Abstract
Capsule Use of Light Detection and Ranging (LiDAR) data identified suitable Willow Warbler habitat based on mean vegetation height. This habitat model provided maps of distribution and occupation of suitable habitat.
Aims To identify habitat associations in woods with different vegetation structure and management systems during a period of low Willow Warbler populations.
Methods Locations of all Willow Warblers were mapped during the breeding season in three woods of contrasting management; recent low intervention, actively coppiced woodland and high forest with clear‐fells. Height profile models of each wood were derived from airborne LiDAR. The mean vegetation height at locations with Willow Warblers and a sample from the rest of the wood were used to produce models of optimum habitat and breadth of habitat occupied in each wood. The habitat model was then used to produce maps of suitable habitat.
Results The habitat models did not differ between woods, with highest probability of Willow Warbler occurrence in mean vegetation heights of 3.7–5.3 m. Habitat of heights 6–11 m appeared less suitable, being only partly occupied. Habitat maps showed that habitat of suitable height was only occupied when it occurred as large patches; smaller patches (mostly <0.5 ha) and edges along rides and fields were not used.
Conclusion The use of LiDAR derived measures of vegetation height identified areas of suitable habitat for Willow Warblers. Willow Warblers occupied areas of low mean vegetation height either as early successional or open canopy woodland in all woods. Height‐based habitat maps can identify areas of suitable habitat within larger expanses of heterogeneous woodland and are a potentially useful tool in assessing changes in extent of what are often temporary patches of habitat.
INTRODUCTION
In the UK, the Willow Warbler Phylloscopus trochilus population is in long‐term decline making it a species of conservation concern (Gregory et al. Citation2002, Fuller, Noble, et al. Citation2005). However, there are large differences in population trends between different parts of the country; declines are concentrated in England and Wales, whereas in Scotland and Northern Ireland Willow Warblers have increased (Amar et al. Citation2006, Risely et al. Citation2008). A recent survey (Repeat Woodland Bird Survey) of changes in woodland bird populations over several decades found that declines were associated with habitat changes, being greatest at sites where canopy cover and tree basal area had increased and where low cover had decreased (Amar et al. Citation2006). Such changes are consistent with the effects of woodland maturation, lack of management and deer browsing (Morecroft et al. Citation2001, Fuller, Noble, et al. Citation2005). Thus, large‐scale changes in woodland habitat structure appear to be associated with the decline of Willow Warblers in the UK, though habitat change on the breeding grounds is not necessarily the primary driver of the declines (Fuller, Noble, et al. Citation2005).
In forests and woodland, vegetation structure is an important factor determining the diversity and composition of bird communities (MacArthur & MacArthur Citation1961, Willson Citation1974, Helle & Fuller Citation1988). Numerous studies have found that different species of woodland birds inhabit areas with distinct structural profiles (Fuller & Henderson Citation1992, Fuller & Green Citation1998, James Citation1971) and the habitat occupied by particular species is often tacitly assumed to have a similar structural profile throughout the species range. However, there can be shifts in the vegetation structure used by a species in different parts of its geographical range, for example, due to niche shifts in response to interspecific competition or to climate and resource differences in different locations (Cody Citation1978, Wilson et al. Citation2002, Fuller Citation2002).
Willow Warblers occur in a wide range of woodland types and have been described as birds of shrubby or early successional woodland, and woodland with sparse tree cover, often showing a preference for birch trees (Yapp Citation1962, Simms Citation1985, Cramp Citation1992). In northern latitudes Willow Warblers can occur in relatively mature birch woodland (Gillings et al. Citation1998). The few quantitative descriptions of Willow Warbler habitat published (Cody Citation1978, Sæther Citation1983, Fuller & Henderson Citation1992) have identified the importance of vegetation height and density, with Willow Warblers occurring in habitats with high densities of vegetation below 5 m and low densities of vegetation in taller trees. These studies have used multivariate analyses based on field recording of tree heights and of vegetation density in different height bands to define the habitat used.
In recent years, satellite remote sensing has been used to identify broad habitat associations (reviewed by Gottschalk et al. Citation2005) and airborne remote sensing is exploited increasingly to investigate animal–habitat relationships in more detail (Hinsley, Hill, Gaveau, et al. Citation2002, Hinsley, Hill, Bellamy, et al. Citation2006, Hinsley, Hill, Fuller, et al. in press, Bradbury et al. Citation2005). First and last return airborne Light Detection and Ranging (LiDAR), as used in this study (Gaveau & Hill Citation2003), is still uncommon in animal–habitat research, but has several advantages over other methods of habitat measurement (Vierling et al. Citation2008). It is able to capture habitat data at a spatial resolution of less than a metre, much finer than is possible with the more commonly used satellite reflectance data (Gottschalk et al. Citation2005) or full wave form LiDAR (Goetz et al. Citation2007). First and last return airborne LiDAR can characterize three‐dimensional habitat structure in great detail and across broad areas. This allows analyses of ecological processes across entire woodlands, forests and landscapes, such as spatially explicit associations between bird territories and the vertical structure of forest canopy (Broughton et al. Citation2006, Hinsley et al. Citation2006).
In this study, we used airborne LiDAR to identify and compare the habitat occupied by Willow Warblers in three woods with different vegetation structures and management regimes in lowland England after the main period of population decline.
METHODS
Study sites
The three study sites were all lowland mixed deciduous woodlands in eastern or central England; Monks Wood in Cambridgeshire (52°24′N, 0°14′W), Bradfield Woods in Suffolk (52°11′N, 0°50′E), and Sheephouse Wood in Buckinghamshire (51°54′N, 0°59′W).
Monks Wood included Monks Wood National Nature Reserve (NNR), 157 ha, and the adjacent grounds of the Centre for Ecology and Hydrology, 24 ha. The site is mainly mature Ash Fraxinus excelsior, Oak Quercus robur and Field Maple Acer campestre woodland (10–18 m high) with a hawthorn Crataegus spp., Hazel Corylus avellana and Blackthorn Prunus spinosa under‐storey (2–10 m high). It also includes some elm Ulmus sp. and scattered Silver Birch Betula pendula and Aspen Populus tremula, areas of scrub, young regenerating woodland, grass fields, small clearings and wide grassy rides with managed edges (Massey & Welch Citation1994, Hill & Thomson Citation2005). Most of the management in Monks Wood is concentrated along the rides and in the open glades and fields, the woodland has been left mostly to develop naturally since widespread clear‐felling in the 1920s (Steele & Welch Citation1973).
Bradfield Woods, 70 ha, is predominantly actively coppiced woodland with the trees cut on a rotation of approximately 25 years (Fuller & Henderson Citation1992, Hinsley et al. in press). The dominant coppiced tree species are Birch, Alder Alnus glutinosa, Hazel and Ash. There is a thin scattering of retained Oak, Ash and Birch standard trees. Because the wood is managed by coppicing, much of the canopy is relatively low (<10 m).
Sheephouse Wood, 59 ha, is predominantly mature Oak woodland managed for timber production, with a canopy height of 14–18 m over much of its area (Hinsley et al. in press). The under‐storey is mainly Hazel, Field Maple and hawthorn. Two areas of about 2 ha were clear‐felled and several smaller patches cut as group‐fells in the 1980s.
Bird census
In all three woods, bird locations were obtained by mapping bird activity during the breeding season. For Monks Wood, data were collected in 2004 by searching the whole wood for singing Willow Warblers on six occasions between mid‐April and mid‐May, with 3–7 days between visits. For Sheephouse Wood and Bradfield Woods bird data were collected in 2003 using territory mapping methods (Bibby et al. Citation2000). Birds were mapped on eight visits to Bradfield Woods and six visits to Sheephouse Wood between early April and early June. The average locational accuracy of mapping was estimated to be approximately 5–10 m, and depended on vegetation structure and proximity of the observer to the bird. For the analysis, no attempt was made to allocate registrations from successive visits to individual birds, pairs or territories. However, the approximate numbers of territories involved were 26 in Monks Wood, 20 in Bradfield Woods and 3 in Sheephouse Wood.
Habitat structure
Woodland structure variables were obtained from Digital Canopy Height Models (DCHMs) of the woods derived from airborne LiDAR (Optech ATLM) flown in June 2000 for Monks Wood and September 2003 for both Bradfield and Sheephouse Woods. The DCHMs mapped only the exposed ‘top canopy’ and did not include any height information from concealed shrub and ground layers. Full details of airborne LiDAR data acquisition and processing for Monks Wood are given in Gaveau & Hill (Citation2003) and Patenaude et al. (Citation2004). Data processing for Bradfield and Sheephouse woods followed similar methods to Monks Wood. However, the higher sampling density of the LiDAR data at Bradfield and Sheephouse (one laser hit per c. 1 m2) compared with Monks Wood (one laser hit per c. 5 m2) allowed for interpolation of a raster grid with finer spatial resolution (0.5 m pixel size compared with 1 m). A pilot analysis using Willow Warbler census data from 2000 for a 4 ha area of woodland contiguous with Monks Wood NNR identified two correlated vegetation structural variables which gave a good description of Willow Warbler habitat; the proportion of sample area (see later) with a top height between 1 m and 5 m, representing the exposed shrub layer, and mean vegetation height for the whole sample area. Of the two, mean vegetation height for the whole sample area was selected because it integrated information on the proportions of the sample area under tree canopy, shrub canopy and open ground with that of the heights of the trees and shrubs present. An analysis of the sensitivity of results to the sample area used showed little difference in model results for 5 m, 10 m and 15 m radius samples in the pilot data. Therefore, it appeared that habitat measures, as used in this study, were not unduly sensitive to sample area. A sample area of 15 m radius was chosen to be small enough to be specific to the bird location while also in the upper range of mapping accuracy, reducing model errors due to locational inaccuracy. Thus, for each bird registration, a circular sample area of 15 m radius (707 m2) centred on the bird was taken to represent the vegetation that the bird was using when recorded.
Habitat models
Binary logistic regression was used to define the shape of the relationship between Willow Warbler occurrence and vegetation structure using bird presence or absence in sample plots. Records of absence were created using a 150 m grid laid over each study wood and placing 15 m radius plots at each grid intersection. Any such plots that overlapped with a plot around a bird record were eliminated. The remaining plots provided a sample of vegetation representing bird absence. The 150 m sampling grid was chosen to provide a balanced sample of bird presence and absence for modelling the habitat in Monks Wood (the largest wood).
The pilot data identified quadratic logistic regression models (Gaussian logistic curve) (ter Braak & Looman Citation1995) as providing the best fit for the relationships between Willow Warbler occurrence and mean vegetation height. A quadratic model for optimal habitat makes ecological sense; Willow Warblers do not breed in fields without some woody growth, and infrequently in mature closed canopy woodland, but mainly occur in open woodland and scrub. A quadratic curve was the best fit for the Monks Wood data, but not for Sheephouse or Bradfield Woods. This was due to a little open ground in these two woods; in Sheephouse Wood no data points from the 150 m sampling grid were located in areas with mean vegetation of less than 1 m and in Bradfield Woods none were less than 3.6 m. Therefore, to facilitate comparison across all three woods, the models for Sheephouse and Bradfield Woods were transformed to quadratic curves by adding a single data point for bird absence at 0.1 m to both data sets.
Where the sample plot around a bird registration overlapped with a plot from an earlier census date it was excluded from the analysis to avoid duplication. The use of observed locations of birds, rather than inferred territories, avoided the problems of assigning territory boundaries, and particularly so if boundaries should vary over the breeding season (Bibby et al. Citation2000). Also, the use of registrations allowed data to be used from the exact locations used by individual birds.
The fit of the model to observed Willow Warbler presence/absence was assessed using Cohen’s kappa (Fielding & Bell Citation1997). Kappa gives an overall value of predictive ability using both the numbers of correctly predicted presences and absences, whilst also taking account of the number of samples used in the model. For each wood, correctly predicted presences were taken as those with a predicted probability of occurrence greater than the proportion of locations with birds present, i.e. 0.44, equivalent to an occupancy rate of 61 out of 137 locations for Monks Wood, for Bradfield Woods, 0.76 (71 out of 94) and for Sheephouse Wood, 0.24 (9 out of 38). Kappa has a value of between 0 and 1, where values below 0.4 are poor predictors; 0.4 to 0.6 are moderate predictors; and greater than 0.6 are good predictors (Manel et al. Citation2001). The value of kappa is relatively unaffected by differences in the proportion of occupied samples (prevalence) when using prevalence as the threshold for distinguishing between predicted presences and absences (Manel et al. Citation2001, Liu et al. Citation2005).
Comparison of habitat models between woods
Two important parameters of the Gaussian logistic curve are the ‘optimum’ and the ‘tolerance’. The optimum is the value of the predictor which gives the maximum probability of species occurrence and the tolerance is the spread of the curve about the optimum. The values and standard errors of the optimum and tolerance were estimated in genstat 6.1 for the models for each wood and differences between woods in optimum habitat and/or the range of vegetation occupied were compared.
Habitat maps
Although presence or absence of birds at point samples can give a good description of the habitat favoured by Willow Warblers, the spatial arrangement of suitable habitat may affect where Willow Warblers are found in addition to direct effects of habitat structure. By locating occupied/unoccupied sample points on a map of predicted suitable habitat the spatial context of habitat use and errors in associated models could be examined. After comparing habitat models, the model which best described Willow Warbler habitat was used to produce habitat maps for each wood within arcview 3.1. The mean canopy height was calculated for a 15 m radius circle around each pixel of the DCHM of each wood. Each pixel was then classified as suitable habitat, or shorter or taller than habitat using a probability of occurrence of >0.5 to define suitable habitat. Willow Warbler and sample grid locations were then marked on the resulting habitat maps to compare the distributions of Willow Warblers in relation to potential habitat suitability and distribution.
RESULTS
Habitat models
In all three woods, a Gaussian logistic curve using mean vegetation height as the predictive variable gave moderate or good predictions for the observed occurrences of Willow Warblers (Table , Fig. ). Overall, the models indicated optimum vegetation height for Willow Warblers to be in the range 3.7–5.3 m (see later). There were more predictive errors identifying suitable habitat which lacked Willow Warblers (28% of locations with predicted habitat had no birds in Monks Wood, 11% in Bradfield Woods, 18% in Sheephouse Wood) than for birds found in unsuitable habitat (birds were present in 7% of locations predicted to be unsuitable for Monks Wood, 30% for Bradfield Woods, 0% for Sheephouse Wood). In general, the models for all woods made fewest errors in predicting non‐habitat greater than 11 m (incorrect predictions: 4% for Monks Wood, 12% for Bradfield Woods and 0% for Sheephouse Wood) and for suitable habitat between 1 m and 6 m, (incorrect predictions: 17% for Monks Wood, 5% for Bradfield Woods and 11% for Sheephouse Wood) (Fig. ). Most errors occurred for heights either side of the 1–6 m band of greatest suitability (6–11 m, incorrect predictions: 36% for Monks Wood, 20% for Bradfield Woods and 17% for Sheephouse Wood; 0–1 m, incorrect predictions: Monks Wood only, 29%) (Fig. ).
Figure 1 Habitat used by Willow Warblers in three woods. Presence (probability = 1) and absence (probability = 0) of Willow Warblers in relation to mean vegetation height in 15 m radius sample circles for (a) Monks Wood; (b) Bradfield Woods; and (c) Sheephouse Wood. The line represents the logistic quadratic curve fitted to the data (Table ).
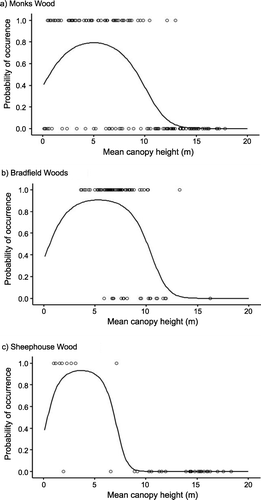
Figure 2 Distribution of mean heights for sample points in (a) Monks Wood; (b) Bradfield Woods; and (c) Sheephouse Wood. The number of locations in each height band is represented as the percentage of the total number of locations in the wood. Shaded bars indicate samples where Willow Warbler presence or absence was incorrectly predicted from the fitted quadratic logistic models (Table ).
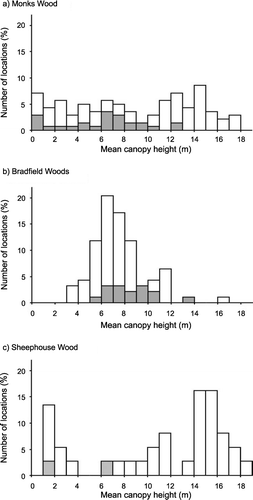
Table 1. Quadratic equations fitted to Willow Warbler presence/absence data and mean canopy height for three woods and measures of the goodness of fit of the models to the data.
Comparison of habitat models between woods
Despite the different ranges of habitat heights available in the different woods, there was no statistical difference in the modelled optimum height of habitat occupied by Willow Warblers (Bradfield Woods mean = 5.3 m, 95% CI = 3.1–8.4 m; Monks Wood 5.0 m, 4.0–6.0 m; Sheephouse Wood 3.7 m, 2.3–5.1 m; Table ). The range of vegetation heights predicted to be suitable for Willow Warblers was also similar in Monks Wood and Bradfield Woods, despite the lack of sample points in low vegetation from Bradfield Woods. Willow Warblers were predicted to be present with a probability of > 0.5 in vegetation with a mean height of between 0.6–9.3 m in Monks Wood and 0.6–10.1 m in Bradfield Woods. However, in Sheephouse Wood, the lack of vegetation in the middle range of canopy heights (only five sample points between 5 m and 10 m) excluded the taller end of Willow Warbler habitat used in the other two woods and led to a much narrower definition of Willow Warbler habitat. They were predicted to be present with a probability of > 0.5 in vegetation with a mean height of 0.4–6.8 m. The large standard errors of some of the model parameters resulted in no statistical difference at P < 0.05 being found for habitat tolerance between woods. However, the difference between Monks Wood and Sheephouse Wood which showed the greatest contrast was marginally significant at P < 0.10 (Table ).
Table 2. Comparison between woods of most preferred habitat height (optimum) and range of habitat heights (tolerance).
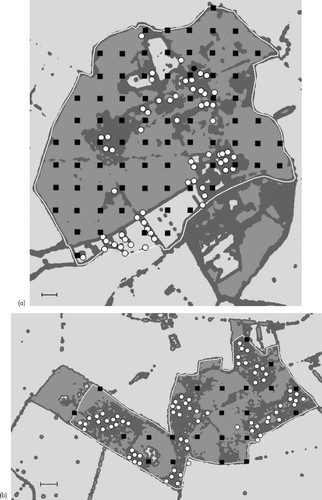
Monks Wood provided data representing the full range of habitat heights and gave good predictions of Willow Warbler habitat across all three woods (predicted fit of data from each wood to the Monks Wood model (kappa); Monks Wood = 0.62, Bradfield Woods = 0.58, Sheephouse Wood = 0.74). Therefore, the habitat model for Monks Wood was used to produce habitat maps for all three woods with a mean height of between 0.6 and 9.3 m as suitable habitat (Fig. ).
Figure 3 Maps showing the distribution of suitable habitat in (a) Monks Wood; (b) Bradfield Woods; and (c) Sheephouse Wood. Dark grey areas are suitable habitat between 0.6 m and 9.3 m, medium grey areas are unsuitable habitat taller than 9.3 m, pale grey areas are unsuitable habitat shorter than 0.6 m. The maps are based on the mean height of a 15 m radius around each pixel of the Digital Canopy Height Model for each wood. Habitat categories are based on the Monks Wood habitat model (Table ). Each study area is outlined in white and the locations of birds used in the habitat models are shown by white dots, locations of grid sample absences are shown by black squares. The scale bar on each map represents 100 m.

Habitat maps
From these habitat maps, 12 birds (8.5% of records) occurred in vegetation taller than that predicted to be suitable habitat, 6 each in Bradfield Woods and Monks Wood. The median distance of these birds from patches of suitable habitat was 6 m. Only one, at 29.8 m, was greater than 20 m from a predicted habitat patch.
The distribution of suitable habitat in all three woods consisted of several large patches connected by linear strips of habitat along ride and field edges. There was also a scattering of small patches of habitat, some of which were isolated in fields or in areas of taller woodland. Others were connected to other small or large patches. All of the Willow Warblers were located in the larger patches of habitat; none were located in small patches or in habitat occurring as thin linear edges. Although all of the larger areas of suitable habitat had Willow Warblers in them, birds were not distributed evenly through these habitat patches. In some, a large proportion of the habitat had no records of Willow Warblers.
DISCUSSION
The optimum vegetation height identified in this study is consistent with the results of two previous quantitative models of Willow Warbler habitat, using field estimates of vegetation height and density (Cody Citation1978, Sæther Citation1983). Both these studies identified Willow Warbler habitat as having the highest density of vegetation below 7 m and very low densities of taller vegetation. Also, in an upland birch wood, there was a strong positive correlation of Willow Warbler numbers with the percentage cover of birch at 1–4 m (Gillings et al. Citation1998). However, the results produced in our current study using remote sensed estimates of canopy height gave a more robust and simpler habitat model than the earlier attempts (Cohen’s kappa: current study, 0.56–0.80 [Table ]; N. England, 0.53 [Cody Citation1978]; S. Sweden, 0.52 [Cody Citation1978]; C. Norway, 0.59 [Sæther Citation1983]). The previous studies were carried out prior to the recent population declines recorded in England and both noted the wide range of habitats used by Willow Warblers compared with the other warblers studied, as did Tiainen et al. (Citation1983). In these studies, Willow Warblers were occasionally recorded in vegetation up to 15–20 m, but it was also the only species which showed a strong preference for open habitats and early seral stages. Therefore, the more specific and restricted habitat found in English woods in this study may be a reflection of the species occupying predominantly the most suitable habitat when population size is relatively small.
The good agreement in the optimum height for Willow Warbler habitat between woods of different height profiles, particularly for Monks Wood and Bradfield Woods, indicates a strong habitat preference for a mean vegetation height of about 5 m. Most of the errors occurred in heights on the edges of the predicted habitat range and may represent marginal habitat where other factors, such as finer‐scale structural details or tree species composition, may contribute to habitat suitability. Willow Warblers are known to choose habitats with good insect availability such as wet willow scrub, young woodland with dense young growth and tree species with pest outbreaks (Cramp Citation1992, Hogstad Citation2005). In particular, it has been suggested that Willow Warblers show a preference for birch trees (Yapp Citation1962, Mason Citation1997). However, such a preference may be compounded with structural differences because birch is often a pioneer species most frequent in open areas of woodland. The association between Willow Warblers and birch has been suggested as a reason for differences in occupation of otherwise similar habitats (Mason Citation1997). Developments in the use of aerial photography and multispectral reflectance data in the identification of tree species (Zhang et al. Citation2004) may add to our ability to identify woodland bird habitats using remote sensing techniques.
Although mean canopy height provided a simple measure of habitat suitability, the configuration of habitat patches was also important in whether or not habitat was used. The habitat maps showed that narrow strips of habitat on external edges of woodland, along rides and around clearings were not used. Other studies have recorded lower densities of Willow Warblers near roads (Kuitenen et al. Citation1998) and on woodland edges (Mason Citation2001). Most of the small patches of habitat not occupied were less than 0.5 ha, which, although within the range of territory sizes recorded for Willow Warblers, would be unlikely to contain more than one pair (Cramp Citation1992). Mean territory sizes from a range of studies were mostly between 0.2 and 1.2 ha, although small territories of <0.1 ha have also been recorded (Cramp Citation1992). The reasons for not using edges and small patches are not understood; there may be differences in habitat perception, inter‐specific competition and predation risk or conspecific attraction may attract birds preferentially to larger patches. There has been little attention paid to defining the size and shape of suitable habitat patches within heterogeneous woodland. Some of the problems stem from the difficulty in identifying and delimiting habitat patches for bird species in extensive areas of heterogeneous habitat, for example, distinguishing areas that are actively used from those simply traversed. The ability of remote sensed data to quantify vegetation structure at high resolution offers the potential to develop simple algorithms for identifying species‐specific use of heterogeneous habitat mosaics.
Most of the birds that occurred in habitat categorized as unsuitable were individuals in taller trees and all but one of these were very close to suitable habitat. These may be due to errors in the accuracy of recording locations in the field or may have been birds using tall trees on the edges of their territories as song posts. Unoccupied samples of apparently suitable habitat, as well as uneven distribution of Willow Warblers across suitable habitat patches on the habitat maps, may have been due to a lack of habitat saturation, i.e. suitable habitat may remain vacant if populations are too small to occupy all available habitat. The current study was undertaken after the main decline of Willow Warbler populations in southern England creating the potential for unoccupied habitat (Eaton et al. Citation2004). When populations increase, individuals may occupy marginal or even unsuitable habitat which may differ in structure from ideal breeding habitat (Hinsley et al. Citation1996; Fernández‐Juricic Citation2001). Stability of occupation may be a strong indicator of habitat quality. However, for species occupying early successional stages, the transient and shifting nature of such habitat would have to be taken into account.
Thus, habitat quality and/or occupation can potentially vary over time, and with other factors including climate (Hinsley et al. Citation2006) and/or large‐scale range changes (Wilson et al. Citation2002). There is also an increasing awareness of the influence of landscape structure on populations and how habitat selection may take place in a hierarchical process at different spatial scales (Bennett et al. Citation2004; George & Zack Citation2001; De Graaf et al. Citation1998). Woodland size and surrounding landscape configuration can influence suitability of woods for Willow Warblers (Hinsley et al. Citation1995; Bellamy et al. Citation2003). Combining detailed structural information at the wood scale (Hill et al. Citation2004) with landscape scale information on woodland cover and configuration (Fuller, Devereaux, et al. Citation2005) could lead to a greater understanding of habitat selection and species distributions.
The use of LiDAR‐derived models was highly successful in characterizing woodland structure at a resolution and spatial extent that allowed the detection of an ecologically‐significant habitat association. Investigations of this kind have previously been restricted using field‐based data, which provide data of limited spatial extent and poor vertical characterization (Vierling et al. Citation2008). In countries such as the UK, where the employment of skilled fieldworkers to conduct field‐based surveys is relatively expensive, LiDAR can also be a cost‐effective method of gathering data on a landscape scale. The costs of hardware (including aircraft) and data processing will still be limiting in many circumstances, although increasing commercial availability of LiDAR data will reduce this with time. Use of LiDAR has advantages over labour‐intensive field‐based sampling in spatially extensive and strongly three‐dimensional habitats such as forests (see review in Vierling et al. [Citation2008]). The development of further applications in airborne remote sensing will also increase its potential value for ecologists. For example, It is now possible to generate three‐dimensional models of the forest under‐storey using LiDAR (Hill & Broughton Citation2009).
Conclusions
Based on Willow Warbler presence/absence in the woods studied, the most frequently used habitat occurred within a height range of 1–6 m, with more marginal habitat in the additional ranges of <1 m and 6–11 m. The habitat occupied had a consistent height profile despite different management regimes and vegetation structure. These woods were all lowland deciduous woods with similar topography, soil and climate. It would be of interest to explore the height profiles of Willow Warbler habitat in other parts of the species range, for example, in the more open canopy woodlands in Scotland where Willow Warblers have increased in numbers in contrast to the population declines in southern England.
ACKNOWLEDGEMENTS
English Nature kindly gave permission for work in Monks Wood NNR, the Claydon Estate for Sheephouse Wood and the Suffolk Wildlife Trust for Bradfield Woods NNR. We are grateful to the Environment Agency of England and Wales for funding and acquiring the LiDAR data for Monks Wood. The LiDAR data for Sheephouse and Bradfield Woods were acquired by the NERC ASRF in conjunction with the Unit for Landscape Modelling at the University of Cambridge. This work was part‐funded by the Natural Environment Research Council (NERC) Earth Observation Programme.
REFERENCES
- Amar , A. , Hewson , C.M. , Thewlis , R.M. , Smith , K.W. , Fuller , R.J. , Lindsell , J.A. , Conway , G. , Butler , S. and MacDonald , M.A. 2006 . What’s Happening to Our Woodland Birds? Long‐Term Changes in the Populations of Woodland Birds , Thetford : BTO . BTO Research Report 169
- Bellamy , P.E. , Rothery , P. and Hinsley , S.A. 2003 . Synchrony of woodland bird populations: the effect of landscape structure . Ecography , 26 : 338 – 348 .
- Bennett , A.F. , Hinsley , S.A. , Bellamy , P.E. , Swetnam , R.D. and MacNally , R. 2004 . Do regional gradients in land‐use influence richness, composition and turnover of bird assemblages in small woods? . Biol. Con. , 119 : 191 – 206 .
- Bibby , C.J. , Burgess , N.D. , Hill , D.A. and Mustoe , S. 2000 . Bird census techniques, , 2nd edition , London : Academic Press .
- Bradbury , R.B. , Hill , R.A. , Mason , D.C. , Hinsley , S.A. , Wilson , J.D. , Balzter , H. , Anderson , Q.A. , Whittingham , M.J. , Davenport , I.J. and Bellamy , P.E. 2005 . Modelling relationships between birds and vegetation structure using airborne LiDAR data: a review with case studies from agricultural and woodland environments . Ibis , 147 : 443 – 452 .
- Broughton , R.K. , Hinsley , S.A. , Bellamy , P.E. , Hill , R.A. and Rothery , P. 2006 . Marsh Tit Poecile palustris territories in a British broad‐leaved wood . Ibis , 148 : 744 – 752 .
- Cody , M.L. 1978 . Habitat selection and interspecific territoriality among the Sylviid warblers of England and Sweden . Ecol. Monogr. , 48 : 351 – 396 .
- Cramp , S. , ed. 1992 . Birds of the Western Palearctic , Vol. 6 , Oxford : Oxford University Press .
- DeGraaf , R.M. , Hestbeck , J.B. and Yamasaki , M. 1998 . Associations between breeding bird abundance and stand structure in the White Mountains, New Hampshire and Maine, USA . Forest Ecol. Manag. , 103 : 217 – 233 .
- Eaton , M.A. , Noble , S.G. , Cranswick , P.A. , Carter , N. , Wotton , S. , Ratcliffe , N. , Wilson , A. , Hilton , G.M. and Gregory , R.D. 2004 . The State of the UK’s Birds 2003 , Sandy : BTO, RSPB, WWT .
- Fernández‐Juricic , E. 2001 . Density‐dependent habitat selection of corridors in a fragmented landscape . Ibis , 143 : 278 – 287 .
- Fielding , A.H. and Bell , J.F. 1997 . A review of methods for the assessment of prediction errors in conservation presence/absence models . Environ. Con. , 24 : 38 – 49 .
- Fuller , R.J. . Spatial differences in habitat selection and occupancy by woodland bird species in Europe: a neglected aspect of bird–habitat relationships . Proceedings of the 11th Annual IALE(UK) Conference . Avian Landscape Ecology: Pure and Applied Issues in the Large‐scale Ecology of Birds , Edited by: Chamberlain , D. and Wilson , A. pp. 101 – 111 . IALE .
- Fuller , R.J. and Green , G.H. 1998 . Effects of woodland structure on breeding bird populations in stands of coppiced lime (Tilia cordata) in western England over a 10‐year period . Forestry , 71 : 199 – 218 .
- Fuller , R.J. and Henderson , A.C.B. 1992 . Distribution of breeding songbirds in Bradfield Woods, Suffolk, in relation to vegetation and coppice management . Bird Study , 39 : 73 – 88 .
- Fuller , R.J. , Noble , D.G. , Smith , K.W. and Vanhinsbergh , D. 2005 . Recent declines in populations of woodland birds in Britain: a review of possible causes . Br. Birds , 98 : 116 – 143 .
- Fuller , R.M. , Devereaux , B.J. , Gillings , S. , Amable , G.S. and Hill , R.A. 2005 . Indices of bird‐habitat preference from field surveys and remote sensing of land cover: a study of south‐eastern England with wider implications for conservation and biodiversity assessment . Global Ecol. Biogeog. , 14 : 223 – 239 .
- Gaveau , D.L.A. and Hill , R.A. 2003 . Quantifying canopy height underestimation by laser pulse penetration in small‐footprint airborne laser scanning data . Can. J. Remote Sensing , 29 : 650 – 657 .
- George , T.L. and Zack , S. 2001 . Spatial and temporal considerations in restoring habitat for wildlife . Restoration Ecol. , 9 : 272 – 279 .
- Gillings , S. , Fuller , R.J. and Henderson , A.C.B. 1998 . Avian community composition and patterns of bird distribution within birch‐heath mosaics in north‐east Scotland . Ornis Fenn. , 75 : 27 – 37 .
- Goetz , S. , Steinberg , D. , Dubayah , R. and Blair , B. 2007 . Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA . Remote Sensing Environ. , 108 : 254 – 263 .
- Gottschalk , T.K. , Huettmann , F. and Ehlers , M. 2005 . Thirty years of analysing and modelling avian habitat relationships using satellite imagery data: a review . Int. J. Remote Sensing , 26 : 2631 – 2656 .
- Gregory , R.D. , Wilkinson , N.I. , Noble , D.G. , Robinson , J.A. , Brown , A.F. , Hughes , J. , Procter , D. , Gibbons , D.W. and Galbraith , C.A. 2002 . The population status of birds in the United Kingdom, Channel Islands and Isle of Man: an analysis of conservation concern 2002–2007 . Br. Birds , 95 : 410 – 448 .
- Helle , P. and Fuller , R.J. 1988 . Migrant passerine birds in European forest successions in relation to vegetation height and geographical position . J. Anim. Ecol. , 57 : 565 – 579 .
- Hill , R.A. and Broughton , R.K. 2009 . Mapping understorey of deciduous woodland from leaf‐on and leaf‐off airborne LiDAR data: a case study in lowland Britain . ISPRS J. Photogramm. Remote Sensing. , 64 : 223 – 233 .
- Hill , R.A. , Hinsley , S.A. , Gaveau , D.L.A. and Bellamy , P.E. 2004 . Predicting habitat quality for Great Tits (Parus major) with airborne laser scanning data . Int. J. Remote Sensing , 25 : 4851 – 4855 .
- Hill , R.A. and Thomson , A.G. 2005 . Mapping woodland species composition and structure using airborne spectral and LiDAR data . Int. J. Remote Sensing , 17 : 3763 – 3779 .
- Hinsley , S.A. , Bellamy , P.E. , Newton , I. and Sparks , T.H. 1995 . Habitat and landscape factors influencing the presence of individual breeding bird species in woodland fragments . J. Avian Biol. , 26 : 94 – 104 .
- Hinsley , S.A. , Bellamy , P.E. , Newton , I. and Sparks , T.H. 1996 . Influences of population size and woodland area on bird species distributions in small woods . Oecologia , 105 : 100 – 106 .
- Hinsley , S.A. , Hill , R.A. , Bellamy , P. E. and Balzter , H. 2006 . The application of LiDAR in woodland bird ecology: climate, canopy structure and habitat quality . Photogramm. Eng. Remote Sensing , 72 : 1399 – 1406 .
- Hinsley , S.A. , Hill , R.A. , Fuller , R.J. , Bellamy , P.E. and Rothery , P. in press . Bird species distributions across woodland canopy structure gradients . Community Ecol. ,
- Hinsley , S.A. , Hill , R.A. , Gaveau , D.L.A. and Bellamy , P.E. 2002 . Quantifying woodland structure and habitat quality for birds using airborne laser scanning . Funct. Ecol. , 16 : 851 – 857 .
- Hogstad , O. 2005 . Numerical and functional responses of breeding passerine species to mass occurrence of geometrid caterpillars in a subalpine birch forest: a 30‐year study . Ibis , 147 : 77 – 91 .
- James , F.C. 1971 . Ordinations of habitat relationships among breeding birds . Wilson Bull. , 83 : 215 – 236 .
- Kuitenen , M. , Rossi , E. and Stenroos , A. 1998 . Do highways influence density of land birds? . Environ. Manag. , 22 : 297 – 302 .
- Liu , C. , Berry , P.M. , Dawson , T.P. and Pearson , R.G. 2005 . Selecting thresholds of occurrence in the prediction of species distributions . Ecography , 28 : 385 – 393 .
- MacArthur , R.H. and MacArthur , J.W. 1961 . On bird species diversity . Ecology , 42 : 594 – 598 .
- Manel , S. , Williams , H.C. and Ormerod , S.J. 2001 . Evaluating presence–absence models in ecology: the need to account for prevalence . J. Appl. Ecol. , 38 : 921 – 931 .
- Mason , C.F. 1997 . Association between Willow Warbler Phylloscopus trochilus territories and birch in woodlands in southeastern England . Ibis , 139 : 411 – 412 .
- Mason , C.F. 2001 . Woodland area, species turnover and the conservation of bird assemblages in lowland England . Biodiv. Cons. , 10 : 495 – 510 .
- Massey , M.E. and Welch , R.C. , eds. 1994 . Monks Wood National Nature Reserve: the Experience of 40 years 1953–93 , Peterborough : English Nature .
- Morecroft , M.D. , Taylor , M.E. , Ellwood , S.A. and Quinn , S.A. 2001 . Impacts of deer herbivory on ground vegetation at Wytham Woods, central England . Forestry , 74 : 251 – 257 .
- Patenaude , G. , Hill , R.A. , Milne , R. , Gaveau , D.L.A. , Briggs , B.B.J. and Dawson , T.P. 2004 . Quantifying forest above ground carbon content using LiDAR remote sensing . Remote Sensing Environ. , 93 : 368 – 380 .
- Risely , K. , Noble , D.G. and Baillie , S.R. 2008 . The Breeding Bird Survey 2007 , Thetford : BTO . BTO Research Report 508
- Sæther , B.E. 1983 . Habitat selection, foraging niches and horizontal spacing of Willow Warbler Phylloscopus trochilus and Chiffchaff P. collybita in an area of sympatry . Ibis , 125 : 24 – 32 .
- Simms , E. 1985 . British Warblers , London : New Naturalist, Collins .
- Steele , R.C. and Welch , R.C. , eds. 1973 . Monks Wood, a Nature Reserve Record , Huntingdon : Natural Environment Research Council .
- ter Braak , C.J.F. and Looman , C.W.N. 1995 . “ Regression ” . In Data Analysis in Community and Landscape Ecology , Edited by: Jongman , R.G.H. , ter Braak , C.J.F. and van Tongeren , O.F.R. Cambridge : Cambridge University Press .
- Tiainen , J. , Vickholm , M. , Pakkala , T. , Piiroinen , J. and Virolainen , E. 1983 . The habitat and spatial relations of breeding Phylloscopus warblers and the goldcrest Regulus regulus in southern Finland . Ann. Zool. Fenn. , 20 : 1 – 12 .
- Vierling , K.T. , Vierling , L.A. , Gould , W.A. , Martinuzzi , S. and Clawges , R.M. 2008 . LiDAR: shedding new light on habitat characterization and modelling . Front. Ecol. Environ. , 6 : 90 – 98 .
- Willson , M.F. 1974 . Avian community organization and habitat structure . Ecology , 55 : 1017 – 1029 .
- Wilson , A.M. , Henderson , A.C.B. and Fuller , R.J. 2002 . Status of the Nightingale Luscinia megarhynchos in Britain at the end of the 20th century with particular reference to climate change . Bird Study , 49 : 193 – 204 .
- Yapp , W.B. 1962 . Birds and Woods , Oxford : Oxford University Press .
- Zhang , Q.C. , Franklin , S.E. and Wulder , M.A. 2004 . Geostatistical and texture analysis of airborne‐acquired images used in forest classification . Int. J. Remote Sensing , 25 : 859 – 865 .