Spence, Citation2004). The inconsistencies may stem from a number of differences between these studies, including the age of children assessed, the composition of clinical groups (e.g., high fearful vs. clinical groups), the type of stimuli used (e.g., pictures vs. words), and the content they depict (e.g., a mix of social and physical threat stimuli vs. a specific fear stimulus) (Vasey & McLeod, Citation2001).
Recently, Hadwin et al. (Citation2003) examined visual search efficiency for threatening, neutral and happy schematic face stimuli in 7 – 10-year-old children and the influence of high levels of trait anxiety. Notable advantages of that study in comparison with earlier research are that reliance upon reading capability was avoided by utilising picture stimuli (Vasey et al., 1996) and that the effects of individual differences in trait anxiety could be assessed in relation to children's processing of evolutionarily relevant fear stimuli (e.g., Mineka & Öhman, Citation2002). Of interest was children's reaction-time (RT) to determine the presence and absence of angry, happy and neutral schematic faces or cartoon drawings set among background faces with mixed up facial features within arrays of four, six, and eight items. Children were faster to detect angry faces than happy or neutral faces and were faster to decide that targets were present than absent. Furthermore, RT increased linearly with increases in set size. These results accord with the developmental literature on children's visual selective attention that demonstrates considerable top-down modulation in children in both parallel and serial processing, and that the latency of visual search increases with task difficulty in children (Taylor & Khan, Citation2000). Of interest was that increased levels of children's trait-anxiety produced significantly shorter RT when an angry face was absent from backgrounds compared with when a neutral or happy face was absent. This contrasts with findings from adults that have shown facilitated detection of feared targets in fearful adults rather than fear-related differences on target-absent trials (e.g., Gilboa-Schechtmann et al., Citation1999; Öhman, Flykt, et al., Citation2001). Because participants must decide when to terminate the search on target-absent trials rather than this being driven by target detection, Hadwin et al. proposed that high trait anxiety in children may have led to more rapid decisions about the absence of angry faces in order to disengage attention from angry stimuli as rapidly as possible. Taken together, the findings of Hadwin et al. suggest that children preferentially process stimuli depicting evolutionary fear relevance to humans, consistent with contemporary models of fear activation (e.g., Mineka & Öhman, Citation2002) and with empirical evidence from adults (e.g., Fox et al., Citation2000; Öhman, Flykt, et al., Citation2001). Furthermore, the Hadwin et al. study highlights that individual differences in trait anxiety in children may differentially influence children's decision thresholds about fear stimuli under visual search conditions.
The present study had two major aims. Given the paucity of research examining the relevance of evolutionary models of fear activation (e.g., Mineka & Öhman, Citation2002; Öhman, Citation1993; Öhman & Mineka, Citation2001) across the lifespan (e.g., Hadwin et al., Citation2003; Öhman, Flykt, et al., Citation2001, experiments 1 and 2), the first aim was to examine visual search efficiency for snakes and spiders in children. The second aim was to examine whether fear of snakes and spiders differentially influences children's visual search (e.g., Öhman, Flykt, et al., Citation2001, experiment 3; Hadwin et al., Citation2003). Addressing these aims is important for expanding our understanding of the evolutionary basis of human fear activation and whether aberrant processing of fear stimuli is characteristic of children with excessive levels of fear, as seen in anxious adults.
To address these aims systematically, Experiment 1 was conducted with non-selected adults to provide a control for the findings obtained in Experiments 2 and 3 with children. Based on the findings of Öhman, Flykt, et al. (Citation2001), it was expected that non-selected adults would be (a) faster to detect snakes and spiders among backgrounds of mushrooms and flowers than vice versa, with search efficiency for fear-relevant targets unaffected by the number of distracting items, and (b) faster to determine that backgrounds of snakes and spiders contained no targets in comparison with backgrounds of mushrooms and flowers.
Using the same paradigm with children in Experiment 2, we then tested the hypothesis that children would be faster to detect snakes and spiders among flowers and mushrooms than vice versa. However, based on the Hadwin et al. (Citation2003) findings with emotional face stimuli, it was expected that search for both fear and non-fear targets would be slowed by an increase in set size. It was also expected that children would be faster to determine that targets were absent from backgrounds of snakes and spiders than mushrooms and flowers. In Experiment 3 we aimed to replicate these basic search effects with another sample of children and also examine whether high fearfulness of snakes and spiders influenced search times for these stimuli (Hadwin et al., Citation2003; Öhman, Flykt, et al., Citation2001; experiment 3).
Experiment 1
Method
Participants
Participants were 48 undergraduate psychology students (27 women) aged between 17 and 44 years (M = 20.75 years; SD = 5.90) who participated on an informed consent basis to obtain course credit. Data from four participants were rejected because of computer malfunction or they did not follow instructions. Thus, analyses were based on data from 44 participants (26 female).
Apparatus/Stimuli
The tasks were presented on an IBM-compatible PC with a Dell Trinitron 43 cm colour monitor and controlled by a custom-designed software program run under DOS (graphics mode 800×600) to ensure millisecond timing accuracy. Reaction-time was recorded with a two-key button-box attached to the games port. The two buttons were 1 cm in diameter and placed 4 cm apart (centre to centre). Participants responded using the index and middle fingers of the preferred hand, with the response options relating to the two fingers rotated across participants. Fear stimuli were nine pictures of snakes and spiders, and the non-fear pictures were nine pictures of flowers and mushrooms ( ). The pictures were the same as used in previous studies (Lipp, Derakshan, Waters, & Logies, Citation2004) and were processed using Paint Shop Pro (version 6) to have a colour range of 256 colours (error diffusion method) applying the web-save standardised palette, and were resized to 260×195 pixels (subtending a visual angle of 7.52×5.97° if presented centred).
Figure 1. Examples of fear and non-fear-relevant pictures displayed in 3×3 and 2×2 arrays.

Procedure
After providing informed consent, the array tasks were completed individually together with three other tasks (see Waters et al., Citation2004, Citation2005), which included a dot-probe detection task, an emotional Stroop task and a psychophysiological task. The order of task completion was counterbalanced across participants.
Participants completed two separate array tasks: one containing arrays of four pictures (i.e., Array_4 task: 2 rows×2 columns) and one containing arrays of nine pictures (i.e., Array_9 task: 3 rows×3 columns). The arrays were displayed at eye level in the centre of the computer monitor approximately 5 m in front of the participant. The nine pictures in the Array_9 task subtended a visual angle of 22.42°×17.90° and the four pictures in the Array_4 task were presented in the four corner positions of the nine-picture array. The background space surrounding the arrays was black.
At the commencement of a trial, a white fixation cross (subtending a visual angle of 1.53°×1.53°) appeared in the centre of a black background for 1000 ms and disappeared with the onset of the array. Participants were asked to press buttons labelled “same” or “different” to indicate whether the pictures in the array were from the same picture category or if one was different. They were asked to respond as fast but as accurately as possible. RTs were measured in milliseconds from the onset of the array to the button press. The maximum presentation time of the arrays was 3000 ms in the Array_4 task and 4000 ms in the Array_9 task. If participants did not respond within these intervals, RT was recorded as missing and the next trial was automatically presented. The time between trials was 1000 ms.
The Array_4 task and the Array_9 task consisted of three experimental conditions. In the target-absent condition no target picture was presented within an array – all the pictures were from the same category. Thus, all pictures in an array were either of snakes, spiders, flowers, or mushrooms only. There were two target-present conditions, and in the first the target picture was mismatched with the category of the other pictures in the array. Either a snake or spider was presented within a background of mushroom or flower pictures, or a mushroom or flower was presented within a background of snake or spider pictures. In the second target-present condition, the target picture was from the same category as the other pictures in the array. Thus, a picture of a snake was presented among spiders and vice versa, or a picture of a mushroom was presented among flowers and vice versa. The latter target-present condition was included because the tasks used in the present experiment were derived from our previous work in which we were interested in the effects of differing background items on search times (Lipp et al., Citation2004).
For target-present conditions, target pictures were presented at each position within the arrays, balanced across trials, resulting in 12 and 27 target trials for each picture type (spiders, snakes, mushrooms and flowers) in the Array_4 and the Array_9 task, respectively. This resulted in 48 target trials in the Array_4 task and 108 target trials in the Array_9 task. To balance the designs, an equal number of no-target trials were constructed for each task (i.e., the target-absent condition), resulting in 96 trials for the Array_4 task and 216 trials for the Array_9 task, with the trials randomised to produce the final sequences. Ten practice trials including trials from all experimental conditions were randomly selected from the main trials and were completed before each task.
Response definitions, data screening and statistical analyses
Analyses were based on mean RT and mean percentages of errors, which included incorrect button presses and RTs longer than the maximum allowable time. Extreme RT scores (defined as deviating from the mean by more than two standard deviations) were identified on a case-by-case basis to take account of individual differences in RTs. These values were rescored as missing and not included in the analyses. The analyses were initially conducted with data included from the same target-present condition in which targets were of the same fear category (fear or non-fear) as the background pictures but of different species (e.g., a snake among spiders or a flower among mushrooms). However, the results showed that a snake or spider was identified faster than a mushroom or flower when embedded in backgrounds from the same fear category (e.g., a snake embedded in spiders; a flower embedded in mushrooms) and when embedded in backgrounds from the opposing fear category (e.g., a snake embedded in flowers; a mushroom embedded in snakes). We report on the effects of differing background items in detail in Lipp et al. (Citation2004). Because these analyses demonstrated that search for fear targets was more efficient than for non-fear targets regardless of the background items and to be consistent with Öhman, Flykt, et al. (Citation2001), the final analyses were based on data from the target-absent condition and the target-present condition in which the target was from the opposing fear category to the background (e.g., a flower or mushroom among snakes or spiders; or a snake or spider among flowers or mushrooms).
Response times and percentages of error were subjected to 2×2×2 (Array Size: Array_4, Array_9)×(Background: non-fear, fear)×(Target: absent, present) repeated measures analyses of variance. Multiple comparisons of means to follow-up on significant effects were made with t tests with the critical values derived from Sidak's tables to control against accumulation of alpha error (Rohlf & Sokal, 1981). The significance level was set at .05 for all analyses. As an estimate of effect size, partial eta squared () was calculated.
Results
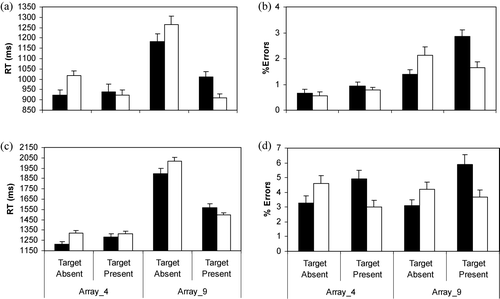
Mean RTs and standard errors for the Array_4 and Array_9 tasks are presented in
. Although participants were faster in the Array_4 task, they were quicker across both arrays to determine that targets were absent from fear than non-fear backgrounds. Adults were also faster to detect fear targets among non-fear backgrounds than vice versa. These observations were confirmed by main effects for Array Size, F(1,43) = 28.11, p < .001, and Background, F(1,43) = 87.30, p < .001,
, as well as interactions for Array Size×Target, F(1,43) = 20.09, p < .001,
, Array Size×Background, F(1,43) = 96.54, p < .001,
, Background×Target, F(1,43) = 86.56, p < .001,
, and Array Size×Background×Target, F(1,43) = 8.59, p = .002,
. Follow-up t tests of the three-way interaction were based on RTs for fear and non-fear backgrounds with and without targets for the Array_4 task and Array_9 tasks separately. In both array sizes, participants were significantly faster to determine that targets were absent from backgrounds of fear than non-fear pictures, both t(43) > 6.70. They were also significantly faster to find snakes and spiders in mushrooms and flowers than vice versa, but only in the larger arrays, t(43) = 8.64. The difference in RTs in the smaller arrays was not significant, t(43) = 1.28.
Figure 2. (a,c) Mean reaction times (RTs) and (b,d) percentages of errors (+SE) for (a,b) adults in Experiment 1 and (c,d) children in Experiment 2 during target-absent and target-present conditions as a function of the fear-relevance of the background items in the Array_4 and Array_9 tasks. In target-present conditions, targets are either snakes or spiders in non-fear-relevant backgrounds and mushrooms or flowers in fear-relevant backgrounds. (▪) Fear background; (□) non-fear background.
Öhman, Flykt, et al. (Citation2001) did not report overall analyses of variance such as those described above. Rather, to examine whether detection of fear and non-fear targets within opposite backgrounds was unaffected by an increase in distracting items, Öhman, Flykt, et al. (Citation2001) performed separate analyses for the target-present and -absent conditions across the two array sizes. We performed the same analyses to permit comparisons between the studies. For target-present trials, participants were faster to detect snakes and spiders within mushrooms and flowers than they were to detect mushrooms and flowers among snakes and spiders, F(1,43) = 52.39, p < .001, . However, a significant interaction, F(1,43) = 28.82, p < .001,
, indicated that the effect of fear relevance of targets differed as a function of array size. Similar to Öhman, Flykt, et al. (Citation2001), participants were slower to find mushrooms and flowers in the larger arrays than the smaller arrays, t(43) = 6.41, whereas the number of distracting stimuli did not affect speed to detect snakes and spiders, t(43) = 1.18. Also in accord with Öhman, Flykt, et al. (Citation2001), for arrays without targets, participants were faster to decide that a target was absent if the array was small, F(1,43) = 59.77, p < .001,
, and if the background was fear relevant than non-fear relevant, F(1,43) = 52.39, p < .001,
. The interaction was not significant, F = 0.48, ns., p > .05.
The percentages of errors committed in the two array tasks are displayed in
. The pattern of results resembles that for RTs, with longer RTs associated with more errors. This pattern is inconsistent with a speed – accuracy trade-off. The statistical analysis yielded main effects Array Size, F(1,43) = 64.21, p < .001, , Target, F(1,43) = 4.51, p = .03,
, and Background, F(1,43) = 20.69, p < .001,
, as well as a significant Array Size×Background interaction, F(1,43) = 20.11, p < .001,
, reflecting that more errors were committed in non-fear backgrounds in the larger than the smaller arrays. No other effects were significant (F < 1.50).
Discussion
The results of Experiment 1 can be summarised as follows: participants were faster to find snakes or spiders among flowers or mushrooms than they were to locate flowers or mushrooms among snakes or spiders, but this preferential effect was most evident in the larger array task. Also, whereas RTs to detect non-fear targets in fear backgrounds increased as the array size increased, RTs for snakes and spiders were unaffected by an increase in distracting items. Participants also were faster to establish that targets were absent from fear than non-fear backgrounds.
The current findings are consistent with those of Öhman, Flykt, et al. (Citation2001). Although the absence of a set size effect for the detection of fear-relevant targets such as snakes or angry faces has been discussed in terms of “pop-out”, which would indicate parallel processing (e.g., Hansen & Hansen, Citation1988), these search asymmetries are now typically discussed in terms of more and less efficient processing, which avoids speculation about the possible mode of processing (e.g., Fox et al., Citation2000). In accord, the present results show that participants were faster in shifting attention to fear-relevant targets and in disengaging attention (Posner & Petersen, Citation1990) from fear stimuli. One could argue that slower detection of non-fear targets in fear backgrounds compared to vice versa is because the search for mushrooms and flowers occurred against a background of snakes and spiders, which would themselves capture and hold attention at the expense of finding these targets. However, if this were the case, one would expect longer RTs to decide the absence of a target in snake and spider than in mushroom and flower backgrounds. The present findings, like those of Öhman, Flykt, et al. (Citation2001), revealed the opposite pattern: shorter RTs for snake and spider than mushroom and flower arrays without targets. Hence, the findings are consistent with the notion that there is a genuine search asymmetry for preferential processing of animal fear-relevant stimuli (Öhman, Flykt, et al., Citation2001). These findings replicate those of Öhman, Flykt, et al. (Citation2001) and support the contention that phylogenetically based fear stimuli are processed preferentially (Öhman & Mineka, Citation2001).
The study also found that within the smaller arrays, time to locate snakes and spiders in backgrounds of mushrooms and flowers did not differ significantly from time to identify mushrooms and flowers among snakes and spiders. This finding cannot be compared with previous research because Öhman, Flykt, et al. (Citation2001) did not report this contrast. This result suggests that with only three distracting items, there was no advantage for detecting fear targets over non-fear targets. Previous research has shown that efficiency of visual search can be affected by characteristics other than fear relevance, such as the degree of target-distractor discriminability (Wolfe, Citation1998) and by whether the number of items in the array is small or large (Pashler, Citation1987). Thus, although fear targets among non-fear backgrounds were just as rapidly detected in the smaller compared with the larger arrays, the equivalence of RTs for fear and non-fear targets in the smaller arrays suggests that when the set size is relatively small (i.e., four items or fewer), humans are capable of rapidly discriminating stimuli that are discrepant from their background, irrespective of fear valence.
Experiment 2
In an extension of Experiment 1 and those of Öhman, Flykt, et al. (Citation2001), we sought to examine whether snakes and spiders were detected more efficiently than mushroom and flower targets in non-selected children. Based on evidence of increased search times for emotional faces as a function of increased set size in children (Hadwin et al., Citation2003), it was expected that children would be faster to detect snakes and spiders than mushrooms and flowers within opposing backgrounds, but that search for targets would be longer when the set size increased. For arrays without targets, it was expected that children would be faster to determine that targets were absent from backgrounds of snakes and spiders than mushrooms and flowers.
Method
Participants
A sample of 105 non-selected children aged between 9 and 12 years (M = 10.59 years; SD = 1.00) participated. Children were recruited from a local primary school following Education Department approval and parental consent. Only children who were reported by parents to be free from intellectual impairment or a learning disorder and who spoke English as their first language were included. Because data from one participant were rejected due to high rates of extreme scores, analyses were based on data from 104 participants (63 girls).
Apparatus and procedures
The study was introduced to children in their classrooms and interested children gave their parents a research information package and returned the completed forms if parental consent was given. Children completed the array tasks and the three other tasks described in Experiment 1 (Waters et al., Citation2004, Citation2005) individually in a resource room in one of two sessions set a week apart. The order of task completion was counterbalanced across and within the sessions. The equipment was the same as in Experiment 1. The tasks were explained prior to the practice trials and children were given 30-s breaks every 3 min. Each session lasted approximately 50 min.
Because speed to locate fear targets in non-fear backgrounds was not significantly different to speed to locate non-fear targets in fear backgrounds in the smaller arrays in Experiment 1, the picture display was modified in Experiment 2 so that the four pictures were presented in the centre of the screen (subtending a visual angle of 15.07°×12.0°). In previous research (Lipp et al., Citation2004), we found that search for snakes and spiders over mushrooms and flowers within opposite backgrounds was significantly quicker when the four pictures were presented edge to edge, which is similar to the picture configuration in the Array_9 task. It should be noted that this configuration did not require participants to shift attention back and forth on consecutive trials over varying visual angles: Array_4 and Array_9 trials were presented as separate tasks in counterbalanced order rather than interspersed within the same task as in the Öhman, Flykt, et al. (Citation2001) study.
To take account of children's concentration abilities, the Array_9 task was shortened from 216 to 164 trials by reducing the number of no-target trials matched for each Target condition from 27 to 14. This prevented direct comparisons with adults' RTs in Experiment 1. However, the aim was to examine visual search efficiency for phylogenetically based fear stimuli in children using a within-subject design, in accordance with Öhman, Flykt, et al. (Citation2001). The response definitions and statistical analyses were the same as in Experiment 1.
Results
Children's mean RTs and standard errors in the two array tasks are presented in
. As shown, children were faster to respond if the arrays were small, if a target was present and if the arrays contained fear rather than non-fear items. In support, main effects were observed for Array Size, F(1,103) = 393.09, p < .001, , Target, F(1,103) = 127.73, p < .001,
, and Background, F(1,103) = 24.11, p < .001,
. However,
indicates that although children were faster across both array sizes to determine that targets were absent in fear than in non-fear backgrounds, their responses in the target-present condition varied across the two tasks. In support, the overall Array Size×Background×Target interaction was significant, F(1,103) = 20.38, p < .001,
. Significant effects were also obtained for Background×Target, F(1,103) = 71.40, p < .001,
, Array Size×Target, F(1,103) = 308.96, p < .001,
, and Array Size×Background, F(1,103) = 11.88, p = .001,
.
Follow-up t tests of the three-way interaction revealed that regardless of array size, children were faster to decide that targets were absent from fear than non-fear backgrounds, both t(103) > 9.80. Contrary to Öhman, Flykt, et al. (Citation2001) results but consistent with Experiment 1, children were faster only in the larger arrays to detect snakes and spiders among mushrooms and flowers than vice versa, t(103) = 5.88. The effect was not significant in the smaller arrays, t(103) = 2.47.
As in Experiment 1, separate analyses of RTs across both array sizes for target-absent and target-present conditions were performed to permit comparison with the Öhman, Flykt, et al. (Citation2001) results. For target-present trials, children's RTs were shorter in the smaller arrays than the larger arrays, F(1,103) = 110.82, p < .001, . However, a significant interaction, F(1,103) = 35.90, p < .001,
, indicated that the effect of fear relevance of targets differed as a function of array size. Similar to results reported by Öhman, Flykt, et al. (Citation2001) and in Experiment 1, children were significantly slower to detect non-fear targets in backgrounds of fear pictures in the larger arrays than the smaller arrays, t(103) = 24.24. However, children's speed to detect fear targets among non-fear backgrounds was also detrimentally affected when the number of distracting items increased, t(103) = 15.76. The interaction reflected that speed to detect fear and non-fear targets was significantly different in the larger, t(103) = 5.88, but not the smaller arrays, t(103) = 2.47. For arrays without targets, children were faster to decide that a target was absent if the array was small, F(1,103) = 505.65, p < .001,
, and if the background items were fear than non-fear relevant, F(1,103) = 63.06, p < .001,
. The interaction was not significant, F(1,103) = 0.18, p > .05.
The percentages of errors committed in the two array tasks are displayed in
. As in Experiment 1, the pattern of results resembles that for RTs, with longer RTs associated with more errors. This pattern is inconsistent with a speed – accuracy trade-off. The statistical analysis yielded a significant Target×Background interaction, F(1,103) = 24.97, p < .001, . For target-present trials, more errors were committed when the target was non-fear than fear relevant, t(103) = 3.43. For trials without targets, more errors were committed when searching through non-fear than fear backgrounds, t(103) = 3.06. No other effects were significant (F < 3.00).
Discussion
The results of Experiment 2 demonstrated that preferential processing of animal fear-relevant stimuli was characteristic of young children, as seen in adults. Like adults in Experiment 1 and in Öhman, Flykt, et al. (Citation2001), children were faster to detect snakes or spiders among mushrooms or flowers than vice versa. Similarly, this effect was significant in the larger Array_9 task but not in the Array_4 task. Although search times for both types of targets were detrimentally affected when the number of distracting stimuli increased, this effect was less pronounced for fear targets in non-fear backgrounds. Hadwin et al. (Citation2003) reported a similar set size effect with children, in that search for angry faces was prolonged when the number of distracting items increased. Thus, consistent with Hadwin et al., children's preferential detection of fear targets most likely occurred in a serial search process that took longer in the larger arrays. Finally, children were also faster to determine that targets were absent from arrays of snakes and spiders than of mushrooms and flowers.
The combined findings are among the first to demonstrate that evolutionary-relevant fear stimuli are processed preferentially from early in development (Mineka & Öhman, Citation2002). However, together with results from Experiment 1, the present findings indicated that among three distracting stimuli, fear relevance was not the only stimulus characteristic influencing children's visual search. Other distinguishing characteristics, such as the small number of distracting items (Pashler, Citation1987; Purcell et al., Citation1996; Wolfe, Citation1998) also influences speed of visual search even in young children.
Experiment 3
In Experiment 3 we aimed to replicate the finding of preferential search for fear targets and to examine whether children's visual search for snakes and spiders was affected by high versus low levels of snake and spider fearfulness. Öhman, Flykt, et al. (Citation2001; experiment 3) showed that adults specifically fearful of snakes but not spiders (and vice versa) showed facilitated search for their feared stimulus regardless of the number of distracting stimuli but did not differ from controls in search for non-feared fear-relevant or fear-irrelevant targets. Children in Experiment 3 were not permitted to be out of class longer than 30 min to complete the experimental session due to heavy extracurricular activities at the time of data collection. Because Experiment 2 confirmed that children could not complete the Array_4 task, the Array_9 task and the questionnaires within this timeframe, the Array_4 task was dropped from Experiment 3 on the basis that results from Experiments 1 and 2 and those reported in Lipp et al. (Citation2004) showed that preferential visual search for snakes and spiders was not stable in the Array_4 task. Although this prevented the examination of set size effects, it permitted us to examine the influence of children's fear upon search for snake and spider stimuli within the Array_9 task.
Method
Participants
Eighty-one children (51 girls) aged between 10 and 12 years (M = 11.02 years; SD = 0.86) participated and were recruited from the same primary school as in Experiment 2. No child participated in both experiments. The inclusion criteria were the same as in Experiment 2.
Apparatus and materials
The equipment, picture stimuli and parameters of the Array_9 task were the same as in Experiment 2. Children completed the Snake Fear Questionnaire (SNAQ) and the Spider Fear Questionnaire (SPQ) (Klorman, Weerts, Hastings, Melamed, & Lang, Citation1974), which were designed to assess specific fears of snakes and spiders. Both measures have good psychometric characteristics and are frequently used in research and clinical practice. On both the SNAQ and the SPQ, participants respond “true” or “false” to indicate their attitudes towards and beliefs about snakes and spiders (e.g., “I avoid going to parks or on camping trips because there may be snakes about”). Although there is a shortened version of the Spider fear questionnaire that has been adapted for use with children (e.g., Kindt, Brosschot, & Muris, Citation1996), there is not an equivalent shortened version of the Snake fear questionnaire. Hence, the full scales were utilised in the present experiment given that percentile cut-off scores needed to be specified on both measures so that high- and low-fearful groups could be identified. Children had no difficulty completing these measures.
Parents' ratings of their child's fear of snakes and spiders were also assessed via phone contact to provide corroborating information about children's self-reported fears. Parents provided ratings of their child's snake and spider fear using a 0 – 8 fearfulness scale (0 = not at all fearful, 2 = a little, 4 = somewhat, 6 = a lot, 8 = extremely fearful). The scale was explained to parents over the phone and their understanding of the scale was checked before they provided their ratings.
Overall anxiety was measured using the child and parent versions of the Spence Children's Anxiety Scale (SCAS-C; SCAS-P; Nauta et al., Citation2004; Spence, Citation1998). The SCAS-P and the SCAS-C assess children's anxiety in accordance with the symptom clusters specified in the Diagnostic and Statistical Manual of Mental Disorders (4th ed.; DSM-IV). Both versions of the SCAS have good psychometric properties (Nauta et al., Citation2004; Spence, Citation1998).
Procedure
Children were invited to participate in Experiment 3 in the same manner as in Experiment 2. Following the return of consent forms and questionnaires, parents were contacted by phone and their ratings of their child's fear of snakes and spiders were obtained. Children were assessed individually in a resource room during regular class time within 1 week of the call with parents. Children completed the Array_9 task first followed by the three questionnaires completed in rotated order across participants, to avoid either the snake or spider questionnaire always being completed first. The questionnaires were completed after the computer task to avoid drawing children's attention to snakes and spiders beforehand. Children completed 10 practice trials before the Array_9 task and were given 30-s breaks every 3 min.
Response definitions and data analysis
The response definitions and analysis for the full sample of 81 children were the same as in Experiment 2, with the exception that the Array Size factor was not included. The specific hypotheses relating to children's fearfulness were (a) that high fear would produce faster search times for feared than non-feared stimuli in target-present trials, as seen in adults (e.g., Öhman, Flykt, et al., Citation2001), and (b) that in target-absent trials, high fear would result in faster decisions about the absence of feared stimuli from non-fear backgrounds than vice versa (e.g., Hadwin et al., Citation2003). These specific hypotheses were assessed in a mixed 2×2 (Target Valence: fear, non-fear×Group: low, high) analysis of variance for target-present trials and a mixed 2×2 (Background: fear, non-fear×Group: low, high) analysis of variance for target-absent trials.
Results
Visual search performance for the full sample of 81 children
presents children's mean RTs and standard errors during target-absent and -present conditions as a function of the fear relevance of the background. As can be seen, children were slower on target-absent than -present trials, F(1,80) = 195.43, p < .001, . Moreover, a significant interaction reflected that speed of target detection varied depending on the fear-relevance of both target and background, F(1,80) = 49.91, p < .001,
. Children were faster to determine that targets were absent from fear than non-fear backgrounds, t(80) = 4.23, and to determine that fear targets were embedded in non-fear backgrounds than vice versa, t(80) = 6.34.
Figure 3. (a) Mean reaction times (RTs) and (b) percentages of errors (+SE) for children in Experiment 3 during target-absent and -present conditions as a function of the fear-relevance of the background items in the Array_9 task. In target-present conditions, targets are either snakes or spiders in non-fear-relevant backgrounds and mushrooms or flowers in fear-relevant backgrounds. (▪) Fear background; (□) non-fear background.
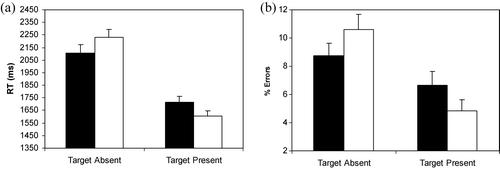
The percentages of errors in the Array_9 task are shown in
. Because slower response times were associated with more errors, this pattern is inconsistent with a speed – accuracy trade-off. The statistical analysis yielded a main effect of Target, F(1,80) = 30.45, p < .001, , as well as a Target×Background interaction, F(1,80) = 11.15, p < .001,
. As in Experiment 2, for target-present trials, significantly more errors were made when targets were non-fear than fear relevant t(80) = 2.94. For trials without targets, significantly more errors were made when searching through non-fear than fear backgrounds, t(80) = 2.11.
Analyses of questionnaire measures for high- and low-fearful children
Children were defined as highly fearful if their scores on the SPQ and the SNAQ were above the 75th percentile. For girls, this was a score of ≥15 on the SPQ and ≥16 on the SNAQ. For boys, this was a score of ≥7 on the SPQ and ≥9 on the SNAQ. Öhman, Flykt, et al. (Citation2001) also reported using different cut-off scores for determining high and low fear in male and female adults. Like Öhman, Flykt, et al. (Citation2001), we planned to select children specifically fearful of either spiders or snakes but not of both animals by selecting children whose scores on the non-feared fear-relevant scale were below the median. However, most children who were fearful of one animal also were fearful of the other, meaning that fearful children's scores were above the 75th percentile on both scales. Of the 18 children who met this criterion, 13 rated their fear of snakes to be stronger than their fear of spiders. Öhman, Flykt, et al. (Citation2001) defined low fearfulness as participants who scored below the median on both the spider and snake fear questionnaires. However, to ensure we compared the highest with the lowest fearful children in this non-selected sample, we compared high-fearful children with those scoring below the 25th percentile on both measures.
This resulted in 11 high-fearful and 10 low-fearful girls and seven high-fearful and seven low-fearful boys. As shown in , scores for girls and boys combined on the SNAQ and the SPQ were significantly different between high- and low-fearful children, t(33) = 10.06, p < .001 and t(33) = 8.19, p < .001, respectively. Parents' ratings of high-fearful children's fear of snakes were significantly different to the ratings of parents of low-fearful children, t(31) = 2.94, p = .006, and approached significance for fear of spiders, t(31) = 1.79, p = .08. Parents of fearful children also rated their children as generally more anxious on the SCAS-P compared with the ratings of parents of non-fearful children, t(33) = 3.15, p < .001. Similarly, high-fearful children rated themselves as generally more anxious on the SCAS-C, t(33) = 4.28, p < .001.
Table I. Child- and parent-report questionnaire measures for high- and low-fearful children
Visual search task analyses for high- and low-fearful children
Mean RTs and standard errors for high- and low-fearful children (boys and girls combined) are presented in
. The analysis for target-present trials indicated a significant Target Valence main effect, F(1,33) = 16.58, p < .001, , indicating that children in general were faster to detect fear targets within non-fear backgrounds than vice versa. The interaction and the Group main effect were not significant, F(1,33) = 2.80, p > .05, and F(1,33) = 0.50, p > .05).
Figure 4. Mean reaction times (RTs) and percentages of errors (+SE) for low-fearful and high-fearful children in Experiment 3 during target-absent and -present conditions as a function of the fear-relevance of the background items in the Array_9 task. In target-present conditions, targets are either snakes or spiders in non-fear-relevant backgrounds and mushrooms or flowers in fear-relevant backgrounds. (▪) Fear background; (□) non-fear background.
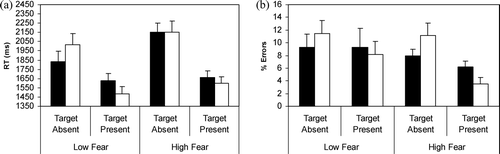
For arrays without targets, the analysis indicated a significant main effect of Background, F(1,33) = 4.52, p = .04, , which was subsumed by a significant interaction between Background and Group, F(1,33) = 4.27, p = .04,
. The Group main effect was not significant, F(1,33) = 1.91, p > .05. Follow-up t tests of the interaction confirmed observations in
that low-fearful children were significantly faster to determine that backgrounds of snakes or spiders contained no targets in comparison with backgrounds of mushrooms or flowers, t(33) = 3.02. By contrast, the search advantage through fear backgrounds in comparison with non-fear backgrounds was not observed for high-fearful children, t(33) = 0.05.
The analyses of the percentage of errors as a function of fear group status (
) showed that on target-present trials, both groups tended to make more errors when the target was non-fear than fear relevant, main effect of Target, F(1,33) = 3.29, p = .07, . The interaction and Group main effect were not significant (F < 1.56). On target-absent trials, both groups tended to make more errors when the background was non-fear than fear relevant, main effect of Target, F(1,33) = 3.05, p = .09,
. The other effects were not significant (F < 0.20).
Post-hoc analyses
Additional analyses were performed with children's SCAS-C scores to confirm that visual search was influenced by fear of snakes and spiders and not by general anxiousness. The same analyses were performed using visual search data from children scoring above the 75th and below the 25th percentiles on the SCAS-C, but no significant effects were obtained involving SCAS-C groups (all F < 0.45).
Also, when RTs were re-analysed with data excluded from the 18 high-fearful children (i.e., N = 63), a significant Target main effect, F(1,62) = 140.31, p < .001, , and Target×Background interaction, F(1,62) = 55.78, p < .001,
, were observed, consistent with the analyses for the full sample of 81. Children were faster to determine that targets were absent from fear backgrounds (M = 2080.70; SD = 623.82) than non-fear backgrounds (M = 2221.32, SD = 622.92), t(62) = 4.76, and to determine that fear targets were embedded in non-fear backgrounds (M = 1596.84; SD = 427.09) than vice versa (M = 1722.25, SD = 434.68), t(62) = 6.99.
Discussion
The results of Experiment 3 showed that children were faster to detect snakes and spiders among mushrooms and flowers than vice versa, and they were faster to detect target absence from fear compared with non-fear backgrounds in the Array_9 task. Moreover, the advantage for searching through fear backgrounds over non-fear backgrounds to determine that a target was absent was not characteristic of children fearful of snakes and spiders as it was of low-fearful children.
The results for the full sample of 81 children replicated those of Experiments 1 and 2. In so doing, the present findings support the contention that humans as young as 9 years of age are capable of preferentially detecting phylogenetically based fear stimuli (Mineka & Öhman, Citation2002), particularly when visual conditions are more complex. Most importantly, that children with higher levels of fear of snakes and spiders did not show the search advantage to detect target absence in arrays of snakes and spiders compared with flowers and mushrooms suggests that children's fear interfered with the ability to shift attention rapidly across snake and spider pictures to determine they were all the same. No evidence was found, however, for facilitated search for spider or snake targets embedded in flower or mushroom arrays compared to vice versa in high- relative to low-fearful children. Such an effect would be expected based on previous studies with adults (Öhman, Flykt, et al., Citation2001). However, Öhman, Flykt, et al. (Citation2001) tested the hypothesis that fearful adults would be faster to detect their feared target (e.g., a spider for a spider-fearful participant) in comparison with non-feared fear-relevant targets (e.g., a snake for a spider-fearful participant) using a planned contrast with participants who were specifically fearful of one animal but not the other. Because children in the present high-fearful group were fearful of both snakes and spiders and were identified from within a non-selected sample rather than based on specific fear scores, such a contrast could not be performed.
Nevertheless, in conjunction with the Hadwin et al. (Citation2003) study with children, the present results suggest that developmental considerations need to be taken into account. Similar to the present findings, Hadwin et al. did not find significant visual search differences for angry faces on target-present trials as a function of children's trait anxiety. Rather, children's trait anxiety influenced visual search for angry faces on target-absent trials. Hadwin et al. concluded that this may reflect that when targets are absent, children must decide when to end the search (i.e., to disengage attention) rather than this being driven by target detection.
Hadwin et al. (Citation2003) also found that high trait anxiety was associated with faster decisions that an angry face was absent from arrays of faces with mixed up facial features, whereas fearful children's search in the present experiment was slowed through arrays of all snakes or spiders to decide target absence. A notable difference between the studies is that target-absent trials in the Hadwin et al. study involved searching putatively neutral arrays (i.e., mixed up faces) to decide that an angry, happy, or neutral target was absent, whereas the present experiment involved searching fearful arrays (i.e., snakes and spiders) to decide that a neutral target (i.e., mushroom or flower) was absent and vice versa. Thus, the Hadwin et al. findings may reflect quicker decisions to terminate the search (i.e., disengage attention) to avoid excessive dwell-time on angry faces, whereas the present findings suggest that when confronted with feared stimuli, high-fearful children may have difficulty disengaging their attention from these stimuli (Posner & Petersen, Citation1990).
Attention engagement with and disengagement from threat stimuli have not been examined as a function of anxiety status in children, but recent studies with adults have shown that the component of attention underlying attention bias toward threat stimuli may be slowed disengagement of attention from threat (e.g., Koster, Crombez, Verschuere, & De Houwer, Citation2004; Yiend & Mathews, Citation2001). The present findings are important for extending our understanding of the role of cognitive factors in anxiety and fear states in children. Moreover, they accord with cognitive models of fear and anxiety that propose that biases in processing threat stimuli play a contributory role to anxiety (e.g., Mogg & Bradley, Citation1998; Williams et al., Citation1997). Furthermore, the present findings suggest that visual search may be a useful paradigm for differentiating the effects of fear and anxiety on the processing of fear stimuli in children. Future studies would benefit from examining visual search in children who are fearful of either snakes or spiders but not of both animals in comparison with controls. Such an investigation would elucidate the extent to which the degree of specific fear relevance and the nature of the search task (i.e., deciding about target-absence vs. stimulus-driven detection of a target's presence) are factors that permit the effects of children's fearfulness to become apparent (Hadwin et al., Citation2003). Furthermore, because only the Array_9 task was completed in the present experiment, further studies are required to determine whether snake and spider fearfulness influences children's search efficiency when the number of distracting stimuli varies.
Conclusions
Across three experiments, the present study examined whether preferential visual search for fear stimuli, shown repeatedly in adults, is also found in children and whether high fear levels influence children's visual search. These experiments showed that children detected snakes and spiders in mushrooms and flowers significantly faster than mushrooms and flowers within snakes and spiders, particularly in more complex arrays and most likely through a serial search process. At lower levels of visual complexity it appears that target-distracter discriminability as well as the fear relevance of targets can influence visual search efficiency, even in young children.
These results extend recent models of preferential processing of phylogenetically based fear stimuli (e.g., Mineka & Öhman, Citation2002) by showing that snakes and spiders are capable of selectively capturing the visual attention of children as young as 9 years of age. From an evolutionary perspective, these results provide evidence that when confronted with stimuli depicting danger and threat, the human perceptual system, which is part of an adaptive defensive behavioural system that motivates organisms to escape and avoid danger (e.g., Blanchard & Blanchard, Citation1988; Bolles, Citation1970; Fanselow & Lester, Citation1988), functions relatively efficiently very early in life, although most likely continues to improve with development.
Together with the finding of Hadwin et al. (Citation2003), the present findings indicate that high fear and trait anxiety interfere with children's search efficiency, which, depending on task demands, may reflect on underlying differences in the disengagement of attention from feared stimuli. Future research would benefit from examining whether these findings can be replicated in children with clinically significant phobias and anxiety, particularly when the number of distracting stimuli varies.
References
- Blanchard, D. C., and Blanchard, R. J., 1988. Ethoexperimental approaches to the biology of emotion, Annual Review of Psychology 39 (1988), pp. 43–68.
- Bolles, R. C., 1970. Species-specific defense reactions and avoidance learning, Psychological Review 77 (1970), pp. 32–48.
- Ehrenreich, J. T., and Gross, A. M., 2002. Biased attention behaviour in child anxiety: A review of theory and current empirical investigation, Clinical Psychology Review 22 (2002), pp. 991–1008.
- Fanselow, M. S., and Lester, L. S., 1988. "A functional behavioristic approach to aversively motivated behaviour: Predatory imminence as a determinant of the topography of defensive behaviour". In: Bolles, R. C., and Beecher, M. D., eds. Evolution and learning. Hillsdale, NJ: Erlbaum; 1988. pp. 185–212.
- Fox, E., Lester, V., Russo, R., Bowles, R. J., Pichler, A., and Dutton, K., 2000. Facial expressions of emotion: Are angry faces detected more efficiently?, Cognition and Emotion 14 (2000), pp. 61–92.
- Gilboa-Schectman, E., Foa, E. B., and Amir, N., 1999. Attentional biases for facial expression in social phobia, Cognition and Emotion 13 (1999), pp. 305–318.
- Hadwin, J. A., Donnelly, N., French, C. C., Richards, A., Watts, A., and Daley, D., 2003. The influence of children's self-report trait anxiety and depression on visual search for emotional faces, Journal of Child Psychology and Psychiatry 44 (2003), pp. 432–444.
- Hansen, C. H., and Hansen, R. D., 1988. Finding the face in the crowd: An anger superiority effect, Journal of Personality and Social Psychology 54 (1988), pp. 917–924.
- Kindt, M., Brosschot, J. F., and Muris, P., 1996. Spider phobia questionnaire for children (SPQ-C): A psychometric study and normative data, Behaviour Research and Therapy 34 (1996), pp. 277–282.
- Kindt, M., Bierman, D., and Brosschot, J. F., 1997. Cognitive bias in spider fear and control children: Assessment of emotional interference by a card format and a single-trial format of the Stroop task, Journal of Experimental Child Psychology 66 (1997), pp. 163–179.
- Kindt, M., Brosschot, J., and Everaerd, W., 1997. Cognitive processing bias of children in a real life stress situation and a neutral situation, Journal of Experimental Child Psychology 64 (1997), pp. 79–97.
- Klorman, R., Weerts, T. C., Hastings, J. E., Melamed, B. G., and Lang, P. J., 1974. Psychometric descriptions of some specific fear questionnaires, Behavior Therapy 5 (1974), pp. 401–409.
- Koster, E. H. W., Crombez, G., Verschuere, B., and De Houwer, J., 2004. Selective attention to threat in the dot probe paradigm: Differentiating vigilance and difficulty to disengage, Behaviour Research and Therapy 42 (2004), pp. 1183–1192.
- Lipp, O. V., Derakshan, N., Waters, A. M., and Logies, S., 2004. Snakes and cats in the flowerbed: Fast detection is not specific to pictures of fear-relevant animals, Emotion 4 (2004), pp. 233–250.
- Mineka, S., and Öhman, A., 2002. Phobias and preparedness: The selective, automatic, and encapsulated nature of fear, Biological Psychiatry 52 (2002), pp. 927–937.
- Mogg, K., and Bradley, B. P., 1998. A cognitive-motivational analysis of anxiety, Behaviour Research and Therapy 36 (1998), pp. 809–848.
- Moradi, A. R., Taghavi, M. R., Neshat-Doost, H. T., Yule, W., and Dalgleish, T., 1999. Performance of children and adolescents with PTSD on the Stroop colour-naming task, Psychological Medicine 29 (1999), pp. 415–419.
- Nauta, M. H., Scholing, A., Rapee, R., Abbott, M., Spence, S. H., and Waters, A. M., 2004. A parent-report measure of children's anxiety: Psychometric properties and comparison with child-report in a clinic and normal sample, Behaviour Research and Therapy 42 (2004), pp. 813–839.
- Öhman, A., 1993. "Fear and anxiety as emotional phenomena: Clinical phenomenology, evolutionary perspectives, and information processing mechanisms". In: Lewis, M., and Haviland, J. M., eds. Handbook of emotions. New York: Guilford; 1993. pp. 511–536.
- Öhman, A., Flykt, A., and Esteves, F., 2001. Emotion drives attention: Detecting the snake in the grass, Journal of Experimental Psychology: General 130 (2001), pp. 466–478.
- Öhman, A., Lundqvist, D., and Esteves, F., 2001. The face in the crowd revisited: A threat advantage with schematic stimuli, Journal of Personality and Social Psychology 80 (2001), pp. 381–396.
- Öhman, A., and Mineka, S., 2001. Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning, Psychological Review 108 (2001), pp. 483–522.
- Pashler, H., 1987. Detecting conjunctions of color and form: Reassessing the serial search hypothesis, Perception and Psychophysics 41 (1987), pp. 191–201.
- Posner, M. I., and Petersen, S. E., 1990. The attention system of the human brain, Annual Review of Neuroscience 13 (1990), pp. 25–42.
- Purcell, D. G., Stewart, A. L., and Skov, R. B., 1996. It takes a confounded face to pop out of a crowd, Perception 25 (1996), pp. 1091–1108.
- Rohlf, F. J., and Sokal, R., 1981. Statistical tables. San Francisco: Freeman; 1981.
- Spence, S. H., 1998. A measure of anxiety symptoms among children, Behaviour Research and Therapy 36 (1998), pp. 545–566.
- Taghavi, M. R., Dalgleish, T., Moradi, A. R., Neshat-Doost, H. T., and Yule, W., 2003. Selective processing of negative emotional information in children and adolescence with generalized anxiety disorder, British Journal of Clinical Psychology 42 (2003), pp. 221–230.
- Taylor, M. J., and Khan, S. C., 2000. Top-down modulation of early selective attention processes in children, International Journal of Psychophysiology 37 (2000), pp. 135–147.
- Vasey, M. W., Daleiden, E. L., Williams, L. L., and Brown, L. M., 1995. Biased attention in childhood anxiety disorders: A preliminary study, Journal of Abnormal Child Psychology 23 (1995), pp. 267–279.
- Vasey, M. W., and MacLeod, C., 2001. "Information-processing factors in childhood anxiety: A review and developmental perspective". In: Vasey, M. W., and Dadds, M. R., eds. The developmental psychopathology of anxiety. New York: Oxford University Press; 2001. pp. 27–42.
- Waters, A. M., Lipp, O. V., and Spence, S. H., 2004. Attentional bias toward fear-related stimuli: An investigation with non-selected children and adults and children with anxiety disorders, Journal of Experimental Child Psychology 89 (2004), pp. 320–337.
- Waters, A. M., Lipp, O. V., and Spence, S. H., 2005. The effects of affective picture stimuli on blink modulation in adults and children, Biological Psychology 68 (2005), pp. 257–281.
- Williams, J. M. G., Watts, F. N., MacLeod, C., and Mathews, A. M., 1997. Cognitive psychology and emotional disorders. Chichester, UK: Wiley; 1997.
- Wolfe, J. M., 1998. "Visual search". In: Pashler, H., ed. Attention. Hove, England: Psychology Press; 1998. pp. 13–73.
- Yiend, J., and Mathews, A., 2001. Anxiety and attention to threatening pictures, Quarterly Journal of Experimental Psychology 54A (2001), pp. 665–681.