Abstract
Capsule The capture–recapture model M(o) is an efficient way to estimate local population size.
Aims To test if a single capture–recapture modelling approach, combined with a simple survey method, can produce estimates of local population size from a dataset involving large‐scale multi‐observer surveys
Methods We sampled the presence of Nightjars in three separate sessions at three forests. Territory numbers were estimated using conventional territory‐mapping criteria. We ran different capture–recapture models to analyse the detection histories of territories obtained across the three sampling sessions and in the three different forests, using either only registrations of churring birds or all contacts.
Results The capture–recapture model M(o), assuming a constant detection probability, was the most efficient one to produce estimates of local population size. Using only two of the three sampling sessions gave less precise, though quite similar, estimates of the number of territories, with standard deviations representing 5–10% of the estimate values. However, this was reduced to 0.7–3.5%, i.e. three to seven times lower, when using the three sessions.
Conclusion Repeated sampling sessions to map territories can be efficiently used within the capture–recapture model M(o) to estimate detection probability and produce precise estimates of local population size.
European Nightjars Caprimulgus europaeus are a key breeding species of dry, open habitats including open coniferous woodlands, clear‐fells in plantations, lowland heaths and their woodlands edges, and also broad‐leaved coppiced coupes. Nightjars are primarily crepuscular and nocturnal, being most active around dusk and dawn. They are insectivorous long‐distance migrants which visit Europe to breed during May–August. Nightjars have been identified as a species of conservation concern at the European level on account of historical declines in their population sizes and ranges, and are listed on Annex I of the EU ‘Birds’ directive (Directive on the conservation of wild birds 79/409/EEC) and should, therefore, be subject to special conservation measures concerning their habitats by EU member states. The long‐term and substantial loss, degradation and fragmentation of lowland heathland habitats has been the major factor associated with the observed population declines (Langston et al. Citation2007).
Estimating the population size of Nightjars is necessary to identify key sites and estimate population change (negative or positive) or stability, especially as detection probability might be density dependent (Williams & Berkson Citation2004). The sizes of local populations of Nightjars have usually been estimated using standard territory‐mapping procedures to register ‘churring’ males (Morris et al. Citation1994), while large‐scale censuses have relied on the participation of numerous volunteer observers (Conway et al. Citation2007) . In such schemes, Nightjar detectability could potentially vary among observers, sampling events and sites, because of varying observers’ skills, ongoing breeding cycle and associated behaviours and habitat‐ or density‐dependent factors. The development of closed capture models and their applications to capture–recapture histories of detected individuals or territories could allow such potential biases to be taken into account when estimating population sizes, by allowing the estimates to be corrected for non‐perfect and potentially heterogeneous detection probabilities (Rexstad & Burnham Citation1991, Ganter & Madsen Citation2001). Here we have applied closed capture models to census data of Nightjar territories collected in three forests of western France. We have used this to test the efficiency of territory‐mapping procedures to produce precise population estimates, and to define which capture–recapture model is best used to obtain unbiased estimates. The closure assumption is necessary as there is no way to identify individually the detected birds, such individual identification would have allowed the detection of transient birds within the global population.
METHODS
Study area and sampling design
The survey was carried out in three forests of the Vienne département, western France, by multiple observers: Moulière (covering 4200 ha; 47 observers; Fig. ), Vouillé (1630 ha; 10 observers) and Mareuil (670 ha; 7 observers; Fig. ). Each forest was divided in sectors covering between 200 and 300 ha each (on average 252 ha), each sector being attributed to one observer in charge of its survey. In each forest, three sampling sessions were organised: the first in the first half of June, the second in the second half of June, and the third in the first half of July. There was an average of 15 days between each sampling session Nightjars arrive from migration on average on 6 May in the region (T. Williamson unpubl. data). Occasionally, the observers differed between the sampling sessions. In each forest and for each session, all sectors were surveyed simultaneously by the observers, walking, biking or driving slowly all available roads and tracks, and noting all contacts with Nightjars on 1:10 000 scale maps of their sector. A distinction was made between churring males and other types of contact (alarm call, flight call, visual detection). Observers did not use tape‐lurring. Finally, the second author followed standard territory‐mapping procedures (Bibby et al. Citation1997) to register ‘churring’ males. The second author summarised all collected maps in a global territory map for each forest, using information on multiple and simultaneous contacts to disentangle neighbouring territories. An ‘observed territory’ was defined by a location or set of locations where at least one churring bird was recorded, using simultaneous contacts with different males to separate adjacent territories. Finally, a matrix of detection histories – across the three sampling events – was built up for the territories within each forest, using either only contacts with churring males or all contacts. For example, a territory where a churring male was recorded during the first and the third sampling session is denoted ‘101’, etc. In Moulière, 163 observed territories were mapped (total of 315 contacts, of which 288 were churring contacts), in Vouillé 46 observed territories were mapped (total contacts 110, of which 102 churring contacts), and in Mareuil 36 observed territories were mapped (total contacts 80, 75 churring contacts).
Figure 1 Locations of male Nightjar detections and territories within the woodland of Moulière (4200 ha), Vienne, France. Detections obtained at each of the three sampling sessions are represented with different colours, while polygons encompass individual territories deduced by a territory‐mapping procedure.
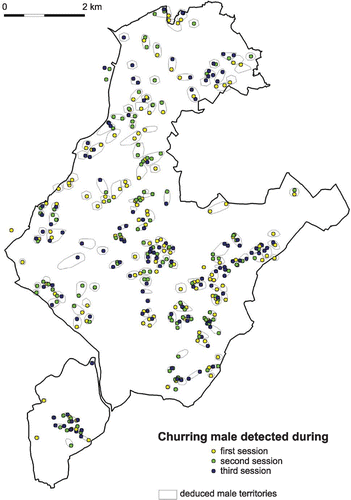
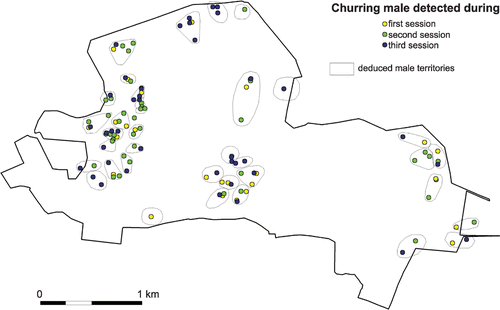
Figure 2 Location of male Nightjar detections and territories within the woodland of Mareuil (670 ha), Vienne, France. Detections obtained at each of the three sampling sessions are represented with different colours, while polygons encompass individual territories deduced by a territory‐mapping procedure.
Capture–recapture analyses
We used probabilistic capture–recapture models developed to estimate the size of closed animal populations (Otis et al. Citation1978, Pollock et al. Citation1990). We used the program capture and its model selection algorithm (Rexstad & Burnham Citation1991) to investigate the relevance of different capture–recapture models for estimating population size within each forest. Among the eight capture–recapture models initially proposed by Otis et al. (Citation1978) and relevant for population size estimation, the simplest one, labelled M(o), considers no effect on the probability of detection. Three models, labelled M(t), M(b) and M(h), assume respectively an effect of time (between sessions), behaviour (difference in detection probability between the first detection and the others) or heterogeneity among territories upon the probability of detection. The four remaining models, namely M(bh), M(th), M(tb) and M(tbh), allow for testing combinations of the aforementioned effects. The program capture includes a selection algorithm allowing the user to select the model that best fits the data. The program was used to select the best model for each forest. In case of model M(tbh) being the best‐fitted model, the model presenting the second best score was retained since there is no estimator available for M(tbh).
RESULTS
In four of the six studied cases, the model M(o) was selected as the best model, and estimates of local population sizes were thus obtained using this model (see Table ). For churring males at Mareuil, the model M(tbh) was selected as the best, and the second best model was M(o), with a score of 0.97. For churring males at Vouillé, the best selected model was M(t) (Table ), while M(o) was only the third best model with a score of 0.52 and an estimated 46 ± 1 sd (CI, 46–52) territories with an average detection probability of 0.73. This case is, therefore, the only one out of six where the detection probability is not considered as constant across territories, sampling sessions and detection events. Table presents the results of the estimated local population size of Nightjars in each forest, along with the estimated average detection probabilities.
Table 1. Observed and estimated numbers of Nightjar territories in the three study sites, using churring only or all contacts for the three sampling sessions to build up capture histories of observed occupied territories. The selected capture–recapture model is cited, as well as the average population size estimate ± sd (and 95% CI) and the average detection probability(‐ies) estimated by the model.
Table presents results of the two models selected previously (M(o) and M(t)), but applied to all possible pairs of sessions (first and second, first and third, and second and third) for churring birds or for all contacts, in the three forests. Using only two of the three sessions gave less precise, though quite similar, estimates, with standard deviations representing 5–10% of the estimate values, while this was reduced to 0.7–3.5%, i.e. three to seven times lower, when using the three sessions. Values obtained while using data from the second and third sessions were closer to those obtained while using the three sessions (compare Tables and ), and were associated with higher average detection probabilities compared with results obtained with the other pairs of sampling sessions (Table ).
Table 2. Estimated numbers of Nightjar territories in the three study sites using churring only or all contacts, for pairs of sampling sessions to build up capture histories of observed occupied territories. Results are presented for two different capture–recapture models, M(o) and M(t), and the associated population size estimate ± sd (and 95% confidence interval) and the average detection probability(‐ies).
DISCUSSION
Population estimation techniques combine a count of animals with an estimate of the fraction that went undetected. They produce an estimate of the total number of animals present along with a set of statistically determined upper and lower bounds to that estimate. Estimating Nightjar population size has often been achieved by conducting territory‐mapping surveys involving numerous volunteer observers, but without accounting for imperfect detection probability (Morris et al. Citation1994, Conway et al. Citation2007). Here, we estimated local population sizes in three different forests by using detection history of occupied territories across three sampling sessions within a capture–recapture framework (Williams et al. Citation2002). The estimates obtained were very similar to those measured directly by territory‐mapping procedures in two of the three study sites, but this did not hold true for the largest forest inhabited by the largest population.
The average detection probability of a territory was very high (between 0.53 and 0.87), given the nocturnal habits of the species, and was variable among forests, and in one case among the sampling sessions. Detection rates were higher in the two smallest forests, which were probably easier to survey. The capture–recapture model most often selected by the selection algorithm of the capture software, M(o), assumes a constant detection probability between sampling sessions and/or between the first and subsequent detection(s). Using three sampling sessions to build up the capture–recapture histories led to the most precise estimates, but it was necessary to run the selection algorithm to select the model fitting the data most closely. This allowed us to apply further the selected capture–recapture models to histories combining only two sessions, and to conclude that two late sampling sessions – from mid‐June to mid‐July – can be sufficient to obtain reasonable population size estimates if one accepts standard deviations representing up to 10% of the population size. If population sizes have to be known more precisely, for example if we want to detect possible changes of between 5% and 10%, there is an obvious need to consider three sampling sessions. Adding proof of territory occupancy obtained through non‐churring contacts also increased average detection probabilities and the precision of population size estimates.
The goal of this paper was not to provide estimates of local population sizes for Nightjars. We wished to investigate whether a simple survey method associated with a single modelling approach, namely the capture–recapture model M(o), can be used confidently to obtain estimates of local population size from a data set involving large‐scale multi‐observer surveys, and in which sampling conditions (habitat, events) can be either homogeneous or heterogeneous. The answer to this question is positive, and three field sessions dedicated to record Nightjar territories were sufficient to run models and estimate realistic population sizes. With some conditions on estimate errors, reducing the sampling effort to only two visits is possible. This might be an important point for nature conservationists wishing to assess the performance of sites designated as being of particular importance for the conservation of Nightjars (e.g. special protection areas), to determine the adequacy of the protected sites network for the species, and to assess the effectiveness of site‐based conservation measures on local population size (Langston et al. Citation2007).
ACKNOWLEDGEMENTS
We thank the Office National des Forêts (ONF) for providing access to the three national forests where we conducted our surveys, and Jean‐Pierre Moussus for providing helpful comments. This work is dedicated to all volunteers who have censused Nightjars for the survey: volunteers from LPO Vienne and staff from the ONF and Pinail nature reserve.
Related Research Data
REFERENCES
- Bibby , C.J. , Burgess , N.D. and Hill , D.A. 1997 . Bird Census Techniques , London , , UK : Academic Press .
- Conway , G. , Wotton , S. , Henderson , I. , Langston , R. , Drewitt , A. and Currie , F. 2007 . The status and distribution of European Nightjars Caprimulgus europaeus in the UK in 2004 . Bird Study , 54 : 98 – 111 .
- Ganter , B. and Madsen , J. 2001 . An examination of methods to estimate population size in wintering geese . Bird Study , 48 : 90 – 101 .
- Langston , R.H.W. , Wotton , S.R. , Conway , G.J. , Wright , L.J. , Mallord , J.W. , Currie , F.A. , Drewitt , A.L. , Grice , P.V. , Hoccom , D.G. and Symes , N. 2007 . Nightjar Caprimulgus europaeus and Woodlark Lullula arborea – recovering species in Britain? . Ibis , 149 (Suppl. 2) : 250 – 260 .
- Morris , A. , Burges , D. , Fuller , R.J. , Evans , A.D. and Smith , K.W. 1994 . The status and distribution of Nightjars Caprimulgus europaeus in Britain in 1992. A report to the British Trust for Ornithology . Bird Study , 41 : 181 – 191 .
- Otis , D.L. , Burnham , K.P. , White , G.C. and Anderson , D.R. 1978 . Statistical inference from capture data on closed animal populations . Wildlife Monographs , 62
- Pollock , K.H. , Nichols , J.D. , Brownie , C. and Hines , J.E. 1990 . Statistical inference for capture–recapture experiments . Wildlife Monographs , 107
- Rexstad , E. and Burnham , K.P. 1991 . User’s Guide for Interactive Program CAPTURE. Abundance Estimation of Closed Animal Populations , Fort Collins , CO : Colorado State University .
- Williams , A.K. and Berkson , J. 2004 . Reducing false absences in survey data: detection probabilities of Red‐backed Salamanders . J. Wildlife Manage , 68 : 418 – 428 .
- Williams , B.K. , Nichols , J.D. and Conroy , M.J. 2002 . Analysis and Management of Animal Populations , San Diego , CA : Academic Press .