Abstract
Capsule Golden and White‐tailed Eagles selected different habitats for nesting.
Aim To investigate differences in nesting habitat used by sympatrically breeding eagles in western Scotland, following reintroduction of White‐tailed Eagles from 1975 onwards.
Methods Nest‐site locations from national surveys in 2003–05 were entered into a geographical information system (GIS) in order to measure a set of geographic parameters for each nest site. Binary logistic regression with backwards deletion of non‐significant terms was used to derive minimum adequate models at two spatial scales of the likelihood of an eagle nest belonging to one species or the other. We compared changes in occupancy between 1992 and 2003 of Golden Eagle territories inside and outside a GIS model of potential White‐tailed Eagle habitat and according to proximity to White‐tailed Eagle nests.
Results White‐tailed Eagles nested at lower altitudes than Golden Eagles, in more wooded habitats with more open water close by, tending to nest in trees where these were present. There were 3359 km2 of potential White‐tailed Eagle nesting habitat within 25 km of existing White‐tailed Eagle nests, containing 54 Golden Eagle territory centres, but we found no difference in change of occupancy for Golden Eagle territories close to White‐tailed Eagles compared with those further away.
Conclusion White‐tailed and Golden Eagles appear to partition nesting habitat in the west of Scotland by altitude. This corresponds with behaviour in western Norway and with the situation described in historical accounts of nest‐sites in western Scotland prior to extinction of White‐tailed Eagles. It is also consistent with recent studies showing little overlap in breeding season diet of Golden and White‐tailed Eagles in western Scotland, and likely partitioning of foraging habitat by altitude. We conclude that the likelihood of competitive exclusion is less than previously suggested.
INTRODUCTION
Lack (Citation1946) suggested that raptors (birds of prey) may be particularly suitable subjects for examining effects of interspecific competition, but it is only in recent years that this research field has attracted much interest. Studies have examined interspecific niche relationships within guilds (Jaksić & Braker Citation1983, Poole & Bromley Citation1988, Hakkarainen & Korpomäki Citation1996, Aumann Citation2001, Krüger et al. Citation2002) and between morphologically similar species (Bechard et al. Citation1990, Gerstell & Bednarz Citation1999, Burton & Olsen Citation2000, Katzner et al. Citation2003, Vrezec & Tome Citation2004, Carrete et al. Citation2005) with mixed results on evidence for competitive effects. Results that are more consistent have been found in studies that have examined the influence of large raptors on smaller raptors, including intra‐guild predation. For example, Golden Eagles Aquila chrysaetos affect the distribution and abundance of several smaller raptors (Poole & Bromley Citation1988, Ratcliffe Citation1993, Gainzarain et al. Citation2000, Fielding et al. Citation2003, Sergio et al. Citation2004), as do Eagle Owls Bubo bubo (Gainzarain et al. Citation2002, Sergio et al. Citation2003) and Northern Goshawks Accipiter gentilis (Kostrzewa Citation1991, Selås Citation1996, Krüger Citation2002, Petty et al. Citation2003).
In 1975, White‐tailed Eagles Haliaeetus albicilla were reintroduced into Scotland using nestlings from Norway (Love Citation1983, Evans et al. Citation2003, Bainbridge et al. Citation2003). Currently the population is over 40 breeding pairs and in a phase of marked expansion (Evans et al. Citation2009). Although this reintroduction pre‐dated the adoption of international guidelines for conservation translocations (IUCN Citation1987, Citation1998), good practice now requires pre‐release assessment of effects of the released species on the receiving ecosystem and post‐release study of long‐term processes of adaptation of the population of the released species. It has been suggested that the continued spread of White‐tailed Eagles in Scotland might lead to displacement of Golden Eagles through competitive effects as coastal ranges are reclaimed (Watson et al. Citation1992, Watson Citation1997, Halley & Gjershaug Citation1998, Halley Citation1998). On the Isle of Mull, however, where both species occur at relatively high density there was little evidence of competitive effects (Whitfield et al. Citation2002). Other work has recently challenged the basis for earlier predictions of competitive effects, such as dietary overlap (Madders & Marquiss Citation2003), purported historical evidence for competition and a greater digestive efficiency of White‐tailed Eagles (Whitfield et al. Citation2002). Both species may occupy the nest‐sites of the other (reviewed by Whitfield et al. [Citation2002]), and Fielding et al. (Citation2003) characterised the relationship between the two species in Scotland as one of ‘armed neutrality’, in that while both species may anecdotally kill each other, this did not appear to have a predatory basis.
Comparisons of the ‘food niches’ of White‐tailed and Golden Eagles in Scotland have tended to concentrate on examining the extent of dietary overlap, with no overt reference to spatial partitioning (Watson et al. Citation1992; although see Madders & Marquiss Citation2003). In other words, a simple assumption has been made that a high dietary overlap implies a high level of competition. This is probably too simplistic (Wiens Citation1989) and even when apparent resource use is very similar, competition may be minimal (Katzner et al. Citation2003). Whitfield et al. (Citation2002) suggested that competitive effects owing to dietary overlap between White‐tailed and Golden Eagles in Scotland would be mediated if the common dietary components were obtained in different locations. In western Norway, Golden Eagles tend to be found more often at higher elevation cliff nest‐sites, whereas White‐tailed Eagles are more closely associated with coastal locations and tree sites (Halley Citation1998). However, both species will use both types of site (Watson Citation1997, Willgohs Citation1961). Elsewhere in Europe, White‐tailed Eagles tend to inhabit lowland areas, whereas the Golden Eagle is characteristically, but not exclusively, an upland species (Helander & Stjernberg Citation2003, McGrady Citation1997). Although this appears also to have been the case historically in Scotland (Love Citation1983), no interspecific comparison of nest‐site use has been undertaken since White‐tailed Eagles were reintroduced. In this paper, we analyse features of nest‐site use in sympatric Scottish populations of White‐tailed and Golden Eagles.
We therefore examined three aspects of potential competition between White‐tailed and Golden Eagles. First, we hypothesised that the two species select different nest‐sites, and specifically that White‐tailed Eagles nest at lower altitudes, closer to water, and in trees more than Golden Eagles. Second, we used nearest neighbour distances to test whether Golden Eagles are more or less tolerant of White‐tailed Eagles than conspecifics. Third, we tested whether changes in the occupancy of Golden Eagle territories were correlated with proximity to White‐tailed Eagles.
METHODS
Study area
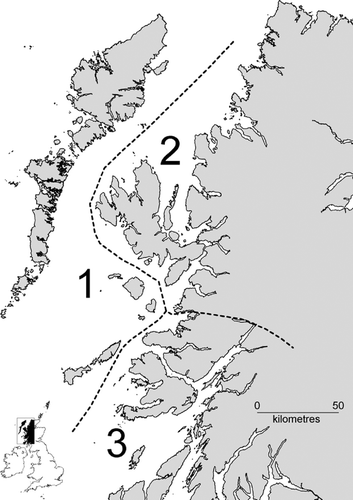
The study area was dictated by the breeding distribution of White‐tailed Eagles in Scotland and was delineated by a 25 km buffer around all known White‐tailed Eagle nest‐sites up to 2005. This buffer was the smallest to result in a single contiguous polygon within a geographical information system (GIS), mapinfo, populated with all nest locations. Within this broad area, we identified three regions with similar sized populations of both eagle species, but differing geographical characteristics: (1) Western and Small Isles; (2) Skye and Ross‐shire; and (3) Argyll (Fig. ). The proportion of woodland cover was particularly variable, at 0.5%, 6.3% and 18.4% of land area, respectively. Boundaries of regions were entered as a layer in the GIS.
Figure 1 The study area in western Scotland divided into three regions. (1) Western and Small Isles (2868 km2); (2) Skye/Ross (4783 km2); (3) Argyll (3625 km2).
Data collection and handling
All eagle nest‐site locations up to 2005 were recorded as national grid references to the nearest 100 m resolution and classed as either ‘tree’ or ‘cliff’ sites (Dennis et al. Citation1984, Green Citation1996, Eaton et al. Citation2007; Endnote Footnotea) with nests in trees growing on cliffs classified as ‘tree’. In all but three territories, all alternative nest‐sites were either ‘tree’ or ‘cliff’. All three territories with both types of site included only one nest of the minority class. Therefore, a further classification of territories as ‘tree’ or ‘cliff’ was reasonable. For analysis, we took the Golden Eagle locations recorded in the 2003 national survey (Eaton et al. Citation2007), and one location from each White‐tailed Eagle territory surveyed between 2003 and 2005 (the whole Scottish breeding population). Golden Eagle nest locations were not available for the Isle of Skye in 2003. Golden Eagles are very traditional in their selection of nest‐site and so for Skye we substituted nest locations recorded in the 1992 national survey (Green Citation1996). In order to test whether this will have made any difference to the results, we examined differences in nest‐site elevation and distance to water between 1992 and 2003 nest‐sites on Mull (where, like Skye, occupied Golden Eagle territory numbers were similar between surveys). There was no significant difference between 1992 and 2003 survey results in the frequency distributions for nest‐site elevation (Kolmogorov–Smirnov z = 0.471, P = 0.980) and distance to water (Kolmogorov–Smirnov z = 0.447, P = 0.988). Thus, we assumed that our use of 1992 nest locations for Skye was acceptable.
To estimate land cover in the GIS we used broad habitat classes of the Land Cover Map 2000 (LCM2000; Fuller et al. Citation2002). LCM2000 is derived from a computer classification of satellite scenes, obtained mainly from landsat. Woodland cover was taken from the National Inventory of Woodlands and Trees (Forestry Commission Citation2001).
We measured the following ‘habitat‐based’ variables in the GIS: (a) altitude (m above sea level); (b) proportion of land cover within 1 km of (i) woodland and (ii) open water; (c) distance to nearest (i) sea; (ii) open water (sea or large freshwater body > 1 km2); (iii) road; (iv) settlement; (v) wood; (vi) conspecific nest‐site; (vii) nest of ‘other’ eagle species. Although proximity to water and woodland are likely to be correlated to some extent with altitude (Table ), freshwater wetlands occur in Scotland at a range of altitudes and woodland cover is geographically variable, so we considered these three variables separately.
Table 1. Matrix of Pearson correlation coefficients among landscape characteristics of Golden and White‐tailed Eagle nest‐sites in western Scotland.
Statistical analysis
Differences in nesting habitat
We examined differences in the nesting habitat of the two eagle species at two spatial scales. First, we examined differences in the precise location of nests by comparing the frequency of nesting in trees and cliffs by each species using Fisher’s exact test. Second, we modelled nest‐site location using wider geographical variables to identify any differences in the landscape context of nests between species. The latter was conducted using logistic regression in minitab version 15.0 with species (White‐tailed [1] or Golden Eagle [0]) as a response variable and the ‘habitat‐based’ explanatory variables (described previously) as predictor variables. Model simplification from a full model was conducted using backwards deletion of non‐significant terms (P > 0.05) to produce a final minimum adequate model. Region was included as a categorical variable. Two models were produced. The first compared White‐tailed Eagle territories with all Golden Eagle territories, while the second examined only the nearest Golden Eagle neighbours of White‐tailed Eagles and, therefore, examined finer‐scale differences.
Golden Eagle tolerance
We compared the distance from nest‐sites of Golden Eagles that neighboured White‐tailed Eagles with the nearest nest‐sites of both species in order to investigate any apparent tolerance by Golden Eagles of one species over the other; this was done using two‐tailed t‐tests for all eagles, for White‐tailed and Golden Eagle neighbours only, and in each region of the study area. A similar comparison was not possible for White‐tailed Eagles because the patchy nature of their distribution at the current stage of re‐establishment meant that no meaningful distance to the nearest conspecific could be derived for isolated territories. For this reason, we did not include distance between eagle nests in our wider analysis of nest‐site characteristics.
Changes in Golden Eagle territory occupancy
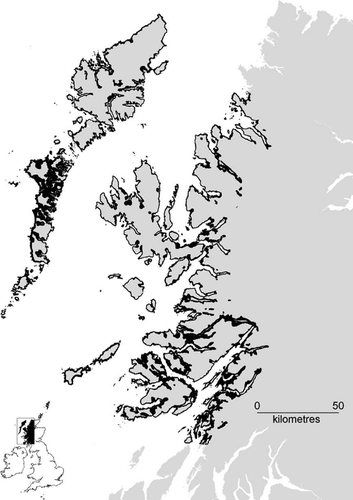
Because Golden Eagle productivity data were not widely available for most years in much of the study area, we took occupancy as an indirect indication of territory quality (Sergio & Newton Citation2003). Changes in Golden Eagle territory occupancy are likely to be related, at least in part, to habitat and land‐use (Whitfield et al. Citation2007). To account for this, and focus purely on those Golden Eagles occupying potential White‐tailed Eagle habitat, we constructed a GIS layer in idrisi kilimanjaro (Clark Laboratories, Worcester, MA) of potential White‐tailed Eagle nesting habitat within the study area, based on the results of the logistic regression model separating White‐tailed from Golden Eagle nests (Fig. ). This model was used to identify all known Golden Eagle territories (whether occupied or not in 2003) potentially influenced by re‐establishing White‐tailed Eagles. Golden Eagle territories with a nest‐site within 6 km of a White‐tailed Eagle nest‐site were classed separately from more distant territories (6 km typically represents the maximum territorial Golden Eagle ranging distance in our study area [McGrady et al. Citation2002, Haworth et al. Citation2006]). The change in status of these territories between the 1992 and 2003 national surveys (Green Citation1996; Eaton et al. Citation2007) – during which time the White‐tailed Eagle population increased from 8 to 31 pairs – was classed as ‘deterioration’, ‘no change’ or ‘improvement’, according to whether territories were occupied by fewer, the same number, or more birds, respectively, in the 2003 survey. We compared the frequency of the status of each territory using Fisher’s exact test (a) within the habitat model, according to whether nests were within 6 km of White‐tailed Eagles; and (b) within 6 km of White‐tailed Eagles, according to whether nests were within the model of suitable White‐tailed Eagle habitat. In order to check for wider‐scale change, we made similar comparisons of the frequency of the status of Golden Eagle territories, but across the whole study area, using the G‐test.
Figure 2 Geographical information system model of potential White‐tailed Eagle nesting habitat (solid black) within the study area (see text for details).
RESULTS
Differences in nesting habitat
Nest‐site choice differed between species according to the use of trees or cliffs. All Golden Eagles in the study area nested on cliffs (n = 252), whereas more than half of White‐tailed Eagle nests were in trees (57.1%, n = 35) (Fisher’s exact test P < 0.001). There was a clear difference between the three regions in the classification of White‐tailed Eagle territories by nest type, with no tree sites (n = 11) in the Western/Small Isles, two‐thirds of sites in trees (n = 12) in Skye/Ross‐shire and all nests in trees (n = 12) in Argyll. These regional differences in the use of trees reflected regional differences in the extent of woodland and in the proximity of woodland to, and extent of woodland around, nest‐sites. There was no difference between the median altitudes of cliff and tree nest‐sites used by White‐tailed Eagles, but cliff nests (median = 0.02 km, n = 16) were closer to water than tree nests (median = 0.2 km, n = 19; U = 88.5; P = 0.035).
At a larger scale, White‐tailed Eagles were more likely to nest at lower altitudes, and with a greater proportion of surrounding woodland, than Golden Eagles (Fig. ). Importantly, the two variables were apparent in both models. Distance to the nearest large water body approached significance as a predictor variable (P = 0.062) in the analysis comparing White‐tailed Eagle nest locations against all Golden Eagle nest locations (Table ). Because of the strength of this relationship, we retained it when modelling our GIS layer of potential White‐tailed Eagle nesting habitat.
Figure 3 Altitude (a), woodland cover within 1 km (b) and distance to large water body (c) for nests of Golden and White‐tailed Eagles in three areas of Scotland – Argyll (A); Skye/Ross‐shire (S/R); and the Western Isles (WI). Plots show means and 95% CIs.
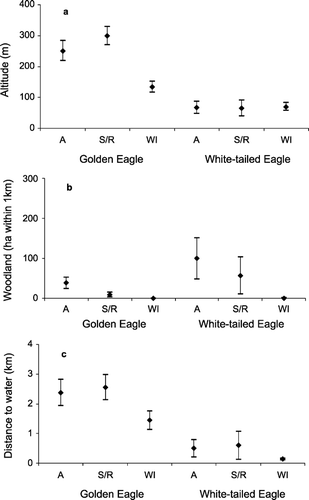
Table 2. Logistic regression tables for minimum adequate predictive models of nest habitat selection by Golden and White‐tailed Eagles.
Golden Eagle tolerance
Golden Eagles nested significantly closer to White‐tailed Eagles than to other Golden Eagles across the whole study area and in each of the three smaller regions (Table ). This was apparent despite the fact that there were fewer White‐tailed Eagle territories than Golden Eagle territories, which would be expected to result in greater distances between Golden and White‐tailed Eagles.
Table 3. Mean distance in km (95% CI) to nearest Golden Eagle (GE) nest‐sites from nest‐sites of conspecifics (GE to GE) and White‐tailed Eagles (WTE to GE) according to region.
Changes in Golden Eagle territory occupancy
Fifteen (43%) White‐tailed Eagle nest locations fell within our GIS model of potential White‐tailed Eagle habitat, with 29 nests (83%) within 100 m and 33 (94%) within 250 m, indicating that our model adequately described White‐tailed Eagle nesting habitat. A total of 54 Golden Eagle territory centres fell within the 3359 km2 (27%) of the study area that overlapped the potential White‐tailed Eagle habitat plus a 250‐m buffer (Fig. ). The change in occupancy of the 20 Golden Eagle nests within 6 km of White‐tailed Eagle nests between national Golden Eagle surveys in 1992 and 2003 did not differ from change in occupancy of the remainder of nests in suitable White‐tailed Eagle habitat (Fisher’s exact test P = 0.378; Table ). Within 6 km of White‐tailed Eagles, more Golden Eagle territories showed reduced occupancy outside our White‐tailed Eagle habitat model than within it, although the difference was not significant (Fisher’s exact test P = 0.173). Examining change of occupancy status of Golden Eagle territories over our whole study area, we again found no difference according to suitability for White‐tailed Eagles based on our GIS model (G = 1.212; df = 1; P = 0.271), or proximity to White‐tailed Eagles (G = 0.066; df = 1; P = 0.798).
Table 4. Change in status between 1992 and 2003 of known Golden Eagle home ranges according to proximity to breeding White‐tailed Eagles and overlap with potential White‐tailed Eagle habitat in western Scotland.
DISCUSSION
Our results showed significant differences in nesting habitat selected by Golden Eagles and White‐tailed Eagles in Scotland, even when considering nest‐sites only of neighbouring pairs (and thus excluding many Golden Eagle sites likely to magnify any difference). These differences are in line with our hypotheses, with White‐tailed Eagles nesting at lower elevations than Golden Eagles and tending to nest in more wooded areas and closer to water. Elevation is likely to be correlated to some extent with proximity to, and extent of, both open water and woodland, as well as with human influence. In our eagle nest data set, altitude was strongly correlated with distance to sea and open water and negatively correlated with the area of sea and open water within 1 km (Table ). Thus, the absence of water‐features as significant explanatory variables in our minimum adequate models can in part be explained by their correlation with altitude.
All Golden Eagle nest‐sites in the study area in 2003 were on cliffs, even in relatively wooded locations. Although tree nesting by Golden Eagles has occurred in the study area, it is uncommon (Watson Citation1997). On the other hand, half of our White‐tailed Eagle nests were in trees and this species appeared to nest in trees where these were available. In areas lacking suitable trees, White‐tailed Eagles nested on cliffs, but selected lower altitude sites than their Golden Eagle neighbours. Cliffs used by White‐tailed Eagles for nesting were nearly all sea cliffs. This was also true for Golden Eagle nest‐sites in our GIS model of White‐tailed Eagle nesting habitat, although here too Golden Eagle sites were at higher elevations than White‐tailed Eagle nests. There was some regional variation and nest‐sites of both eagles were most similar to each other in the treeless Western Isles, where there was also the smallest difference in nest‐site altitude. However, these differences were insufficient for ‘region’ to appear as an explanatory variable in our minimum adequate models.
Distinct choice of nesting habitat when breeding in close proximity to one another suggests that, in western Scotland, White‐tailed Eagles and Golden Eagles may partition breeding habitat by altitude. Golden Eagles tend to nest close to the natural ‘tree line’ (Bergo Citation1984, Watson Citation1997), at the top edge of potential natural woodland. White‐tailed Eagles tend to nest at lower elevations, where woodland cover is greater and suitable nest trees more available. Recent information on diet of White‐tailed Eagles and sympatric Golden Eagles (Marquiss et al. Citation2003, Madders & Marquiss Citation2003) indicates stronger differentiation than that found in an earlier study (Watson et al. Citation1992), with White‐tailed Eagles taking a wider range of prey that included more seabirds and fish than Golden Eagles, while Golden Eagles took more lagomorphs. In some cases, White‐tailed Eagles flew several kilometres from their nests to take seabirds, crossing more than one Golden Eagle territory to do so. Shared food types were likely to be found at a wide range of elevations (Whitfield et al. unpubl. data). Sympatric predators may mediate potential competition by obtaining shared food types from different locations (Gerstell & Bednarz Citation1999). This seems to be the case for Scottish eagles because, while there may be some overlap in habitat use and food supplies, overall there appears to be a larger degree of separation, principally by elevation.
Taken together, this suggests partitioning of both nesting and foraging habitat. Golden Eagles hunt hills above their tree line nests for prey, which is then carried downhill to provision incubating females or chicks. White‐tailed Eagles forage over wetland, coastal or marginal habitats for a wider range of more aquatic prey (Whitfield et al. unpubl. data), which is then carried back to nests in flights that involve net gain in height. This is consistent with differences in wing morphology between the two species. White‐tailed Eagle wings have lower aspect‐ratio and wing‐loading (Alerstam et al. Citation2007) and a squarer and proportionately larger handwing (Kirmse Citation1998), and thus are theoretically better suited to level, active, load‐carrying flight than Golden Eagles.
Even if preference for slightly different ecological niches has resulted from gradual physical adaptation prompted by stronger competition in the past, the current situation suggests that competition between the two species is minimized owing to differences in nesting and probably also foraging habitat. Therefore, negative effects of reintroduced White‐tailed Eagles on established populations of Golden Eagles may well be less than previously suggested (Watson Citation1997, Halley Citation1998). Indeed, if niche partitioning is a result of White‐tailed Eagles avoiding Golden Eagles (Halley & Gjershaug Citation1998), ultimately the potential expansion of White‐tailed Eagles in the west of Scotland could be limited by the presence of Golden Eagles.
Around 40% of Scotland’s Golden Eagle territories are found in the study area and one third of these overlap with potential White‐tailed Eagle habitat. Overlap between the species is greatest in the Hebridean islands off the west coast of Scotland. Golden Eagle numbers here increased by 28% between 1992 and 2003 (Eaton et al. Citation2007), during the same time that the rate of increase in White‐tailed Eagle numbers has accelerated (Evans et al. Citation2009). This suggests that, at current densities, neither food supply nor availability of nest‐sites are limiting for either species. Although individual nest‐sites in Norway and Scotland are known to have been occupied by both species at different times (Willgohs Citation1961, Bergo Citation1987, Watson Citation1997, Crane & Nellist Citation1999, Whitfield et al. Citation2002, Love Citation2003), such dual use has been extremely rare in Scotland since reintroduction. The expansion of White‐tailed Eagles on Mull has had no discernible effect on Golden Eagles (Whitfield et al. Citation2002). More recent expansion on Skye and the Western Isles has revealed no obvious adverse influence on the local Golden Eagle populations (numbers of Golden Eagles have remained stable or expanded, respectively, with no decline in productivity [Eaton et al. Citation2007]). We did not investigate possible competitive effects on eagle breeding success, largely because data for Golden Eagle productivity were lacking for much of the study area and in particular for areas where nesting habitat of both species was most similar. The only metric we were able to apply across the whole study area was territory occupancy, which is a good indication of territory quality in many species (Sergio & Newton Citation2003). Our study found no effect of proximity of White‐tailed Eagles on the occupancy status of Golden Eagle territories with characteristics apparently better suited to White‐tailed Eagles (and thus where one might expect to detect the earliest indications of competitive exclusion).
In the past, some White‐tailed Eagles nested at higher elevations in Scotland than those recorded in this study (Baxter & Rintoul Citation1953, Love Citation1983). The historical record includes some cases where persecution appears to have driven White‐tailed Eagles from preferred low‐lying habitats to higher sites that were less accessible to humans (MacKenzie Citation1921). However, in at least one location, White‐tailed Eagles occupied a relatively high‐elevation area apparently because it was simply not suitable for Golden Eagles. Rannoch Moor (56° 37’ N, 4° 45’ W) is an extensive, flat basin, with both wetland and woodland habitats. Golden Eagles nest (and historically nested) in the surrounding hills, but are (and appear historically to have been) absent from the flat moor itself. Our study area includes some large tracts of relatively flat and wet land, but none is as extensive or at such a high elevation as Rannoch Moor, which can probably justifiably be described as unique in Scotland. Its occupation by White‐tailed Eagles may also have been exceptional. The fact that White‐tailed Eagles became extinct in Scotland suggests that an upper altitude limit may have prevented the preservation of a ‘reservoir’ of the species in high altitude locations, from which subsequent recolonisation might have been possible (in contrast to the likely pattern for Golden Eagle [Love Citation1983]). Therefore, although it is possible that future range expansion by White‐tailed Eagles may require our model to be refined to take account of relief, and possibly also a higher altitude limit, the historical evidence suggests that it should be fundamentally sound.
Given the comparative shortage of mature woodland in many parts of western Scotland and as eagle nests require large, robust trees (Shiraki Citation1994, Helander & Stjernberg Citation2003), it is also possible that White‐tailed Eagle expansion may be limited by a lack of suitable nest trees. Our results, therefore, highlight: (1) the importance of woodland as nesting habitat for White‐tailed Eagles; and (2) the likelihood that any future negative effects of interspecific competition may be most probable where nest trees are absent and competition is primarily over nest‐sites. In the short‐term, provision of artificial nests (Mecionis Citation2003) in areas where prospecting immatures are known to occur, may prove beneficial, or at least illuminating as to the effectiveness of such management if conducted rigorously. In the longer term, woodland management undertaken now, such as planting or regeneration of native woodland, or management of existing conifer plantations, could bring long‐term benefits to White‐tailed Eagles (and other species) if carried out in appropriate locations throughout the Western Highlands and Islands.
ACKNOWLEDGEMENTS
We thank Jamie Boyle, Roger Broad, Martin Carty, Ken Crane, Fiona Harmer, Sean Morris, Kate Nellist, David Sexton and Matt Wilson for their contribution in the field. The 1992 and 2003 national surveys of Golden Eagles were undertaken by the Scottish Raptor Study Groups, the Royal Society for the Protection of Birds and Scottish Natural Heritage. We are extremely grateful to the dedication, enthusiasm and skills of the many voluntary eagle workers in Scotland who contributed to the surveys (acknowledged in Eaton et al. [Citation2007]) and continue to give selflessly to the conservation of Golden Eagles in Scotland. Ian Dillon deserves special thanks through his co‐ordination of and contribution to the non‐voluntary effort in the 2003 survey. David McLeod, Paul Britten and Will George assisted with GIS analyses. We thank Fabrizio Sergio and an anonymous referee for constructive comments on the manuscript.
Notes
a. Owing to the recording resolution for nest‐sites, actual grid reference locations may have been ± 50 m of those recorded. For nests on cliffs, where altitude changes rapidly with horizontal location as recorded by cartographic grid reference, the recorded resolution may have given biased estimates of ‘true’ nest location altitude. We attempted to overcome this potential bias by manually checking all nest‐site locations against a higher resolution digital terrain model (± 5 m) in the GIS. We accepted the recorded location if its grid reference was on the nearest cliff unless (in exceptional cases) this was away from a cliff, when we took the elevation from the mid‐point of the nearest cliff. This exercise may not have circumvented all potential biases, but remaining errors should have applied equally to cliff nests of either species. We also had no reason to believe, after manually checking every grid reference, that residual errors were biased towards any altitudinal direction.
Related Research Data
REFERENCES
- Alerstam , T. , Rosén , M. , Bäckman , J. , Ericson , P.G.P. and Hellgren , O. 2007 . Flight speeds among bird species: allometric and phylogenetic effects . PLoS Biol. , 5 : 1656 – 1662 .
- Aumann , T. 2001 . The structure of raptor assemblages in riparian environments in the south‐west of the Northern Territory, Australia . Emu , 101 : 293 – 304 .
- Bainbridge , I.P. , Evans , R.J. , Broad , R.A. , Crooke , C.H. , Duffy , K. , Green , R.E. , Love , J.A. and Mudge , G.P. 2003 . “ Re‐introduction of White‐tailed Eagles (Haliaeetus albicilla) to Scotland ” . In Birds of Prey in a Changing Environment , Edited by: Thompson , D.B.A. , Redpath , S.M. , Fielding , A.H. , Marquiss , M. and Galbraith , C.A. 393 – 406 . Edinburgh , , UK : The Stationery Office .
- Baxter , E.V. and Rintoul , J.V. 1953 . The Birds of Scotland , Edinburgh , , UK : Oliver & Boyd .
- Bechard , M.J. , Knight , R.L. , Smith , D.G. and Fitzner , R.E. 1990 . Nest sites and habitats of sympatric hawks (Buteo spp.) in Washington . J. Field Ornithol. , 61 : 159 – 170 .
- Bergo , G. 1984 . Habitat and nest site features of Golden Eagle Aquila chrysaetos (L.) in Hordaland, west Norway . Fauna norv. Ser. C. Cinclus , 7 : 109 – 113 .
- Bergo , G. 1987 . Territorial behaviour of Golden Eagles in western Norway . Br. Birds , 80 : 361 – 376 .
- Burton , A.M. and Olsen , P. 2000 . Niche partitioning by two sympatric goshawks in the Australian wet tropics: ranging behaviour . Emu , 100 : 216 – 226 .
- Carrete , M. , Sánchez‐Zapata , J.A. , Calvo , J.F. and Lande , R. 2005 . Demography and habitat availability in territorial occupancy of two competing species . Oikos , 108 : 125 – 136 .
- Crane , K. and Nellist , K. 1999 . Island Eagles: 20 Years Observing Golden Eagles on the Isle of Skye , Glenbrittle , , UK : Cartwheeling Press .
- Dennis , R.H. , Ellis , P.M. , Broad , R.A. and Langslow , D.R. 1984 . The status of the Golden Eagle in Britain . Br. Birds , 77 : 592 – 607 .
- Eaton , M.A. , Dillon , I.A. , Stirling‐Aird , P. and Whitfield , D.P. 2007 . The status of the Golden Eagle Aquila chrysaetos in Britain in 2003 . Bird Study , 54 : 212 – 220 .
- Evans , R.J. , Broad , R.A. , Duffy , K. , MacLennan , A.M. , Bainbridge , I.P. and Mudge , G.P. Re‐establishment of a breeding population of White‐tailed Eagles in Scotland . Proceedings from an International Conference . September 13–17 2000 , Björkö , Sweden. Sea Eagle 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 397 – 403 . Stockholm , , Sweden : Swedish Society for Nature Conservation .
- Evans , R.J, , Wilson , J.D. , Amar , A. , Douse , A. , MacLennan , A. , Ratcliffe , N. and Whitfield , D.P. 2009 . Growth and demography of a re‐introduced population of White‐tailed Eagles Haliaeetus albicilla . Ibis , 151 : 244 – 254 .
- Fielding , A.H. , Haworth , P.F. , Morgan , D.H. , Thompson , D.B.A. and Whitfield , D.P. 2003 . “ The impact of Golden Eagles on a diverse bird of prey assemblage ” . In Birds of Prey in a Changing Environment , Edited by: Thompson , D.B.A. , Redpath , S.M. , Fielding , A.H. , Marquiss , M. and Galbraith , C.A. 221 – 244 . Edinburgh , , UK : The Stationery Office .
- Forestry Commission . 2001 . National Inventory of Woodland and Trees – Scotland , Edinburgh , , UK : Forestry Commission .
- Fuller , R.M. , Smith , G.M. , Sanderson , J.M. , Hill , R.A. and Thomson , A.G. 2002 . The UK Land Cover Map 2000: construction of a parcel‐based vector map from satellite images . Cartogr. J. , 39 : 15 – 25 .
- Gainzarain , J.A. , Arambarri , R. and Rodríguez , A.F. 2000 . Breeding density, habitat selection and reproductive rates of the Peregrine Falcon Falco peregrinus in Álava (northern Spain) . Bird Study , 47 : 225 – 231 .
- Gainzarain , J.A. , Arambarri , R. and Rodríguez , A.F. 2002 . Population size and factors affecting the density of the Peregrine Falcon Falco peregrinus in Spain . Ardeola , 49 : 67 – 74 .
- Gerstell , A.T. and Bednarz , J.C. 1999 . Competition and patterns of resource use by two sympatric raptors . Condor , 101 : 557 – 565 .
- Green , R.E. 1996 . The status of the Golden Eagle in Britain in 1992 . Bird Study , 43 : 20 – 27 .
- Hakkarainen , H. and Korpimäki , E. 1996 . Competitive and predatory interactions among raptors: an observational and experimental study . Ecology , 77 : 1134 – 1142 .
- Halley , D.J. 1998 . Golden and White‐tailed Eagle in Scotland and Norway: co‐existence, competition and environmental degradation . Br. Birds , 91 : 171 – 179 .
- Halley , D.J. and Gjershaug , J.O. 1998 . Inter‐and intra‐specific dominance relationships and feeding behaviour of Golden Eagles Aquila chrysaetos and Sea Eagles Haliaeetus albicilla at carcasses . Ibis , 140 : 295 – 301 .
- Haworth , P.F. , McGrady , M.J. , Whitfield , D.P. , Fielding , A.H. and McLeod , D.R.A. 2006 . Ranging distance of resident Golden Eagles Aquila chrysaetos in western Scotland according to season and breeding status . Bird Study , 53 : 265 – 273 .
- Helander , B. and Stjernberg , T. Action Plan for the Conservation of the White‐tailed Sea Eagle Haliaeetus albicilla . Convention on the Conservation of European Wildlife and Natural Habitats . Strasbourg , France.
- IUCN . Translocation of living organisms . IUCN position statement . September 4 1987 . http://www.iucnsscrsg.org/download/IUCNPositionStatement.pdf (accessed May 2010)
- IUCN . 1998 . “ Guidelines for reintroductions ” . http://www.iucnsscrsg.org/download/English.pdf (accessed May 2010)
- Jaksić , F.M. and Braker , H.E. 1983 . Food–niche relationships and guild structure of diurnal birds of prey: competition versus opportunism . Can. J. Zool. , 61 : 2230 – 2241 .
- Katzner , T.E. , Bragin , E.A. , Knick , S.T. and Smith , A.T. 2003 . Coexistence in a multispecies assemblage of eagles in central Asia . Condor , 105 : 538 – 551 .
- Kirmse , W. 1998 . “ Morphometric features characterizing flight properties of Palearctic eagles ” . In Holarctic Birds of Prey , Edited by: Chancellor , R.D. , Meyburg , B.‐U. and Ferrero , J.J. 339 – 348 . Berlin , , Germany : World Working Group on Birds of Prey .
- Kostrzewa , A. 1991 . Interspecific interference competition in three European raptor species . Ethol. Ecol. Evol. , 3 : 127 – 143 .
- Krüger , O. 2002 . Interactions between Common Buzzard Buteo buteo and Goshawk Accipiter gentilis: trade‐offs revealed by a field experiment . Oikos , 96 : 441 – 452 .
- Krüger , O. , Liversidge , R. and Lindström , J. 2002 . Statistical modelling of the population dynamics of a raptor community in a semi‐desert environment . J. Anim. Ecol. , 71 : 603 – 613 .
- Lack , D. 1946 . Competition for food by birds of prey . J. Anim. Ecol. , 15 : 123 – 129 .
- Love , J.A. 1983 . The Return of the Sea Eagle , Cambridge , , UK : Cambridge University Press .
- Love , J.A. A history of the White‐tailed Sea Eagle in Scotland . Proceedings from an International Conference . September 13–17 2000 , Björkö , Sweden. Sea Eagle 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 39 – 50 . Stockholm , , Sweden : Swedish Society for Nature Conservation .
- MacKenzie , O. 1921 . One Hundred Years in the Highlands , London : Edward Arnold .
- McGrady , M.J. 1997 . Aquila chrysaetos Golden Eagle . Birds West. Palearct. Update , 1 : 99 – 114 .
- McGrady , M.J. , Grant , J.R. , Bainbridge , I.P. and McLeod , D.R.A. 2002 . A model of Golden Eagle (Aquila chrysaetos) ranging behavior . J. Raptor Res. , 36 (1 Suppl.) : 62 – 69 .
- Madders , M. and Marquiss , M. A comparison of the diet of White‐tailed Eagles and Golden Eagles breeding in adjacent ranges in west Scotland . Proceedings from an International Conference . September 13–17 2000 , Björkö , Sweden. Sea Eagle 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 289 – 296 . Stockholm , , Sweden : Swedish Society for Nature Conservation .
- Marquiss , M. , Madders , M. , Irvine , J. and Carrs , D.N. The impact of White‐tailed Eagles on sheep farming on Mull. Final report . Banchory : Centre for Ecology and Hydrology .
- Mecionis , R. Artificial nests for White‐tailed Eagle . Proceedings from an International Conference . September 13–17 2000 , Björkö , Sweden. Sea Eagle 2000 , Edited by: Helander , B. , Marquiss , M. and Bowerman , W. pp. 377 – 378 . Stockholm , , Sweden : Swedish Society for Nature Conservation .
- Petty , S.J. , Anderson , D.I.K. , Davison , M. , Little , B. , Sherratt , T.N. , Thomas , C.J. and Lambin , X. 2003 . The decline of Common Kestrels Falco tinnunculus in a forested area of northern England: the role of predation by Northern Goshawks Accipiter gentilis . Ibis , 145 : 472 – 483 .
- Poole , K.G. and Bromley , R.G. 1988 . Interrelationships within a raptor guild in the central Canadian arctic . Canadian Journal of Zoology , 66 : 2275 – 2282 .
- Ratcliffe , D.A. 1993 . The Peregrine Falcon, , 2nd edn. , London : T. & A.D. Poyser .
- Selås , V. 1996 . Selection and reuse of nest stands by Sparrowhawks Accipiter nisus in relation to natural and manipulated variation in tree density . J. Avian Biol. , 27 : 56 – 62 .
- Sergio , F. and Newton , I. 2003 . Occupancy as a measure of territory quality . J. Anim. Ecol. , 72 : 857 – 865 .
- Sergio , F. , Marchesi , L. and Pedrini , P. 2003 . Spatial refugia and the coexistence of a diurnal raptor with its intraguild owl predator . J. Anim. Ecol. , 72 : 232 – 245 .
- Sergio , F. , Rizzolli , F. , Marchesi , L. and Pedrini , P. 2004 . The importance of interspecific interactions for breeding‐site selection: Peregrine Falcons seek proximity to ravens nests . Ecography , 27 : 818 – 826 .
- Shiraki , S. 1994 . Characteristics of White‐tailed Sea Eagle nest sites in Hokkaido, Japan . Condor , 96 : 1003 – 1008 .
- Vrezec , A. and Tome , D. 2004 . Altitudinal segregation between Ural Owl Strix uralensis and Tawny Owl S. aluco: evidence for competitive exclusion in raptorial birds . Bird Study , 51 : 264 – 269 .
- Watson , J. 1997 . The Golden Eagle , London : T. & A.D. Poyser .
- Watson , J. , Leitch , A.F. and Broad , R.A. 1992 . The diet of the Sea Eagle Haliaeetus albicilla and Golden Eagle Aquila chrysaetos in western Scotland . Ibis , 134 : 27 – 31 .
- Whitfield , D.P. , Evans , R.J. , Broad , R.A. , Fielding , A.H. , Haworth , P.F. , Madders , M. and McLeod , D.R.A. 2002 . Are reintroduced White‐tailed Eagles in competition with Golden Eagles? . Scott. Birds , 23 : 36 – 45 .
- Whitfield , D.P. , Fielding , A.H. , McLeod , D.R.A. , Morton , K. , Stirling‐Aird , P.K. and Eaton , M.A. 2007 . Factors constraining the distribution of Golden Eagles Aquila chrysaetos in Scotland . Bird Study , 54 : 199 – 211 .
- Whitfield , D.P. , Douse , A. , Evans , R.J. , Grant , J. , Love , J. , McLeod , D.R.A. , Reid , R. and Wilson , J.D. 2009 . Natal and breeding dispersal in a reintroduced population of White‐tailed Eagles Haliaeetus albicilla . Bird Study , 56 : 177 – 186 .
- Wiens , J.A. 1989 . The Ecology of Bird Communities, Vol. 2. Processes and Variations , Cambridge , , UK : Cambridge University Press .
- Willgohs , J.F. 1961 . The White‐tailed Eagle Haliaeetus albicilla albicilla (L.) in Norway , Bergen , , Norway : Norwegian University Press .