Abstract
Capsule In winter diving ducks commonly move between wetlands more than 100 km apart, but short-distance daily movements dominate their activity.
Aims To describe the movements of Common Pochards Aythya ferina and Tufted Ducks Aythya fuligula during the wintering season in France and to investigate intrinsic and extrinsic factors potentially affecting the direction and distance of these movement.
Methods Distances, directions and patterns/strategies of winter movements were assessed from a combination of encounter data: physical recaptures at the ringing site, re-sightings of individuals fitted with nasal saddles, and ring recoveries of dead individuals.
Results Patterns of movements derived from multiple locations (i.e. successive re-sightings) of individuals could be assessed, showing that several ‘strategies’ may coexist in the same species (residency, exploitation of two wintering sites with or without return journey, and exploration of several sites). Individuals may use several sites more than 100 km apart during a given winter. Interestingly, directions of movements did not fit any previously observed pattern, especially in Common Pochards, whose movements tended to be biased towards the northeast at the beginning of the wintering season (October–November) whereas south or southwest movements were expected. Weather conditions in winter affected directions of movements in Common Pochards; eastwards movements towards ice-free Alpine lakes were more frequently observed during cold spells.
Conclusion Overall, our results suggest that exchanges of birds between wetlands more than 100 km apart are rather common even though short-distance movements related to daily activities are dominant. Our study provides new insights into the potential origin of ducks suspected of spreading infectious diseases.
INTRODUCTION
Studies on movements are essential to our understanding of the structure and dynamics of wildlife populations, and have important implications in terms of management and conservation. Movements are known to be crucial to avian population dynamics because of their obvious influence on mortality, emigration and immigration rates (Hestbeck et al. Citation1991, Iverson & Esler Citation2006). In migratory ducks, assessing movements is crucial for understanding spatial and/or temporal segregation of individuals according to gender and/or age (Carbone & Owen Citation1995), or for estimating the total number of birds using a wintering site (Hagmeier et al. Citation2008, Caizergues et al. Citation2011).
Winter movements of migratory ducks are known to be influenced by both intrinsic and extrinsic factors. Indeed, directions, distances and probability of winter movements have been found to differ between species, age and/or gender (Korner-Nievergelt et al. Citation2009, Brochet et al. Citation2010) as well as according to weather conditions (Ridgill & Fox Citation1990, Wernham et al. Citation2002, Keller et al. Citation2009, Sauter et al. Citation2010). However, despite significant effort having been devoted to the study of duck movements (Tamisier & Tamisier Citation1981, Cox & Afton Citation1997, Guillemain et al. Citation2002, Legagneux et al. Citation2009), our understanding of winter movements at large spatial scales and of the factors which determine these movements remains sketchy owing to the lack of adequate data. Ring recoveries of dead individuals, upon which most monitoring schemes have been designed, have not allowed the collection of a large data set over short periods due to recovery rates rarely exceeding 10 % (Keller et al. Citation2009, Brochet et al. Citation2010). Moreover, virtually no data on individual patterns of movements and differences in movement strategies are available.
Nasal saddles, which have proven to be very useful for monitoring duck movements in North America (Blohm Citation1978, Evrard Citation1999), have recently been employed with success in European duck monitoring schemes (Rodrigues et al. Citation2001). However, to date, except for the Eurasian Teal Anas crecca (Guillemain et al. Citation2010), none of these monitoring schemes have investigated large-scale movements of ducks during the wintering period.
Here, we describe winter movements of two diving duck species, Common Pochards Aythya ferina and Tufted Ducks Aythya fuligula. We combine data on the re-sightings of ducks individually marked with nasal saddles, physical recaptures at ringing sites and ring recoveries. By combining these three sources of data, we were able to document a quite large number of movements over a reasonable period of time. First, we compared the different methods, and more particularly re-sightings, of individuals equipped with nasal saddles and ring recoveries of hunted individuals to assess their respective contribution to the analyses of movements. More particularly, we compared the quantity of data collected using these two approaches as well as distances of movements from the ringing site. Second, using data of individuals caught on our main study area (which was the only site to provide a reasonable amount of data over all wintering seasons and over the different periods within each wintering season), we investigated the intrinsic (gender, age, species, period of the wintering season or month) and extrinsic (North Atlantic Oscillation (NAO) and local winter severity indexes) factors potentially influencing the distances and directions of movements (Keller et al. Citation2009, Brochet et al. Citation2010). Finally, using all sources of encounter data combined, we selected individuals with multiple encounters within a wintering season to characterize the different movement patterns/strategies and to assess the influence of intrinsic and extrinsic factors on them.
MATERIALS AND METHODS
Study areas and data collection
On average, about 60 000 Common Pochards and 30 000 Tufted Ducks are counted in France at the peak of the wintering season (Fouque et al. Citation2009), i.e. about 4.5 % and 1.6 %, respectively, of their populations (1 350 000 Common Pochards/1 900 000 Tufted Ducks) in northwestern Europe and in the central Europe–Black Sea–Mediterranean flyways (Wetlands International Citation2006).
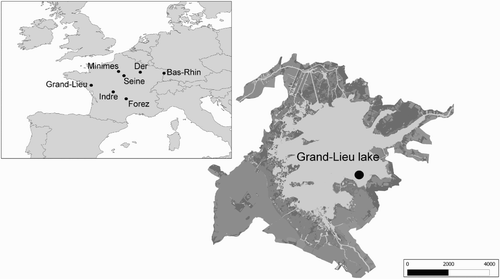
Common Pochards and Tufted Ducks were captured during the wintering season on the seven wintering areas of northern France: lake of Grand-Lieu, lake of Minimes, Seine, Indre, Forez, Bas-Rhin and lake of Der (). Our main study area, the lake of Grand-Lieu () is a shallow lake of about 6000 ha. Numbers of Common Pochards on Grand-Lieu can reach 5000 individuals at the peak of the wintering season, whereas numbers of breeding pairs average 700 (Reeber Citation2010). Breeding of Tufted ducks on the lake is anecdotal but winter peak numbers can reach up to 400 individuals.
Table 1. The number of Common Pochards and Tufted Ducks marked on the seven study areas (years of monitoring in the brackets) and the number of birds re-encountered alive (seen alive = physical recaptures and re-sightings), recovered dead, re-encountered alive > 10 km and < 10 km from the ringing site and recovered dead < 10 km from the ringing site.
Figure 1. The lake of Grand-Lieu and the seven other ringing sites of Common Pochards and Tufted Ducks which are part of our monitoring scheme in France. On Grand-Lieu, light grey areas represent open water and aquatic vegetation; dark grey areas indicate the various habitats types surrounding open water (prairies, crops, reed banks, forested areas, etc.). The black dot is the ringing site. (Scale: Grand-Lieu = m, French territory and Western Europe, km).
Ducks were captured from early October to February with barrier traps baited with wheat or corn (Haramis et al. Citation1982). Each individual was aged (yearlings versus adults) and sexed after plumage patterns and/or by checking the presence/absence and size of penis (Rousselot & Trolliet Citation1991). Each individual received a numbered metal ring and was fitted with a nasal saddle bearing an individual code readable using a telescope from up to 250 m (Rodrigues et al. Citation2001). Re-sightings of nasal saddled birds and ring recoveries were collected throughout the wintering season by volunteer birdwatchers or hunters. Except at the lake of Minimes, no special effort was made to read the codes on nasal saddles at the ringing sites.
Data used in our study combine re-sightings of nasal saddled ducks, physical recaptures and ring recoveries of 2696 individuals (1970 Common Pochards, 726 Tufted Ducks) captured in France during 7 wintering seasons (October–February), from 2003–04 to 2009–10 ().
Ring recovery versus re-sighting of nasal saddled individuals
To better assess the contribution of each type of data to the analyses of movements, numbers of locations per individual and distances from ringing sites were computed separately for ring recoveries and re-sightings. In these ‘global’ analyses, data from birds ringed in the seven studies areas were considered together.
To make comparisons between ring recoveries and re-sightings meaningful, when an individual was observed alive several times within a wintering season, one of the re-sightings was randomly selected to compute the movement distance from the ringing site.
Intrinsic and extrinsic factors influencing movement distances and directions
Possible effects of intrinsic and extrinsic factors on distances and directions of movements were assessed using data collected on Grand-Lieu. We did not include data from other sites to avoid possible biases due to unequal sampling (ringing) effort (). Moreover, Grand-Lieu was the only site where substantial numbers of individuals of both species could be ringed each winter, thereby allowing us to test the effects of species, month (or period of the winter) and intrinsic factors on movements.
Intrinsic factors tested in our analyses included sex, age (yearling versus adults), species, month and/or period of the winter (autumn migration: October–November versus midwinter: December–February). We opted for this delineation of periods to better take into account possible differences in movement patterns/propensities. Autumn migration is characterized by an increase in duck numbers on stopover and wintering areas and potentially includes large movement distances linked to post-nuptial migration. In contrast, midwinter is characterized by a relative stability in numbers and is therefore expected to be a period of limited movements (the peak of the pre-nuptial migration generally occurring later in the season) (Scott & Rose Citation1996). To take into account possible effects of time separating two encounters on distance estimates, a time effect (measured as the number of days separating encounter occasions from which the distance was computed) was included in the analyses.
Extrinsic factors included in our analyses were the Hellmann index of winter severity (WS) (Ridgill & Fox Citation1990) and the North Atlantic Oscillation (NAO) index (Hurrell Citation2012). WS was defined as the sum of the absolute value of all negative daily mean temperatures recorded from 1 December to 28 February at the ‘Météo France’ recording station of Rennes (100 km away from Grand-Lieu). The NAO index, which gives information about more global weather conditions, is known to correlate well with various ecological parameters in a variety of habitat types (Stenseth et al. Citation2003, Rainio et al. Citation2006). The variable used in our study was the principal component-based extended winter NAO index (December–March mean).
To avoid possible biases due to unequal encounter occasions between individuals, distances and directions of movements were computed between the location of the ringing site (Grand-Lieu) and a randomly selected location among all types of subsequent locations available (recoveries, physical recaptures and re-sightings) within a given interval of time (either autumn migration, midwinter or month). Movements of less than 10 km, which typically result from daily activities (Tamisier & Dehorter Citation1999, Guillemain et al. Citation2002, Herring & Collazo Citation2005, Legagneux et al. Citation2009, Davis & Afton Citation2010) were discarded from the analyses.
The distances (km) and azimuth (degrees) between two locations were derived from Cartesian coordinates (WGS1984). The estimated distance was orthodrome, i.e. distances along great circle routes (the closest route between two positions on the globe).
All distances and directions of movements retained in theses analyses were computed from a single individual, meaning that our data set was devoid of pseudo-replication.
Movement patterns/strategies and their determinism
Movement patterns/strategies were assessed using data of individuals with at least three encounters either during autumn migration or during midwinter or in both periods. To enhance our sample size, data from birds ringed in the seven studies areas were considered together.
Possible variations of movement patterns/strategies according to intrinsic (sex, age, species, period: autumn-migration versus midwinter) and extrinsic (winter severity and NAO, see below) factors were tested. To avoid pseudo-replication due to the same individual being followed during both autumn migration and midwinter or over several wintering seasons, statistical analyses were performed by randomly selecting one pattern of movement per individual.
Statistical analyses
All statistical analyses presented here were performed using R 2.10.1 software (R Development Core Team Citation2006). Descriptive statistics are expressed as means ± sd.
Method comparison
Distribution of movement distances between methods (recoveries and re-sighting data) were compared using permutation tests (Manly Citation1997).
Movement distances
The determinism of movement distances was assessed using ‘general linear models’ in three steps. In a first step, we tested the effects of intrinsic factors (species, age, sex, period) on distances computed over autumn migration and midwinter separately, taking into account the time elapsed since first encounter. In a second step, the same intrinsic effects were tested on monthly distances. In the third and last step, the effects of both intrinsic and extrinsic factors (winter severity, NAO index) were tested on distances measured during midwinter (the period in which variations in weather conditions were expected to be the most variable).
All theses analyses were run on log-transformed distances, whose normality was checked using a quantile-plot (Cleveland Citation1993).
Model selection was performed using the Akaike's information criterion corrected (AICc) for small sample-sizes (Burnham & Anderson Citation2002). Parameter estimates were derived either from the best model or by using model averaging of models within 2 AICc values.
Movement directions
Movement directions were assessed using circular statistics (Rayleigh test of uniformity). Effects of age, sex, species, period, WS and NAO index were assessed using a randomization procedure on the test of Wheeler–Watson (Jammalamadaka & SenGupta Citation2001). The analyses were run using the same three-step approach as described above, except that no adjustment for the time elapsed since ringing was needed. WS and NAO indexes, which are quantitative variables, had to be transformed into qualitative variables (factors). Two classes were retained for each of these variables depending on whether the values were above or below the average values recorded over the whole study period (winter 2005–05 to winter 2009–10).
Movement patterns/strategies
Possible differences of patterns/strategies according to intrinsic and extrinsic factors were tested using ‘multinomial log linear models’ (multinom procedure in R) in two steps. As with distances and directions of movements, we first assessed the effects of intrinsic factors (sex, age, species and period) on patterns/strategies of movements computed separately for autumn migration and midwinter, whereas intrinsic factors (WS and NAO index) were assessed for patterns observed during midwinter only. Due to limited sample sizes, only the main effects could be taken into account in this analysis. Model selection was done using the same AICc approach as for distances (see above).
RESULTS
Method comparison
Six hundred and nine individuals (472 Common Pochards and 137 Tufted Ducks) were physically recaptured at least once after marking and 926 individuals (573 Common Pochards and 353 Tufted Ducks) were re-sighted at least once after marking (for a total of 3992 re-sightings). Some individuals could be trapped up to 16 times, while others could be re-sighted up to 120 times (). Most physical recaptures (73.3 % Common Pochard, 67.9 % Tufted Duck) were made during the same season as ringing, whereas a substantial proportion of re-sightings (61.8 % for Common Pochard, 51.6 % for Tufted Duck) were made in subsequent wintering seasons.
Figure 2. Distribution of (a) recaptures, (b) resightings of Common Pochards and Tufted Ducks from 2003–04 to 2009–10.
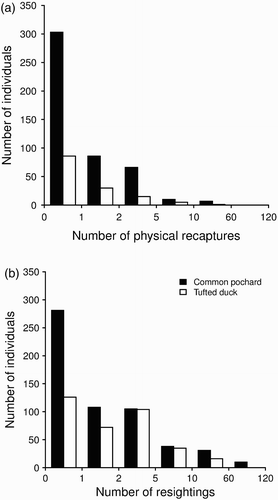
Overall, 53.5 % (n = 2 696) of birds fitted with a nasal saddle were encountered (re-sighted, recaptured and/or recovered) at least once after marking during the study period. For these birds, the average number of encounters (including marking) was 4.7 (± 8.5, range: 2–121) if we include data from different wintering seasons and 3 (± 4.7, range: 1–56) during a given wintering season.
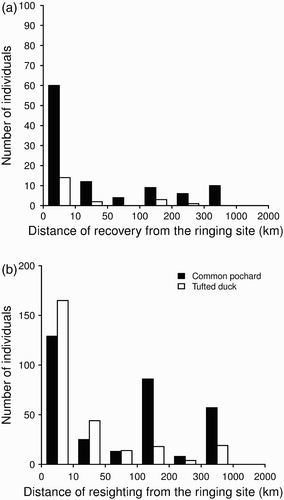
All physical recaptures were made at the site of ringing. Similarly, a large proportion (62.2 %) of individuals was recovered dead in the immediate vicinity of the ringing site. Recovery distances and re-sighting distances showed a similar distribution in Tufted Ducks (Permutation test: P = 0.5, re-sighting distance 52.8 ± 117 and recovery distance 39.8 km ± 74.2 km) but not in Common Pochards, for which movements > 100 km computed from re-sightings were more frequently observed than movements < 10 km, whereas the opposite was true for movements computed using dead recoveries (Permutation test: P < 0.01, re-sighting distance 131.6 ± 161.2 and recovery distance 76.8 km ± 134.4 km, ).
Figure 3. Distribution of movement distances between the ringing site and the site of recovery (a) or the site of resighting (b) (randomly drawn resighting locations among all available resighting locations), performed by Common Pochards and Tufted Ducks during the same wintering season of ringing. The time interval between ringing and recovery was 17.8 ± 13.7 days (1–59) for Tufted Duck and 31 ± 21.6 days (1–92) for Common Pochard. The time interval between ringing and resighting was 37.8 ± 36 days (0–154) for Tufted Ducks and 45.5 ± 36.9 days (0–149) for Common Pochards.
Movement distances
Among 2152 birds marked on the lake of Grand-Lieu (1764 Common Pochards and 388 Tufted Ducks), 396 (362 Common Pochards and 34 Tufted Ducks) and 317 (192 Common Pochards and 125 Tufted Ducks) individuals were encountered at least twice during autumn migration and midwinter, respectively.
In both species, the majority of these birds did not leave the study area (autumn migration: 87 % for Common Pochards and 71 % for Tufted Ducks; midwinter: 74 % for Common Pochards and 70 % for Tufted Ducks). All 86 movements recorded during midwinter remained within the limits of France, whereas 5 movements out of the 57 recorded during autumn migration led the individuals to leave the country ().
Figure 4. Movements of Common Pochards and Tufted Ducks from the lake of Grand-Lieu during autumn migration (October–November, n = 57) and midwinter (December–February, n = 86). Only movements greater than 10 km are shown.
Movement distances were neither affected by the time elapsed since first encounter nor by sex, age, species, period or month (). Similarly, we did not detect any significant effect of winter severity or NAO index fluctuations on movement distances during midwinter (). Estimates of seasonal distances of movements averaged over the best models were all within 7 km, with a ‘grand average’ of 158.9 km (± 147.9 sd, n = 143). In both species about 23 % of all recorded movements (>10 km) were of more than 200 km.
Table 2. Model selection for the analysis of the determinism of log-transformed distances of movements computed by period (autumn migration versus midwinter) or month. Global models for period effect and monthly distances included main effects (time, age, sex, species, month/period or NAO index and WS) and their second-order interactions. AICc, Aikaike information criterion for model selection corrected for small sample size; WS, winter severity; time, number of days between two encounters. Movement distances averaged over the best models were within 7 km.
Movement directions
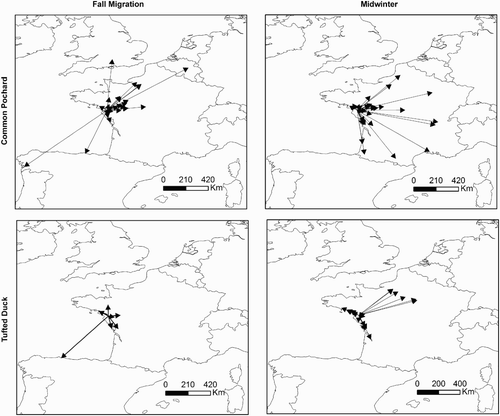
Species-specific differences in movement directions were detected during both autumn migration and midwinter (autumn migration, Wheeler–Watson test = 0.1, P = 0.002; midwinter, Wheeler–Watson test = 0.063, P = 0.005). While Common Pochards mainly moved towards the northeast whatever the period or month considered (Wheeler–Watson test = 0.028, P = 0.068 and Wheeler–Watson test = 0.12, P = 0.2, respectively; mean direction 290.6°, rho = 0.56, Rayleigh test, P < 0.001, n = 97), Tufted Ducks did not favour any particular direction (autumn migration, Rayleigh test, P = 0.2, n = 10; midwinter, Rayleigh test, P = 0.13, n = 36). About 38.9 % of Tufted Ducks moved towards the northwest during midwinter, whereas none of the Common Pochards did so.
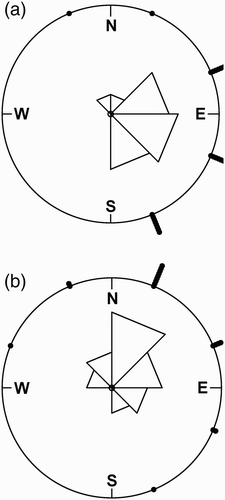
Winter severity and fluctuation in the NAO index tended to influence movement directions of Common Pochards during midwinter (Wheeler–Watson test = 0.1, P = 0.07 and Wheeler–Watson test = 0.16, P = 0.001, respectively), but not that of Tufted Ducks (Wheeler–Watson test = 0.002, P = 0.95 and Wheeler–Watson test = 0.01, P = 0.72, respectively). Common Pochards mainly moved in the northeast direction during mild and dry winters (mean direction 316.5°, rho = 0.7, Rayleigh test, P < 0.001, n = 19 and mean direction 315.2°, rho = 0.71, Rayleigh test, P < 0.001, n = 24 for winter severity and NAO index, respectively) whereas they preferentially moved towards the east during winters colder and wetter than the average (mean direction 257°, rho = 0.74, Rayleigh test, P < 0.001, n = 31 and mean direction 248.6°, rho = 0.8, Rayleigh test, P < 0.001, n = 26 for winter severity and NAO index, respectively) ().
Figure 5. Directions of movements of Common Pochards from the lake of Grand-Lieu during midwinter (December–February) according to winter severity: (a) winters colder than the average (Hellmann index > mean, i.e. 7.43) and (b) winters milder than the average (Hellmann index < 7.43).
Movement patterns/strategies
Most birds with multiple encounters covered movement distances of less than 10 km (autumn migration = 81 %, n = 192; midwinter = 70 %, n = 341). Three types of movements could be distinguished among the 138 individuals displaying at least one movement of more than 10 km (). (1) Most individuals (76 %) displayed ‘one-way’ movements between the site where they were initially seen or captured and another wintering site up to 649–794 km away; movements of less than 10 km were recorded either on the site of origin, in the newly occupied sites, or in both categories of sites. (2) of these, 13 % (138 individuals) displayed ‘round-trip’ movements between the site of origin and another wintering site up to 396–782 km away. In this case, too, short-distance movements could be recorded on either site. (3) Finally, 11 % of individuals displayed ‘exploratory’ movements, being seen each time in a different place more than 10 km away from the previous one and having performed movements in different directions.
Table 3. The different types of movements including at least one long distance (> 10 km) movement, recorded during autumn migration (October–November) and midwinter (December–February) for Common Pochards and Tufted Ducks, marked in the seven study sites. Only individuals being encountered at least three times including the first encounter (ringing or recapture) were included in these analyses.
Neither sex, age or species effect could be detected on the four types of patterns computed within each period of the wintering season (‘residency’ = movements < 10 km, ‘one-way’, ‘round-trip’ and ‘exploratory’). In contrast, whatever their type, movements were significantly more frequently recorded during midwinter than during autumn migration (). However, this result could be an artefact due the fact the midwinter was one month longer than autumn migration, making large-scale movements more likely detectable during this period. Neither extrinsic nor intrinsic factors effects could be detected on the different movement patterns observed during midwinter ().
Table 4. Model selection for the analyses: (1) of the effects of intrinsic factors (age, sex, species, period) on patterns of movements computed for autumn migration and midwinter separately and (2) of the effects of intrinsic and extrinsic factors (WS and NAO index) on patterns of movements during midwinter (December–February) only. Due to small sample size global models included all main effects only. AICc, Aikaike information criterion for model selection corrected for small sample size.
DISCUSSION
Movements of wintering diving ducks have previously been investigated in Europe based on ring recoveries (Keller et al. Citation2009, Brochet et al. Citation2010). These studies have greatly helped the understanding of duck movements during winter, showing that movements of more than 200 km were not uncommon both in diving and dabbling ducks and that their frequency tended to increase with winter severity. Using data from re-sightings of individuals fitted with nasal saddles, we confirm the view that long-distance movements are not uncommon in these two diving duck species and provide new insights into their patterns.
Overall, even though short-distance movements were registered more often than long-distance movements, all data types and distance estimates supported the hypothesis that long-distance movements are relatively common in both species. Moreover, short-distance movements might have been overestimated simply because the density of marked individuals was probably always much higher around our capture sites than elsewhere, making recoveries and/or re-sightings more likely in their vicinity. Due to small sample sizes, we could not take into account possible spatial variations in encounter or survival probabilities, although such variations could have affected both our estimates of movement distances and statistical tests. For example, although the wintering ranges of both species are known to extend to North Africa (Gilissen et al. Citation2002), neither recoveries nor re-sightings were reported in this area, probably owing to low human densities. Overall, however, due to the fact that two different types of data (re-sighting of alive versus dead recoveries of hunted individuals) were analysed, we can assume that possible biases were probably reduced compared to previous studies. With the exception of the lake of Minimes in Vincennes near Paris, neither local recoveries nor local re-sightings displayed strong differences () between study sites, suggesting that at this spatial scale (at least) no dramatic differences in sampling effort could be expected.
Directions of movements from the lake of Grand-Lieu did not fit any previously observed pattern, especially concerning Common Pochards. Indeed, whatever the gender or age, Common Pochards were more likely to move towards the northeast. This direction would be unusual during the autumn migration for individuals belonging to the North-Western European flyway, which are supposed to migrate towards the southwest during this period of the year (Scott & Rose Citation1996). Migration towards the northeast has recently been documented for both Common Pochards and Tufted Ducks breeding in western France (Guillemain & Caizergues Citation2010). The present pattern observed during autumn migration may be partly due to these local breeders undertaking migratory movements late in the season.
In our study, Tufted Ducks were found to move in large numbers towards the northwestern French coast during winter, an area characterized by the presence of large numbers of coastal lagoons where they may feed on molluscs. This type of movement was expected for Tufted Ducks, which are known to winter in estuaries and sheltered coastal areas (Ridgill & Fox Citation1990). Common Pochards, which tend to feed on aquatic weeds, bulbs and seeds, almost never moved in this direction during the same period. It seems clear that such differences in movement patterns between the two species may be explained by differences in food preferences (Olney Citation1963, Citation1968).
Whereas no firm evidence of an influence of weather conditions on movement distances was found in either species, both winter severity and fluctuations in NAO index in mid-winter influenced the direction of movements, especially in Common Pochards. The lake of Grand-Lieu, as well as most other inland water bodies, froze during several days of the two harsher than average winters (2008/2009 and 2009/2010). Surprisingly, instead of moving towards the south, birds favoured movements towards the east during these periods of frost, despite most inland water bodies being supposed to have frozen. Large lakes of the pre-Alpine area, however, rarely freeze, even during the most intense cold spells. This is probably why they are among the most important wintering areas of the eastern European region for diving ducks (Ridgill & Fox Citation1990). Therefore, one can assume that by moving towards the east during cold spells, birds were trying to reach ice-free lakes of the pre-Alpine area. To confirm this hypothesis, it would be very interesting to assess a possible negative correlation between numbers of Common Pochards in Western France and on Alpine lakes.
Patterns of movements derived from multiple locations of single individuals suggest that several winter strategies may coexist in the same species. Most individuals stayed in the same area throughout winter, performing only local movements probably between resting and foraging sites. At the other end of the spectrum, some individuals seemed to definitively leave the wintering area where they were initially ringed to settle down on another site up to 800 km away or to explore several wintering sites without apparently settling down. In between these two extreme ‘strategies’ were individuals performing round-trips between two wintering areas separated by up to 780 km. To our knowledge, our study is the first to document movements of wintering diving ducks based on multiple encounters for a given individual. However, more data are needed to properly assess individual differences concerning these patterns of movements. Such analyses would greatly help to understand the factors underlying duck movement strategies.
In conclusion, our study suggests that an exchange of birds may exist between distant wintering sites more than 100 km apart. Furthermore, individuals caught in western France during autumn migration have been found to move in the direction of the northeast, wintering in northwestern France, the Netherlands, Belgium or Great Britain rather than in southern Europe or North Africa. To our knowledge, such a pattern of movement has never before been described. In terms of epidemiology, our study provides new insights into the potential origin of ducks suspected of spreading infectious diseases, in particular concerning Common Pochards which were suspected of being responsible for the introduction of highly pathogenic avian influenza viruses in the east of France in 2006 (Le Gall-Reculé et al. Citation2008).
ACKNOWLEDGEMENTS
This study was supported by the Office National de la Chasse et de la Faune Sauvage. Many thanks are due to Clément Calenge, Matthieu Guillemain and Jean-Sébastien Guitton for their helpful suggestions on previous versions of the manuscript. The authors would also like to warmly thank Catherine Carter for improving the English. We are especially grateful to the ringers: Christophe Sorin (FDC 44), Nicolas Flamant (VétoAgroSup), Jean-François Arcanger and Eric Marouzé (FDC 53), François Bourguemestre (FDC 36), Olivier Landelle et Vincent Fontaine (ONCFS SD53), Thierry George (ONCFS SD51), Christine Blaize and David Hémery (Grumpy Nature), André Momerency (ONCFS), Gilles Delacour (ONCFS), Jean-Yves Fournier (ONCFS) and Benoît Castanier and Jean-Philippe Rabatel (Fondation Pierre Vérots). Many thanks to birdwatchers and hunters for having provided observations or ring recoveries. Finally we are indebted to people at the French ringing centre (CRBPO) for providing the rings and for their invaluable help in the management of the database.
REFERENCES
- Blohm , R. J. 1978 . Migrational homing of Male Gadwalls to breeding grounds . Auk , 95 : 763 – 766 .
- Brochet , A.-L. , Guillemain , M. , Lebarbenchon , C. , Simon , G. , Fritz , H. , Green , A. J. , Renaud , F. , Thomas , F. and Gauthier-Clerc , M. 2010 . The potential distance of highly pathogenic avian influenza virus dispersal by Mallard, Common Teal and Eurasian Pochard . EcoHealth , 6 : 449 – 457 .
- Burnham , K. P. and Anderson , D. R. 2002 . Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach , New York : Springer-Verlag .
- Caizergues , A. , Guillemain , M. , Arzel , C. , Devineau , O. , Leray , G. , Pilvin , D. , Lepley , M. , Massez , G. and Schricke , V. 2011 . Emigration rates and population turnover of Teal (Anas crecca) in two major wetlands of Western Europe . Wildlife Biol. , 17 : 373 – 382 .
- Carbone , C. and Owen , M. 1995 . Differential migration of the sexes of Pochard Aythya ferina: results from a European survey . Wildfowl , 46 : 99 – 108 .
- Cleveland , W. 1993 . Visualizing Data , Summit, NJ : Hobart Press .
- Cox , R. R. and Afton , A. D. 1997 . Use of habitats by female northern pintails wintering in southwestern Louisiana . J. Wildl. Manage. , 61 : 435 – 443 .
- Davis , B. E. and Afton , A. 2010 . Movement distances and habitat switching by female mallards wintering in the lower Missipi alluvial valley . Waterbirds , 33 : 349 – 356 .
- Evrard , J. O. 1999 . Mallard and Blue-winged Teal philopatry in northwest Wisconsin . North American Bird Bander , 24 : 38 – 42 .
- Fouque , C. , Guillemain , M. and Schricke , V. 2009 . Trends in the numbers of Coot Fulica atra and wildfowl Anatidae wintering in France, and their relationship with hunting activity at wetland sites . Wildfowl special issue , 2 : 42 – 59 .
- Gilissen , N. , Haanstra , L. , Delany , S. , Boere , G. and Hagemeijer , W. 2002 . Numbers and distribution of wintering waterbirds in the Western Palearctic and Souwest Asia in 1997, 1998 and 1999. Results form the International Waterbirds Census . Wetland International Global Series , No. 11
- Guillemain , M. and Caizergues , A. 2010 . L'étude des canards par le biais des marques nasales: intérêt de la méthode et résultats . Ornithos , 17 : 290 – 298 .
- Guillemain , M. , Devineau , O. , Brochet , A.-L. , Fuster , J. , Fritz , H. , Green , A. J. and Gauthier-Clerc , M. 2010 . What is the spatial unit for a wintering Teal Anas crecca? Weekly day roost fidelity inferred from nasal saddles in the Camargue, southern France . Wildlife Biol. , 16 : 215 – 220 .
- Guillemain , M. , Fritz , H. and Duncan , P. 2002 . The importance of protected areas as nocturnal feeding grounds for dabbling ducks wintering in western France . Biol. Conserv. , 103 : 183
- Hagmeier , K. R. , Smith , B. D. and Boyd , W. S. 2008 . Estimating numbers of black brant using sequential spring-staging sites . J. Wildl. Manage. , 72 : 1342 – 1351 .
- Haramis , G. M. , Derleth , E. L. and McAuley , D. G. 1982 . Techniques for trapping, aging, and banding wintering Canvasbacks . J. Field Ornithol. , 53 : 342 – 351 .
- Herring , G. and Collazo , J. A. 2005 . Habitat use, movements and home range of wintering lesser scaup in Florida . Waterbirds , 28 : 71 – 78 .
- Hestbeck , J. B. , Nichols , J. D. and Malecki , R. A. 1991 . Estimates of movement and site fidelity using mark–re-sight data of wintering Canada geese . Ecology , 72 : 523 – 533 .
- Hurrell, J.W. 2012. North Atlantic Oscillation (NAO) indices information. Available at: http://www.cgd.ucar.edu/cas/jhurrell/indices.html.
- Iverson , S. A. and Esler , D. 2006 . Site fidelity and the demographic implications of winter movements by a migratory bird, the Harlequin Duck Histrionicus histrionicus . J. Avian Biol. , 37 : 219 – 228 .
- Jammalamadaka , R. S. and SenGupta , A. 2001 . Topics in Circular Statistics , London : World Scientific Publishing .
- Keller , I. , Korner-Nievergelt , F. and Jenni , L. 2009 . Within-winter movements: a common phenomenon in the Common Pochard Aythya ferina . J. Ornithol. , 150 : 483 – 494 .
- Korner-Nievergelt , F. , Hofer , J. , Sauter , A. and Jenni , L. 2009 . Measuring within-winter movements rates of Tufted Duck Aythya fuligula and Common Pochard A. ferina based on ring re-encounter data . Wildfowl special issue , 2 : 24 – 41 .
- Le Gall-Reculé , G. , Briand , F.-X. , Schmitz , A. , Guionie , O. , Massin , P. and Jestin , V. 2008 . Double introduction of highly pathogenic H5N1 avian influenza virus into France in early 2006 . Avian Pathol. , 37 : 15 – 23 .
- Legagneux , P. , Blaize , C. , Latraube , F. , Gautier , J. and Bretagnolle , V. 2009 . Variation in home-range size and movements of wintering dabbling ducks . J. Ornithol. , 150 : 183
- Manly , B. F.J. 1997 . Randomization, Bootstrap and Monte Carlo Methods in Biology , London : Chapman & Hall .
- Olney , P. J.S. 1963 . The food and feeding habits of Tufted Duck Aythya fuligula . Ibis , 105 : 55
- Olney , P. J.S. 1968 . The food and feeding-habits of the Pochard, Aythya ferina . Biol. Conserv. , 1 : 71
- R Development Core Team . 2006 . R: A Language and Environment for Statistical Computing , Austria : Vienna . (ed. R foundation for statistical Computing)
- Rainio , K. , Laaksonen , T. , Ahola , M. , Vahatalo , A. V. and Lehikoinen , E. 2006 . Climatic responses in spring migration of boreal and arctic birds in relation to wintering area and taxonomy . J. Avian Biol. , 37 : 507 – 515 .
- Reeber, S. 2010. Suivi ornithologique du lac de Grand-Lieu en 2009. Société Nationale de Protection de la Nature.
- Ridgill , S. C. and Fox , A. D. 1990 . Cold Weather Movements of Waterfowl in Western Europe , International Waterfowl and Wetlands Research Bureau special issue 13), pp. 1–89. International Waterfowl and Wetlands Research Bureau, Slimbridge
- Rodrigues , D. J.C. , Fabio , A. M.D. and Figueiredo , M. E.M.A. 2001 . “ The use of nasal markers for monitoring mallard populations ” . In Wildlife, Land, and People: Priorities for the 21st Century. Proceedings of the Second International Wildlife Management Congress , Edited by: Field , R. , Waren , R. J. , Okarma , H. and Sievert , P. R. 316 – 318 . Bethesda, Maryland , , USA : The Wildife Society .
- Rousselot , J. C. and Trolliet , B. 1991 . Critères de détermination du sexe et de l'âge des canards , Echirolles : Imprimerie Poncet .
- Sauter , A. , Korner-Nievergelt , F. and Jenni , L. 2010 . Evidence of climate change effects on within-winter movements of European Mallards Anas platyrhynchos . Ibis , 152 : 600 – 609 .
- Scott , D. A. and Rose , P. M. 1996 . Atlas of Anatidae Populations in Africa and Western Eurasia , The Netherlands : Wetlands International, Wageningen .
- Stenseth , N. C. , Ottersen , G. , Hurrell , J. W. , Mysterud , A. , Lima , M. , Chan , K. S. , Yoccoz , N. G. and Adlandsvik , B. 2003 . Studying climate effects on ecology through the use of climate indices: the North Atlantic Oscillation, El Nino Southern Oscillation and beyond . Proc. R. Soc. Lond. B , 270 : 2087 – 2096 .
- Tamisier , A. and Dehorter , O. 1999 . Camargue, canards et foulques: fonctionnement et devenir d'un prestigieux quartier d'hiver , Nîmes : Centre Ornithologique du Gard .
- Tamisier , A. and Tamisier , M. C. 1981 . Validation by biotelemetry of the functional unit concept among Camargue Teals . Terre Et La Vie-Revue D Ecologie Appliquee , 35 : 563 – 579 .
- Wernham , C. , Toms , M. , Marchant , J. , Clark , J. , Siriwardena , G. and Baillie , S. 2002 . The Migration Atlas: Movements of the Birds of Britain and Irleand , London : Poyser .
- Wetlands International . 2006 . Waterbird Population Estimates: Fourth Edition , Wageningen , , The Netherlands : Wetlands International .