Abstract
Capsule Amount of sparse vegetation show a direct link to reproductive success.
Aims To examine the influence of sparse vegetation on clutch size of Common Redstarts Phoenicurus phoenicurus.
Methods Clutch size and the amount of sparse ground vegetation within territories were measured.
Results Clutch size was positively correlated with the amount of sparse vegetation. Clutches in territories with the highest amounts of sparse vegetation contained approximately one more egg than clutches in territories with the lowest amount of sparse vegetation.
Conclusion The presence of sparse vegetation is correlated with clutch size and thus reproductive success. This finding suggests a direct link between habitat degradation through the loss of sparse vegetation and observed population decreases of bare-ground foraging birds. The results strengthen the argument that the implementation of sparse vegetation in agri-environment schemes is likely to improve the breeding success of bare-ground foraging birds, such as Common Redstarts.
Intensification of farmland management has had dramatic and widespread negative influence on farmland biodiversity since the middle of the last century (Robinson & Sutherland Citation2002, Green et al. Citation2005, Klejin et al. Citation2009). For example, the populations of several farmland birds have shown marked and ongoing declines in many European countries (Fuller et al. Citation1995, Donald et al. Citation2006). To counteract the decline of farmland species, several countries have implemented agri-environment schemes (Kleijn et al. 2004). Unfortunately, the agri-environment schemes have so far only moderately supported biodiversity increases, possibly because they are only rarely targeting the specific needs of species of conservation concern (Kleijn et al. Citation2001, Perkins et al. Citation2011).
Several ground-feeding farmland birds inhabiting grassland dominated landscapes need bare ground, sparse vegetation and short stubble fields (hereafter called sparse vegetation) for in which they forage (Vickery et al. Citation2001, Butler & Gillings Citation2004, Atkinson et al. Citation2005, Schaub et al. Citation2010). Denser and more homogenous swards reduce the detectability and accessibility of prey leading to a decrease of habitat quality for insectivorous birds (MacCracken & Tallowin 2004). Furthermore, a denser sward structure increases the risk of predation (Whittingham & Evans Citation2004). Yet agricultural intensification has tended to result in denser and more homogenous swards (Benton et al. Citation2003) and a lot of measures promoted by agri-environment schemes, e.g. grassy ecological compensation areas, low-intensity and extensive meadows, set-aside land or wildflower areas, are not characterized by sparse vegetation and bare ground. Therefore it has been suggested that the improvement of agri-environment schemes requires the implementation of measures to increase the amount of sparse vegetation in grasslands (Schaub et al. Citation2010).
To date, the benefit of sparse vegetation on biodiversity has been shown using studies on the behaviour of individuals in relation to the availability of sparse vegetation. For instance, radiotracked Eurasian Hoopoes Upupa epops predominantly use sparse vegetation for foraging (Weisshaupt et al. Citation2011). Or in a habitat choice experiment in aviaries, Common Redstarts Phoenicurus phoenicurus caught mealworms almost exclusively in sparse vegetation when mealworms were provided in both sparse vegetation and dense meadow at the same time (Martinez et al. Citation2010). However, whether these behavioural responses to sparse vegetation are relevant for the demography of the population is far from clear. For example, Schön Citation(2011) suggests that naturally occurring patches of sparse vegetation might be a pitfall for Eurasian Skylarks, with high attractiveness yet low breeding success. Therefore, if a direct link between sparse vegetation and a demographic parameter could be demonstrated, this would strengthen the arguments for implementing measures designed to enhance the amount of sparse vegetation in agri-environment schemes.
Here I examined the influence of sparse vegetation on clutch size in Common Redstarts. I define sparse vegetation as ground vegetation with permanent limited or little vegetation and parts of bare ground, thus structures where sit-and wait predators such as Common Redstarts can easily detect and pick up prey items. In Central Europe Common Redstarts inhabit mainly high-stem orchards and gardens (Schmid et al. Citation1998) and they have suffered a steep decline in many European countries during the second half of the last century (Bruderer & Hirschi Citation1984, BirdLife International Citation2004). Thus, the Common Redstart is one of 50 priority bird species for recovery programmes in Switzerland (Keller et al. Citation2010). Former studies have shown that Common Redstarts prefer territories with a high amount of sparse vegetation, and that they mainly forage on such areas, and even prefer sparse vegetation for foraging when prey biomass is much lower in sparse vegetation than in surrounding dense meadow (Martinez et al. Citation2010, Schaub et al. Citation2010). However, it remains unclear if the amount of sparse vegetation directly affects reproductive success. In the present study I used clutch size as an indicator of reproductive success and tested whether the amount of sparse vegetation in a territory is associated with clutch size.
METHODS
Study site and field methods
The study was conducted during the breeding seasons 2010 and 2011 in northwestern Switzerland. The study site covered a mosaic of orchards, vineyards and gardens in mostly intensively managed grasslands. Within the study site I surveyed a population of about 40 pairs of Common Redstarts, where about one-third of the pairs were breeding in natural cavities and the other two-thirds usually breeding in nestboxes. During several visits from mid-April onwards each potential territory was visited at least three times. If a territory was occupied, I searched for the nesting site of the pair by checking all the nestboxes and cavities known from previous breeding seasons. If the pair did not nest in one of these sites, the adults were observed to obtain hints about the location of their nesting site. From the 41 and 40 occupied territories in 2010 and 2011, respectively, the nesting sites in 73 % and 55 % of the occupied territories were found. The located nests were then visited every second day until the female started to incubate and at this point clutch size was determined. Nestboxes and cavities which could not be easily examined were monitored with the help of a small torch light and a laryngeal mirror. However, there were still several nests (17 nests and 5 nests in 2010 and 2011, respectively) where assessment of clutch size was not possible. The number of eggs at the start of breeding was used as a measure of reproductive success in the analyses. I chose clutch size as an indicator of reproductive success as it is an oft-used factor for the determination of reproductive success (Högstedt Citation1980, Christians Citation2002).
Between May and July, I mapped the habitats of all the Redstart territories with known nesting site on a circular plot with radius 50 m (i.e. 7850 m2) around the nesting site. The size of the plots corresponded to the size of a Redstart territory that usually covers an area between 1400 m2 (Menzel Citation1971) up to 10000 m2 (Glutz von Blotzheim Citation1988). Especially when feeding, young Common Redstarts tend to forage closely around the nest (own unpublished data) and usually stay in close distance (< 50 m) to the nest. Therefore, I assume that most nests were more or less centrally placed within territories and that the size and circular shape of the investigated plot was reasonable. For each plot I mapped roads, trees and all occurring vegetation types. The percentage of each vegetation type in the circular ‘territory’ was then determined with a grid of 2500 points. For further analysis all occurring vegetation types were grouped into two habitat types (sparse vegetation and ‘other’, ). The percentage of the points that were assigned as sparse vegetation was then used as the measure of habitat quality. The same method was applied in an earlier study on Common Redstarts (Martinez et al. Citation2010).
Table 1. The 13 vegetation types that were considered as sparse vegetation and the 7 vegetation types that were considered as ‘other’. *Forest vegetation was mainly represented by Fagus silvatica dominated woods with open ground, which is the reason why it was considered as sparse vegetation.
Statistical analysis
In most passerine species, second broods or replacement broods are in general smaller than first broods (Klomp 1970), and this is the case in Common Redstarts (Glutz von Blotzheim Citation1988). Therefore, I only included broods with breeding start date before 15 May in the analysis. To assess the relation of habitat quality (i.e. the percentage of sparse vegetation in a territory) to clutch size, I used a linear regression model with the number of eggs as the dependent variable and the percentage of sparse vegetation as predictor variable. I controlled for the differences between years by also including the factor ‘year’ into the model. The visual check of the residuals from the linear model did not show any signs of violation of model assumptions. All analyses were done using the statistical software r (R Development Core Team Citation2011).
RESULTS
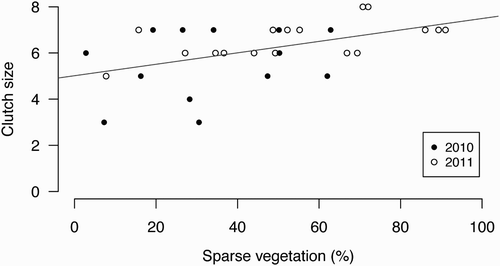
Sparse vegetation cover in territories ranged from 2.80 % up to 91.02 % (mean ± se = 45.04 % ± 4.48, n = 30). Common Redstart pairs in territories with a high proportion of sparse vegetation had higher clutch sizes than pairs in territories with a low proportion of sparse vegetation (linear regression: slope = 0.019, t = 2.123, P = 0.043, ). Average (± se) clutch size was 5.54 ± 0.42 eggs (n = 13) in 2010 and 6.59 ± 0.19 eggs (n = 17) in 2011. The difference in clutch size between year was not significant (linear regression: effect of second year = 0.654, t = 1.479, P = 0.151).
Figure 1. Regression of clutch size on the percentage of sparse vegetation in Common Redstart territories. Pairs in territories with high proportion of sparse vegetation had higher clutch sizes than pairs in territories with low proportion of sparse vegetation. Black dots: data from 2010 (n = 13), open dots: data from 2011 (n = 17), grey line: linear regression line. Note that one dot from 2010 (clutch size: 6, sparse vegetation: 49 %) overlaps with a dot from 2011.
DISCUSSION
Clutch size of Common Redstarts increased with increasing amount of sparse vegetation in the territories (). The average clutch size in territories with the lowest amount of sparse vegetation was about one egg smaller than the clutch size in territories with the highest amount of sparse vegetation. Therefore, if sparse vegetation could be increased in otherwise suitable habitat, the population size of the Common Redstart is likely to increase. Several other declining European farmland birds, e.g. Hoopoes, Woodlarks Lullula arborea and Eurasian Wrynecks Jynx torquilla, also prefer sparse vegetation for foraging (Schaub et al. Citation2010, Arlettaz et al. Citation2011, Weisshaupt et al. Citation2011). It is probable that similar effects of sparse vegetation as described for Common Redstarts play a major role in their ecology and that their populations would also benefit from an increase of sparse vegetation in their habitats.
Two non-exclusive explanations may account for the correlation between the amount of sparse vegetation and clutch size in Common Redstarts. First, females may adjust their clutch size on the basis of territory quality and tend to lay more eggs in better territories. Females that adapt clutch size to environmental factors have been observed in several other passerine species (Högstedt Citation1980, Hussell & Quinney Citation1985). My results suggest that this is also the case in Common Redstarts, with females adapting clutch size to the amount of sparse vegetation in their territories. However, other factors that are correlated with the amount of sparse vegetation could explain my results. Nevertheless, as Common Redstarts prefer territories with a high amount of sparse vegetation and clearly prefer sparse vegetation for foraging (Martinez et al. Citation2010, Schaub et al. Citation2010) this explanation seems to be reasonable. Second, high-quality individuals may get the better territories, i.e. territories with higher amounts of sparse vegetation, than low-quality individuals. As the individuals of the best quality may lay more eggs than low-quality individuals (Klomp Citation1970, Christians Citation2002), a correlation between the amount of sparse vegetation and the clutch size could also be expected.
In both explanations for the correlation between the amount of sparse vegetation and clutch size, the amount of sparse vegetation is a key predictor of the quality of Common Redstart territories. Therefore, if the amount of sparse vegetation could be increased in Common Redstart habitats the average clutch size of affected populations is likely to increase. Such an increase in the population clutch size is likely to result in higher reproductive success. Nevertheless, other aspects such as predation rates on chicks could be negatively correlated with the amounts of sparse vegetation in the territory, and thus higher clutch size does not definitely provide evidence that increasing the amount of sparse vegetation will indeed result in an increase of population size. In order to definitely prove a causal relationship between sparse vegetation and reproductive success, experimental studies should be conducted.
Overall, my results not only confirm the findings of earlier studies showing the importance of sparse vegetation for the foraging behaviour of Common Redstarts (Martinez et al. Citation2010, Schaub et al. Citation2010), but show that the presence of sparse vegetation is correlated with clutch size and thus with an important measure of reproductive success. The results therefore reveal a direct link between habitat degradation through the loss of sparse vegetation and observed population decreases.
Based on this study, I suggest that agri-environment schemes aimed at bare-ground foraging birds would be implemented by promoting sparse vegetation in grasslands. In order to provide evidence for a positive effect of sparse vegetation on population size of bare-ground foraging birds, the influence of ‘sparse vegetation management measures’ on populations of bare-ground foraging bird species should be monitored.
ACKNOWLEDGEMENTS
I thank T. Roth from Hintermann & Weber AG for crucial help with statistical analyses and for important comments on an earlier draft of the article, two anonymous reviewers for their valuable comments, as well as S. Hahn, J. Laesser, H. von Hirschheydt and N. Zbinden from the Swiss Ornithological Institute for useful advices concerning study design and field work and for providing nestboxes. Furthermore, I thank the Ornithological Society of Basel for funding the material of the entire study.
REFERENCES
- Atkinson , P. W. , Fuller , R. J. , Vickery , J. A. , Conway , G. J. , Tallowin , J. R.B. , Smith , R. E.N. , Haysom , K. A. , Ings , T. C. , Asteraki , E. J. and Brown , V. K. 2005 . Influence of agricultural management, sward structure and food resources on grassland field use by birds in lowland England . J. Applied Ecol. , 42 : 932 – 942 .
- Arlettaz , R. , Maurer , M. L. , Mosimann-Kampe , P. , Nusslé , S. , Abadi , F. , Braunisch , V. and Schaub , M. 2011 . “ New vineyard cultivation practices create patchy ground vegetation, favouring Woodlarks ” . In J. Ornithol Online First. DOI: 10.1007/s10336-011-0737-7
- Benton , T. G. , Vickery , J. A. and Wilson , J. D. 2003 . Farmland biodiversity: is habitat heterogeneity the key? . Trends in Ecol. Evol. , 18 : 182 – 188 .
- BirdLife International . 2004 . “ Species factsheet: Phoenicurus phoenicurus ” . Available from http://www.birdlife.org (accessed September 2011)
- Bruderer , B. and Hirschi , W. 1984 . Langfristige Bestandsentwicklung von Gartenrötel Phoenicurus phoenicurus und Trauerschnäpper Ficedula hypoleuca nach schweizerischen Beringungszahlen und Nisthöhlenkontrollen . Der Ornithologische Beobachter , 81 : 285 – 302 .
- Butler , S. J. and Gillings , S. 2004 . Quantifying the effects of habitat structure on prey detectability and accessibility to farmland birds . Ibis , 146 ( Suppl. 2 ) : 123 – 130 .
- Christians , J. K. 2002 . Avian egg size: variation within species and inflexibility within individuals . Biol. Rev. , 77 : 1 – 26 .
- Donald , P. F. , Sanderson , F. J. , Burfield , I. J. and van Bommel , F. P.J. 2006 . Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000 . Agric. Ecosyst. Environ. , 116 : 189 – 196 .
- Fuller , R. J. , Gregory , R. D. , Gibbons , D. W. , Marchant , J. H. , Wilson , J. D. , Baillie , S. R. and Carter , N. 1995 . Population declines and range contractions among lowland farmland birds in Britain . Conserv. Biol. , 9 : 1425 – 1441 .
- Glutz von Blotzheim , U. N. 1988 . Handbuch der Vögel Mitteleuropas Band 11/1: Gartenrotschwanz Phoenicurus phoenicurus: 303–389. Aula Verlag, Wiesbaden
- Green , R. E. , Cornell , S. J. , Scharlemann , J. P.W. and Balmford , A. 2005 . Farming and the fate of wild nature . Science , 307 : 550 – 555 .
- Högstedt , G. 1980 . Evolution of clutch size in birds: adaptive variation in relation to territory quality . Science , 210 : 1148 – 1150 .
- Hussell , D. J.T. and Quinney , T. E. 1985 . Food abundance and clutch size of Tree Swallows . Tachycineta biclor. Ibis. , 129 : 243 – 258 .
- Keller , V. , Ayé , R. , Müller , W. , Spaar , R. and Zbinden , N. 2010 . Die prioritären Vogelarten der Schweiz: Revision 2010 . Der Ornithologische Beobachter , 107 : 265 – 258 .
- Kleijn , D. , Berendse , F. , Smit , R. and Gilissen , N. 2001 . Agri-environment schemes do not effectively protect biodiversity in Dutch agricultural landscapes . Nature , 413 : 723 – 725 .
- Klejin , D. , Berendse , F. , Smit , R. , Gilissen , N. , Smit , J. , Brak , B. and Groeneveld , R. 2004 . Ecological effectiveness of agri-environment schemes in different agricultural landscapes in the Netherlands . Conserv. Biol. , 18 : 775 – 786 .
- Klejin , D. , Kohler , F. , Báldi , A. , Batáry , P. , Concepción , E. D. , Clough , Y. , Díaz , M. , Gabriel , D. , Holzschuh , A. , Knop , E. , Kovács , A. , Marshall , E. J.P. , Tscharntke , T. and Verhulst , J. 2009 . On the relationship between farmland biodiversity and land-use intensity in Europe . Proc. R. Soc. Lond. B. , 276 : 903 – 909 .
- Klomp , H. 1970 . The determination of clutch size in birds. A review . Ardea , 58 : 1 – 124 .
- Martinez , N. , Jenni , L. , Wyss , E. and Zbinden , N. 2010 . Habitat structure versus food abundance: the importance of sparse vegetation for the Common Redstart Phoenicurus phoenicurus . J. Ornithol. , 151 : 297 – 307 .
- McCracken , D. I. and Tallowin , J. R. 2004 . Swards and structure: the interactions between farming practises and bird food resources in lowland grassland . Ibis , 146 ( Suppl. 2 ) : 108 – 114 .
- Menzel , H. 1971 . “ Der Gartenrotschwanz Phoenicurus phoenicurus ” . Neue Brehm Bücherei Nr. 438, A. Ziemsen Verlag. Wittenberg Lutherstadt
- Perkins , A. J. , Maggs , H. E. , Watson , A. and Wilson , J. D. 2011 . Adaptive management and targeting of agri-environment schemes does benefit biodiversity: a case study of the corn bunting Emberiza calandra . J. Appl. Ecol. , 48 : 514 – 522 .
- R Development Core Team . 2011 . R: A Language and Environment for Statistical Computing R Foundation for Statistical Computing
- Robinson , R. A. and Sutherland , W. J. 2002 . Post-war changes in arable farming and biodiversity in Great Britain . J. Appl. Ecol. , 39 : 157 – 176 .
- Schaub , M. , Martinez , N. , Tagmann-Ioset , A. , Weisshaupt , N. , Maurer , M. L. , Reichlin , T. S. , Abadi , F. , Zbinden , N. , Jenni , L. and Arlettaz , R. 2010 . Patches of bare ground as a staple commodity for declining insectivorous farmland birds . PLoS ONE , 5 ( 10 ) DOI: 10.1371/journal.pone.0013115
- Schmid , H. , Luder , R. , Naef-Daenzer , B. , Graf , R. and Zbinden , N. 1998 . “ Schweizer Brutvogelatlas ” . Verbreitung der Brutvögel in der Schweiz und im Fürstentum Liechtenstein 1993–1996. Schweizerische Vogelwarte, Sempach
- Schön , M. 2011 . Long-lived sustainable microhabitat structures in arable ecosystems, and Syklarks (Alauda arvensis) . J. Nat. Conserv. , 19 : 143 – 147 .
- Vickery , J. A. , Tallowin , J. R. , Feber , R. E. , Asteraki , E. J. , Atkinson , P. W. , Fuller , R. J. and Brown , V. K. 2001 . The management of lowland neutral grasslands in Britain: effects of agricultural practices on birds and their food resources . J. Appl. Ecol. , 38 : 647 – 664 .
- Weisshaupt , N. , Arlettaz , R. , Reichlin , T. S. , Tagmann-Ioset , A. and Schaub , M. 2011 . Habitat selection by foraging Wrynecks Jynx torquilla during the breeding season: identifying the optimal habitat profile . Bird Study , 58 : 111 – 119 .
- Whittingham , M. J. and Evans , K. L. 2004 . The effects of habitat structure on predation risk of birds in agricultural landscapes . Ibis , 146 ( Suppl. 2 ) : 10 – 220 .