Abstract
Capsule Eurasian Nuthatches have experienced a northward range expansion in Britain. Younger, more isolated, populations have smaller song meme-pool size, diversity and individual repertoires.
Evolution is the process whereby frequencies of variants in a population change over time due to differential propagation; this definition can apply to both genetic and cultural units (Bentley et al. Citation2004). Genes are propagated via reproduction while certain cultural traits (e.g. behaviours) can be propagated via imitation. Their cultural units are often called ‘memes’ (Dawkins Citation1976).
Evidence suggests that song-types can be transmitted between birds via imitation, and hence they can act as memes (Blackmore Citation1998). For example, Jenkins Citation(1978) found that young male South Island Saddlebacks Philesturnus carunculatus settled some distance from their father and their eventual repertoire only included song-types present in neighbouring males. Song meme pools can lose variation through similar phenomena to gene pools such as founder effects (e.g. Chaffinches Fringilla coelebs: Baker & Jenkins Citation1987; Singing Honeyeaters Lichenostomus virescens: Baker Citation1996) or random drift (e.g. Indigo Buntings Passerina cyanae: Payne et al. Citation1988).
Eurasian Nuthatches Sitta europaea (hereafter ‘Nuthatches’) are oscine passerines (Passeriformes, Passeri) inhabiting deciduous and mixed woodland throughout Europe and Asia (Cramp & Perrins Citation1993). Five distinct song-types (consistent, at least, in Britain, Belgium and Germany) have been described within Nuthatch populations which have been descriptively named ‘slow-ascending’ (SA), ‘fast-ascending’ (FA), ‘descending’ (DE), ‘trill’ (TR) and ‘up–down’ (UD) (Cramp & Perrins Citation1993, Matthysen Citation1997). Each consists of a short, uniform series of notes called ‘strophes’. Strophes of one song-type are repeated in loose ‘bouts’ of up to a minute. Males develop a repertoire of up to five song-types (Matthysen Citation1997). They fulfil Lynch's Citation(1996) description of discontinuous memes as they are discrete with high inter-observer reliability. Nuthatches are monogamous with contiguous, year-round, sharp-boundaried territories (Cramp & Perrins Citation1993).
Matthysen Citation(1997) and Matthysen et al. Citation(2002) studied geographic variation in Nuthatch song-types across Belgium. The song type UD was missing from two populations, one which was thought to have spread from a small isolated population and another that was the most recently to have colonized, and a founder effect was hypothesized. Memes must be defined by imitation (Blackmore Citation1998) and oscine song has been widely observed to be learnt, although precise structural and functional descriptions of song learning programmes are a matter for debate (Marler Citation1997, Beecher & Brenowitz Citation2005). Direct experimental evidence for song learning in Nuthatches appears to be unavailable, but patterns seen in studies suggest the songs are learnt. For example, Matthysen et al. Citation(2002) suggested that a missing song-type UD could not re-establish because of a lack of conspecifics from whom to learn it.
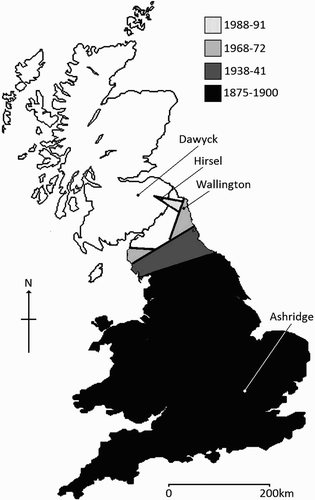
In Britain, Nuthatches were virtually absent north of the English county of Yorkshire (>100 km south of Scotland) between 1875 and 1900 (Holloway Citation1996), a pattern that seemed not to change significantly through the early and mid-20th century (Sharrock Citation1976). The late 20th and early 21st centuries, however, have seen a pronounced northward expansion ().
Figure 1. Locations of study sites in Great Britain in relation to Nuthatch colonization history since 1875. Shaded areas represent approximate maximal range during different periods, substantially simplified for illustrative purposes. Range information for the years labelled is taken from Holloway Citation(1996), Sharrock Citation(1976) and Gibbons et al. Citation(1993).
Nuthatches shun open habitats and populations inhabit ‘islands’ of woodland (González-Varo et al. Citation2007), particularly so in Britain, one of the least forested countries in Europe (Forestry Commission Citation2011). The pattern of expansion appears to take the form of appearances of isolated individuals away from established populations, followed by local increases and outward expansion to surrounding areas (Murray Citation2009), and so on northwards. For example, in Northumbria (colonized 1968) there is evidence that the species is still infilling suitable habitat (T. Dean, Northumbria county bird recorder, pers. comm.).
STUDY SITES AND HYPOTHESIS
Four study sites (, ) were selected to investigate how song meme-pool characteristics varied across this colonization zone. Given that meme-pool size decreased in newly colonized nuthatch populations in Belgium (Matthysen Citation1997, Matthysen et al. Citation2002), I hypothesized that populations nearer the colonization front, which are expected to be increasingly isolated and have smaller population sizes, would have lower song meme-pool size, diversity and individual repertoire sizes than older more established populations.
Table 1. The four study site locations, length of time that they have been occupied by Nuthatches (age), the density of tetrads which were occupied by Nuthatches within a 10 km square in which the study site was located (occupied tetrads/10 km2).The characteristics of the song-type meme pool at each site are indicated by the frequency of occurrence of the five song types (SA, TR, FA, DE, UD), the meme-pool size (S), meme-pool diversity (D) and mean repertoire size (S R ). n is the number of males whose songs were studied at each site.
COLONIZATION BY NUTHATCHES
A minimum population age for each site was estimated from the earliest published record of nuthatches at that site (). Population (thus meme pool) continuity since that record was confirmed with reference to annual county bird reports (these are not individually referenced for brevity, but were accessed at the RSPB Library, Sandy, UK in 2005) and breeding atlases (Sharrock Citation1976, Gibbons et al. Citation1993, Murray et al. Citation1998).
For Dawyck, Hirsel and Wallington, the first record is likely to be precise because their colonization was specifically noted within bird reports, so population age for these is assumed accurate. However, it is very likely that the population in Ashridge, an area of semi-natural ancient woodland in the core of the species' range (Gibbons et al. Citation1993), is older, but records prior to the earliest found (Goldingham Citation1985) are unavailable. The most recent British breeding bird atlas (Gibbons et al. Citation1993) recorded the presence/absence of species within each of a grid of tetrads (2 × 2 km squares) across the country. I used local tetrad density (occupied tetrads/10 km2) as a proxy for the degree of population isolation at the four sites, assuming populations would be less connected and have lower population sizes in areas of lower tetrad occupation. It is possible that tetrad occupancy has changed since the atlas surveys (1988–1991), but these are the most reliable indicator of relative geographical abundance closest in time to the study (2005).
DETERMINATION OF SONG MEME-POOL SIZE
Territory-mapping (Bibby et al. Citation1992) was used to ensure that each subject was a separate individual. All song-types heard by each male during territory-mapping were noted. During a pilot study in February 2005 at a separate site, three visits made on consecutive dry mornings were found adequate to identify individuals and estimate broad territory boundaries.
I used playback (prompting song from a subject by playing it recordings of conspecifics: McGregor Citation2000) to estimate individual repertoires, as Matthysen et al. Citation(2002) found repertoires can be underestimated by only listening for spontaneous song. Two examples of strophes of each song-type (Matthysen et al. Citation2002) were sourced from high-quality recordings (Kettle & Ranft Citation1992, Roche Citation1996, Roche & Chevereau Citation2001). The sequence used consisted of ten strophes of each song-type (both examples alternated five times) played over 1 minute, followed by 1 minute of silence. This was repeated for each song-type. The order of the five song-types was randomized for each bird (Matthysen et al. Citation2002). Then any further responses were recorded for 11 minutes. Strophes of each song-type heard during the 20-minute session were counted.
Playback was carried out between 1 and 30 March 2005 in an arbitrary site order (Ashridge – Hirsel – Dawyck – Wallington) between 09:00 and 17:00 hours GMT/BST. Nuthatch songs are heard relatively constantly from the male December–June and are generally heard throughout the day, lacking dawn/dusk peaks (Matthysen Citation1998), so date and time effects were not controlled for. A constant playback volume was selected during the pilot study to be consistent with males singing within about 20 m.
Twelve playback males from each site were selected at random from those territory-mapped, although only seven males could be located within the area of permitted access at Dawyck, so all were sampled. A Sony TCM 939 cassette player was attached 1.5 m high on a tree trunk at approximately the centre of a bird's mapped territory and playback sessions were started when the male was within about 20 m of the player (possible because males traverse their territories relatively conspicuously). The observer was camouflaged and remained by a tree ≥15 m away. Playback was only carried out in calm, dry conditions (see Catchpole & Slater Citation1995) and only once on each male to minimize disruption of natural territorial behaviour.
For each site meme pool, size (S) was taken as the total number of song-types heard from all sampled males and mean repertoires size (S R ) as the average number of songs for all sampled males. I used a Simpson's index (D) of song-type diversity (Lynch Citation1996) in which n = total number of strophes heard during playback at a site and Pi = mean proportion of strophes sung by each male of the ith song-type:
Males responded actively to playback, most approaching to within 10–20 m and making ‘excitement’ calls. All 43 sang in response, 36 beginning during the first playback song-type, 6 the second and 1 the fourth. No ambiguous or previously undescribed song-types were heard.
It was clear from comparison of playback and spontaneous song (heard during territory-mapping) that playback was not fully effective in elucidating all song-types from repertoires. In 13/43 males one additional song-type (although never more than one) was heard during territory-mapping but not playback. Six of these were SA, two TR, three FA, one DE and one UD. It is unclear why SA in particular was apparently not responded to more than the other song types. One possibility might be that the SA stimulus recordings used happen to be characteristically unusual compared to the study populations. However, this was not detectable to the author when comparing them to response or spontaneous SA strophes heard from the subjects. Another possibility is that the recordings used were of insufficient quality. The recordings were made professionally, and Nuthatch songs are in low frequency bands, making them robust to minor distortion, so playback using even relatively poor recordings is usually possible (E. Matthysen, University of Antwerp, pers. comm.), so this seems unlikely.
As playback alone underestimated repertoire sizes in this study (cf. Matthysen et al. Citation2002), I estimated S and S R from songs heard during both territory-mapping and playback combined. Although the duration of territory-mapping of each subject was not strictly standardized, each male had a relatively similar period of exposure to the observer (i.e. three mornings of territory-mapping observation). Unfortunately, I could not utilize spontaneous song heard during territory mapping in the calculation of D for each site because the actual number of strophes heard of each song-type was not noted, and this measure is required in its estimation. Nevertheless, it may at least provide a relative measure of diversity between the sites.
SONG TYPES AND MEME-POOL SIZE
All five song-types were detected at each site except Dawyck (). Here UD was absent, the rarest song-type heard elsewhere (; note, UD was not heard during playback at Hirsel, but was heard once produced by a single male during territory-mapping so is included in S). As the number of males sampled at Dawyck was lower (7 compared to 12 at other sites) it cannot be ruled out that the lack of UD was a sampling artefact, and it is possible that it is present within the population but at a low frequency such that it was not elucidated from playback (as at Hirsel) or heard during territory-mapping.
Figure 2. Meme-pool characteristics from playback: (a) mean proportion of song-types in repertoires of males at each site; (b) meme-pool diversity (D) in relation to the natural logarithm of local tetrad density (tetrads occupied by breeding birds per 10 km2) with regression line (r 2 = 0.99, F 1,2 = 497.92, P = 0.002).
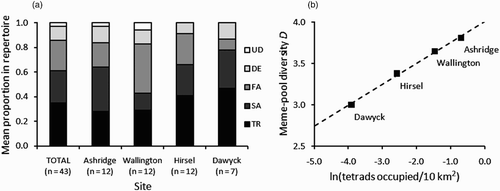
The proportions of song-types heard during playback, and the dominant song-type, varied between sites (. Both D and S R decreased in younger, more isolated populations (), although in univariate linear regression models both had lower P-values with tetrad density (D: r 2 = 0.99, F 1,2 = 497.92, P = 0.002; S R : r 2 = 0.93, F 1,2 = 25.60, P = 0.037) (the former illustrated in than with minimum population age (D: r 2 = 0.90, F 1,2 = 18.44, P = 0.050; S R : r 2 = 0.85, F 1,2 = 11.08, P = 0.080).
SONG PATTERNS
The patterns observed can potentially be explained by memetic processes occurring across the colonization zone. Although potentially an artefact of sampling fewer males, the apparent lack of UD at Dawyck would be consistent with a founder effect. UD has a low frequency of occurrence both within repertoires ( and proportion of males possessing that song (8–25%: ) in other populations, which would reduce the probability of possession by immigrants. Even if the UD had been present in founders, the extinction probability of a song meme can be linked to its meme-pool frequency (Payne et al. Citation1988) so the death of a single or small group of birds could have by chance extirpated it. Interestingly, it is the same song-type probably lost through founder effects in two Belgian populations (Matthysen et al. Citation2002).
Founder effects in song memes have been found in a number of populations, particularly where an island or distant suitable habitat patch has been colonized. For example, reduced meme-pool characteristics have been observed in founder populations of Singing Honeyeaters (Baker Citation1996), Eurasian Treecreepers Certhia familiaris (Baptista & Johnson Citation1982), and Fox Sparrows Passerella iliaca (Naugler & Smith Citation1991; although see Martin Citation1993 and Naugler & Smith Citation1993 for discussion on the latter finding). The UK Nuthatch population history may differ somewhat from a discrete population founder event where population age will increase but the level of isolation may remain relatively constant (for example, Chaffinches colonizing an island off New Zealand: Baker & Jenkins Citation1987). The continued northward range expansion by Nuthatches means both isolation and age of populations across it are likely to vary clinally (demonstrated here) and to change over time (isolation decreasing, age increasing). I was unable to distinguish either of these confounding variables as the key driver, or whether they interact, although there was a slightly stronger univariate relationship between local tetrad density and D and S R .
The pattern seen by Matthysen Citation(1997) suggested that a founder event can cause a change in meme frequencies (i.e. loss of a song-type) that stays relatively constant over time where a population expands but remains highly isolated. Yet in the case of Nuthatch range expansion in Britain, new populations are likely to be reinforced relatively quickly by new immigrants or join with other expanding local populations, which would be expected to diminish differences. For example, Wallington was colonized in 1970 as a relatively isolated population, but is now well within the species' range ().
CLINAL VARIATIONS IN MEME-POOL SIZE
So why is such clinal variation apparent? At least two processes, which can cause positive feedback on the propagation of song-types in accordance to their frequency, might further explain the pattern of reduced song diversity (D) and repertoire size (S R ) in smaller, more isolated populations. ‘Majority-type advantage’ is the process whereby common variants are copied disproportionately to their availability (Lynch Citation1996). For example, young Song Sparrows Melospiza melodia preferentially learnt song-types shared by many neighbours over those used by one or two neighbours (Beecher et al. Citation1994). In addition, ‘selective attrition’ occurs where males drop songs from their repertoires that do not match those of their neighbours (Nordby et al. Citation2007). Nuthatches can employ song-matching (replying to a competitor with the same song-type; Matthysen et al. Citation2002) and song-types possessed by immigrants but not the host population might be dropped (not expressed and then lost through mortality) through lack of response, reinforcing their absence/rarity even with further immigration of that song-type.
A fall in S R could be a function of reduced S, meaning a lower selection of songs from which to build a repertoire, or reduced D, meaning less chance of exposure to rarer song-types. However, it could also be a result of lower local territory densities. Bitterbaum & Baptista Citation(1979) suggested that having fewer conspecifics with which to interact, due to decreased population densities, was the reason behind a founder population of introduced House Finches Carpodacus mexicanus having a lower mean repertoire size than the parent population. Matthysen et al. Citation(2002) also found that repertoire size was higher in territorial Nuthatches with more neighbours. As study sights were not fully searched for all territories, I cannot compare repertoire size with the number of neighbours, but birds at Dawyck could potentially have had fewer neighbours, as only seven territories could be located in the area studied.
Could differences in song meme pools ( be explained by memetic selection pressures, whereby certain song-types were differentially imitated depending on their ability to be copied? For example, this might occur via differential degradation through vegetation (see Podos & Warren Citation2007) or social factors (e.g. imitation of call-memes of Black-capped Chickadees Poecile atricapillus may be linked to bird dominance: Baker & Gammon Citation2008). Across the current colonization zone it is not possible to give a reason why some song-types such as UD might be selected against. Additionally, some studies have suggested that song memes are often functionally equivalent and selectively neutral so their relative frequencies are a result of chance events such as drift, rather than selection (see Lynch Citation1996).
CONCLUSIONS
This was an observational study with relatively small sample sizes of sites and birds. There were some limitations to the data, as playback techniques underestimated individual repertoires and sample size was lower at one site. However, interesting clinal variation in meme-pool characteristics was observed in that song meme-pool diversity and song-repertoire sizes all fell in younger, more isolated populations. In the youngest population at the colonization ‘front’ the song-type least frequent in the other populations was either possibly locally extinct or extremely rare. This general pattern concurs with that seen in Belgium (Matthysen Citation1997, Matthysen et al. Citation2002).
Given the characteristics of Nuthatch song-types, memetic drift and/or founder effects, compounded by majority-type advantage and/or selective attrition might explain the observed patterns. However, elucidating the precise mechanisms behind patterns observed might be achieved via monitoring in real time the colonization of new patches. Marking of territorial males and recording on an annual basis the repertoires of these and new males could allow observation of whether reduced meme pools exist at early colonization, if this pattern remains stable over time, if other song-types are introduced (via immigration or perhaps mutation) and how the frequencies of song-types vary over time. The expansion of Nuthatches in Britain has continued since this study (Toms Citation2009). Detailed recordings made by volunteers and bird recorders could be used to initially identify new patches containing individual or small numbers of new birds but with potential for local population expansion. Laboratory/aviary studies, or the manipulation of local population densities via translocation, could also help to further test the effect of number of neighbours (and number of neighbouring song-type memes) on the developing repertoires of young birds and the precise song learning programme (Beecher & Brenowitz Citation2005) of Nuthatches.
ACKNOWLEDGEMENTS
This work was funded privately and by the University of Edinburgh. Many thanks to Peter Jones, Eric Matthysen, Kate Heal, Richard Ennos, Beccy Coe, Ray Murray, Tim Dean, Gemma Jenkins and Hayley Douglas. Field access was kindly granted by the National Trust, Roger Dodd & Co., the Douglas-Home family, and the Royal Botanic Garden Edinburgh. Thanks to the RSPB for library access. Ken Norris, Dave Gammon and one anonymous reviewer provided helpful comments on an earlier draft.
REFERENCES
- Baker , A. J. and Jenkins , P. F. 1987 . Founder effect and cultural evolution of songs in an isolated population of chaffinches, Fringilla coelebs, in the Chatham Islands . Anim. Behav. , 35 : 1793 – 1803 .
- Baker , M. C. 1996 . Depauperate meme pool of vocal signals in an island population of singing honeyeaters . Anim. Behav. , 51 : 853 – 858 .
- Baker , M. C. and Gammon , D. E. 2008 . Vocal memes in natural populations of chickadees: why do some memes persist and others go extinct? . Anim. Behav. , 75 : 279 – 289 .
- Baptista , L. F. and Johnson , R. B. 1982 . Song variation in insular and mainland California Brown Creepers (Certhia familiaris) . J. Ornithol. , 123 : 131 – 144 .
- Beecher , M. D. and Brenowitz , E. A. 2005 . Functional aspects of song learning in songbirds . Trends Ecol. Evol. , 20 : 143 – 149 .
- Beecher , M. D. , Campbell , S. E. and Stoddard , P. K. 1994 . Correlation of song learning and territory establishment strategies in the song sparrow . Proc. Natl Acad. Sci. USA , 91 : 1450 – 1454 .
- Bentley , R. A. , Hahn , M. W. and Shennan , S. J. 2004 . Random drift and culture change . Proc. R. Soc. London B , 271 : 1443 – 1450 .
- Bibby , C. J. , Burgess , N. D. and Hill , D. A. 1992 . Bird Census Techniques , London : Academic Press .
- Bitterbaum , E. and Baptista , L. F. 1979 . Geographical variation in songs of California house finches (Carpodacus mexicanus) . Auk , 96 : 462 – 474 .
- Blackmore , S. 1998 . Imitation and the definition of a meme . J. Memet. , 2: [online only: http://jom-emit.cfpm.org/1998/vol2/balckmore_s.html]
- Catchpole , C. K. and Slater , P. J.B. 1995 . Bird Song: Biological Themes and Variation , Cambridge : Cambridge University Press .
- Cramp , S. and Perrins , C. M. 1993 . Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic, Vol. VII: Flycatchers to Shrikes , Oxford : Oxford University Press .
- Dawkins , R. 1976 . The Selfish Gene , Oxford : Oxford University Press .
- Forestry Commission . 2011 . Woodland Area, Planting and Restocking , London : UK National Statistics .
- Gibbons , D. W. , Reid , J. B. and Chapman , R. A . 1993 . The New Atlas of Breeding Birds in Britain and Ireland 1998–1991 , Edited by: Gibbons , D. W. , Reid , J. B. and Chapman , R.A . London : T & AD Poyser .
- Goldingham , A . 1985 . British Birds from Nature: The Tresco Collection of Early Victorian Paintings and Conservations by Frances Mary Isabella Smith , Edited by: Goldingham , A . London : Hamlyn .
- González-Varo , J. P. , López-Bao , J. V. and Guitián , J. 2007 . Presence and abundance of the Eurasian nuthatch Sitta europaea in relation to the size, isolation and the intensity of management of chestnut woodlands in the NW Iberian Peninsula . Landscape Ecol. , 23 : 79 – 89 .
- Holloway , S. 1996 . The Historical Atlas of Breeding Birds in Britain & Ireland 1875–1900 , London : T & AD Poyser .
- Jenkins , P. F. 1978 . Cultural transmission of song patterns and dialect development in a free-living bird population . Anim. Behav. , 26 : 50 – 78 .
- Kerr , I. 2001 . Northumbrian Birds , Newcastle : Northumberland & Tyneside Bird Club .
- Kettle , R. and Ranft , R . 1992 . British Bird Sounds on CD , Edited by: Kettle , R. and Ranft , R . London : The British Library Nation Sound Archive .
- Lynch , A. 1996 . “ The population memetics of birdsong ” . In Ecology and Evolution of Acoustic Communication in Birds , Edited by: Kroodsma , D. E. and Miller , E. H. Ithaca , NY : Cornell University Press .
- Marler , P. 1997 . Three models of song learning: evidence from behaviour . J. Neurobiol. , 33 : 501 – 516 .
- Martin , D. J. 1993 . Song similarity in populations of fox sparrows: a rejection of Naugler's and Smith's conclusions . Condor , 95 : 1057 – 1059 .
- Matthysen , E. 1997 . Geographic variation in the occurrence of song types in Nuthatch Sitta europaea populations . Ibis , 139 : 102 – 106 .
- Matthysen , E. 1998 . The Nuthatches , London : T & AD Poyser .
- Matthysen , E. , Devos , Y. and Hepp , E. 2002 . Loss of song variation through cultural drift in Nuthatches Sitta europaea: testing an hypothesis . Avian Science , 2 : 1 – 9 .
- McGregor , P. K. 2000 . Playback experiments: design and analysis . Acta Ethol. , 3 : 3 – 8 .
- Murray , R. D. 1991 . The first successful breeding of nuthatch in Scotland . Scottish Bird Report , 22 : 51 – 55 .
- Murray , R. D. 1998 . Borders Bird Report, No. 18 , Aberlady : Scottish Ornithologists' Club .
- Murray , R. D. 2009 . Borders Bird Report, No. 29 , Aberlady : Scottish Ornithologists' Club .
- Murray , R. D. , Holling , R. D. , Dott , H. E.M. and Vandome , P. 1998 . The Breeding Birds of South-east Scotland: A Tetrad Atlas 1988–1994 , Edinburgh : Scottish Ornithologists' Club .
- Naugler , C. T. and Smith , P. C. 1991 . Song similarity in an isolated population of Fox Sparrows (Passerella iliaca) . Condor , 93 : 1001 – 1003 .
- Naugler , C. T. and Smith , P. C. 1993 . Similarity breeds confusion: a reply to Martin . Condor , 95 : 1059 – 1060 .
- Nordby , J. C. , Campbell , S. E. and Beecher , M. D. 2007 . Selective attrition and individual song repertoire development in song sparrows . Anim. Behav. , 74 : 1413 – 1418 .
- Payne , R. B. , Payne , L. L. and Doehlert , S. M. 1988 . Biological and cultural success of song memes in indigo buntings . Ecology , 69 : 104 – 117 .
- Podos , J. and Warren , P. S. 2007 . The evolution of genetic variation in birdsong . Adv. Stud. Behav. , 37 : 403 – 458 .
- R Core Development Team . 2009 . R Version 2.10.0 , Vienna : R Foundation for Statistical Computing .
- Roche , J. C . 1996 . Bird Songs and Calls of Britain and Europe 4: Sylvia Warblers to Bunting [CD] , Edited by: Roche , J.C . Salthouse : Wildsounds .
- Roche , J. C. and Chevereau , J . 2001 . [Guia Sonora de las Aves de Europa] [CD] , Edited by: Roche , J. C. and Chevereau , J . Barcelona [in Spanish] : Lynx Editions .
- Sharrock , J. T.R. 1976 . The Atlas of Breeding Birds in Britain and Ireland , Calton : T & AD Poyser .
- Toms , M. 2009 . The long road north . Bird Table , 57 : 8 – 11 .