Abstract
Capsule Two juvenile Harpy Eagles were tagged at their nests with satellite transmitters. During the first two years after hatching both birds moved very short distances, with a maximum recorded distance of 1.3 km from the nest. During the study period, neither bird started juvenile dispersal, but while one of the birds showed a significant overall increase in mean monthly distance from nest with age, the other bird did not. Protecting territories of adult Harpy Eagles may enhance the protection of juveniles for a long period of time.
In large, long-lived raptors with delayed maturity, the period between fledging and recruitment to the breeding population may take several years and is usually poorly understood (Whitfield et al. Citation2009). Juvenile dispersal represents the movements undertaken by juveniles, once they become independent from their parents, to find a breeding site (Clobert et al. Citation2001). The onset of juvenile dispersal is not easy to detect and usually it has been more or less arbitrarily defined. Nevertheless, a simple and adequate approach to estimate the onset of dispersal considers that this stage of bird's life-cycle starts when juveniles abandon their parental territory (sometimes based on when juveniles reach the mid-distance between adjacent nests in the region and remain beyond that distance for a given period of time; e.g. Ferrer Citation1993, Soutullo et al. Citation2006a, Cadahía et al. Citation2008). The period between the first flights and the onset of juvenile dispersal is known as the post-fledging dependency period, when juveniles are still dependent on their parents (e.g. Soutullo et al. Citation2006a). The fate of juvenile birds during these periods may have important consequences for population trends and, thus, they are of particular importance from a conservation point of view (Ferrer Citation2001, Whitfield et al. Citation2004, Penteriani et al. Citation2006, Citation2011, Soutullo et al. Citation2008a). However, because of the difficulty of obtaining adequate data during these periods, little is known on the juvenile stage of most raptor species (Penteriani & Delgado 2009).
The Harpy Eagle Harpia harpyja is one of the largest raptors of the American continent, being distributed from southern Mexico to northern Argentina (Ferguson-Lees & Christie Citation2001). Currently, the species is threatened across its whole distribution range and population numbers are declining, mainly due to human persecution and habitat loss and fragmentation (Vargas et al. Citation2006, BirdLife International Citation2012). For this reason, the Harpy Eagle is globally listed as Near Threatened (BirdLife International Citation2012). In Ecuador (where this study was conducted) it is listed as Vulnerable (Granizo et al. Citation2002). Although several aspects of its biology and ecology have been studied at several areas across its distribution range, such as Brazil (e.g. Galetti & de Carvalho Jr Citation2000, Lenz & Marajo dos Reis Citation2011), Ecuador (Muñiz-López 2007, 2008), Panama (e.g. Vargas & Vargas Citation2011), Peru (Piana Citation2007) and Venezuela (Álvarez-Cordero Citation1996), the Harpy Eagle is one of the least-known raptor species of the world. Most of the information available on the species is related to diet and adult behaviour at their breeding areas (Rettig Citation1978, Álvarez-Cordero Citation1996, Muñiz-López Citation2008) and recently also on population genetics (Banhos et al. Citation2008, Lerner et al. Citation2009). The species feeds primarily on tree-dwelling mammals, particularly monkeys and sloths, as well as on large birds and reptiles (Fowler & Cope Citation1964, Rettig Citation1978, Álvarez-Cordero Citation1996, Galetti & de Carvalho Jr Citation2000, Piana Citation2007, Muñiz-López Citation2008, Springer et al. Citation2011).
The Harpy Eagle has probably the longest breeding period of any raptor, usually breeding once every two and a half or three years (Rettig Citation1978, Álvarez-Cordero Citation1996). According to the few studies conducted on this species, adult home-ranges are generally large, ranging between 16 and 79 km2 (Álvarez-Cordero Citation1996, Muñiz-López Citation2007, Vargas & Vargas Citation2011). Juvenile Harpy Eagles have rarely been monitored and the only data regarding their first movements suggest that fledging occurs when birds are between 120 and 160 days old (Rettig Citation1978, Álvarez-Cordero Citation1996, Muñiz-López Citation2007). After that, juveniles are mostly inactive, making only short flights within the nest-tree or nearby trees, and being fully dependent on parents in terms of feeding (Álvarez-Cordero Citation1996). Harpy Eagles are supposed to be in this post-fledging dependency period for a long time (Álvarez-Cordero Citation1996), although no quantitative data to confirm this have been obtained to date.
In recent years, the use of satellite telemetry has become widespread and predominant in the study of migratory movements of several raptor species (e.g. Limiñana et al. Citation2007, Citation2012a,Citationb, Strandberg et al. Citation2010, López-López et al. Citation2010, Mellone et al. Citation2011), as well as in detailed studies of juvenile dispersal and habitat use (e.g. Urios et al. Citation2007, Soutullo et al. Citation2008b, Cadahía et al. Citation2010). Here, we use GPS-satellite telemetry to describe the movements of two free-ranging juvenile female Harpy Eagles in Ecuador during the first two years after hatching.
This study was conducted within the reserve ‘Reserva de Producción Faunística Cuyabeno’ (–0.117°N, –75.833°E, northeastern Ecuador). This reserve is dominated by rainforest, although habitats range from evergreen forests to flooded lowlands (Cerón et al. Citation1999, Palacios et al. Citation1999). There, two juvenile Harpy Eagles (‘Masakay’ and ‘Tava’) were trapped at their nests. ‘Masakay’ was tagged on 15 July 2006 in its third month after hatching, and ‘Tava’ on 28 March 2009 in its seventh month after hatching. Age of birds at tagging was known because the nests were monitored before the eggs hatched by Cofanes natives, who collaborated in the project and recorded hatching date. Both birds were sexed as females by means of body size and weight measures, according to reference values for juveniles in del Hoyo et al. Citation(1994). Eagles were tagged with 70-g Argos/GPS satellite transmitters (Microwave Telemetry Inc.), which were affixed to their backs using a Teflon harness. The full transmitter equipment did not exceed 1.5% of the juveniles' body mass, which is considerably below the 3% suggested to minimize the effects of additional mass on birds' movements (Kenward Citation2001). Satellite transmitters were programmed to record one GPS position (nominal accuracy ± 18 m) every hour between 09:00 and 03:00 for ‘Masakay’ and between 11:00 and 03:00 for ‘Tava’ (local time). Data were recorded until both individuals reached the age of two years (i.e. the last day of their 23rd month after hatching, taking into account age of birds at tagging). For both birds, we excluded data from the tagging month from the analyses, because it was only partially recorded.
During the study period, a total of 7170 GPS locations were obtained for ‘Masakay’ and 6337 for ‘Tava’. Locations were transformed from Geographic Coordinate System to UTM coordinates for calculations. For each individual, we calculated the loxodromic distance from every recorded position to its nest. We evaluated the relationship between the distance to nest and age of birds (months after hatching) using linear and quadratic regression analysis (distance to the nest ∼ bird age × bird identity + bird age squared × bird identity). Since these initial models presented heterogeneity of variances, we then conducted Generalized Least Square (GLS) regression models to overcome this analytical issue (Pinheiro & Bates Citation2000, Zuur et al. Citation2010). Therefore, considering the possible existence of proportional or potential structures of variances and also considering that different variance structures could emerge for each bird, these alternative variance structures were evaluated for each one of the independent variables (Zuur et al. Citation2009). In order to find the optimal variance structure we computed the same model using different variance structures and compared each GLS model using Akaike's Information Criterion (AIC) (Burnham & Anderson Citation2002, Zuur et al. Citation2009). All these models were fitted with the gls function from the nlme package (Pinheiro et al. Citation2011) for R software (R Development Core Team Citation2011). When doing this, the best model (lowest AIC value) was the one that included the second-order polynomic variable (distance to nest ∼ bird age × bird identity + bird age squared × bird identity) and included a power variance structure (varPower ∼ bird age|bird identity, in R), thus taking into account differences in monthly spread of data for each bird. As interactions in this model were significant (P < 0.001 in all cases), these results show that during the study period the two tracked juvenile eagles exhibited significantly different behaviours.
To deconstruct the movements pattern of each individual, we conducted GLS models separately to test if distance to nest showed a linear or quadratic trend for each individual (distance to nest ∼ bird age + bird age squared), also accounting for the observed heterogeneity of variances. The best individual GLS models (according to AIC) included the age squared term, but a different variance structure. The selected model for ‘Masakay’ included a power variance structure of the quadratic covariate (varPower ∼ bird age + squared bird age, in R) (parameter estimates: intercept = 16.1, P < 0.001; age = 2.7, P < 0.001; age squared = –0.1, P < 0.001). The selected model for ‘Tava’ included a variance structure proportional to the quadratic covariate (varFixed ∼ bird age + squared bird age, in R) (parameter estimates: intercept = 110.0, P < 0.001; age = –10.6, P < 0.001; age squared = 0.8, P < 0.001). These differences in parameter estimates revealed different individual trends in distance to nest during the study period (). Whereas ‘Tava’ showed an overall increasing trend in its monthly distances to nest, ‘Masakay’ first showed a slight increase in distances from the nest but then it began to reduce the extent of its movements from the nest, spending more time closer to the nest (). During the study period, maximum distance to the nest achieved by ‘Tava’ was 1300 m when it was 23 months old, whereas for ‘Masakay’ this maximum distance was 296 m at the age of 13 months ().
Figure 1. Movements of two Harpy Eagles (A: ‘Masakay’ and B: ‘Tava’), tracked by satellite telemetry during two years after hatching. Open dots represent every recorded distance to nest for every month within this two-year period. The line represents the quadratic model that better described the movements of both eagles in the study period (see text for details and parameter estimates).
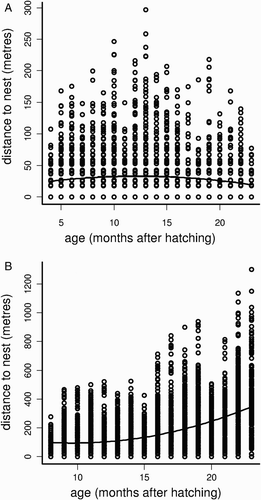
Juvenile dispersal is the ecological process that leads juvenile birds to find a site for breeding (Clobert et al. Citation2001). In large bird species with delayed maturity and relatively large home-ranges and breeding territories, this process may take several years until a breeding territory is occupied (e.g. Urios et al. Citation2007, Cadahía et al. Citation2009, Whitfield et al. Citation2009). Given the extent of the adult home-ranges in the Harpy Eagle, ranging between 14 and 79 km2 (Álvarez-Cordero Citation1996, Vargas & Vargas Citation2011), and considering the movements reported here, it is clear that none of the tracked juveniles left the parental territory during the study period. Therefore, the onset of the juvenile dispersal in the species does not take place within the first two years after hatching, and the stage of life reported here corresponds to the post-fledging dependence period (Soutullo et al. Citation2006a, Cadahía et al. Citation2008). Similar results for the Harpy Eagle were found by Álvarez-Cordero (1996), who reported distances to the nest of 300–600 m for juveniles 2 years old; however, the data of Álvarez-Cordero (1996) are based on direct observations, which, in the closed-canopy forests where Harpy Eagles live, may have precluded getting observations at larger distances from nest.
The individual eagles showed a different behaviour during the study period. ‘Tava’ showed a progressive distancing from nest during the study period, while ‘Masakay’ first showed a distancing from nest but then began to spend more time near the nest (see ). Progressive distancing from the nest during the post-fledging dependency period is common in several large raptors (e.g. O'Toole et al. Citation1999, Soutullo et al. Citation2006a,Citationb, Cadahía et al. Citation2008). However, during this dependency period, juvenile birds are mostly fed by parents, which may explain the constant return of both birds to their nests observed here. However, during this period, juvenile raptors also make their first hunting attempts within parental territories (e.g. Ferrer Citation1993, Kitowski Citation2009), and this has been also observed for Harpy Eagles (Muñiz-López Citation2007). Therefore, differences in prey abundance or availability in the areas used by these Harpy Eagles may explain, at least partially, the different behaviour of these individuals, with the eagle moving closer to its nest possibly being in a less-productive area.
Large, tropical birds usually show reduced clutch sizes and long breeding seasons due to the relative environmental stability of their habitats (Russell & Rowley Citation2000). As a consequence of the reduced brood size, parental care is usually longer and juveniles spend more time at the parental territory to enhance their survival probability (Skutch Citation1976) or to be in better condition at the onset of dispersal (Young Citation1996). Age of first reproduction for Harpy Eagles breeding in captivity has been estimated as 4 years; however, taking into account that the onset of juvenile dispersal for free-ranging individuals in undisturbed environments takes place at ages older than 2 years, age of first reproduction in the wild is likely to take place at ages probably much older than 4 years (see also Shaner Citation2011), which would represent one of the longest dispersal periods among raptors. However, as Harpy Eagles live in habitats that are largely homogeneous and undisturbed, there may be little need to cover very large distances to find a breeding site, contrary to other species in temperate heterogeneous habitats (e.g. Urios et al. Citation2007, Cadahía et al. Citation2009, Whitfield et al. Citation2009), which would reduce the overall time spent dispersing. Future tracking studies of individuals should focus on these aspects, because this floating period of large raptors is of great importance from a conservation point of view (Penteriani et al. Citation2006, Citation2011, Soutullo et al. Citation2008a).
Our results and conclusions are limited because of the small sample size that is often associated with such satellite-tracking studies. However, this approach has the advantage of obtaining spatially explicit information about raptors' movements, which is of great importance from a conservation point of view, even if only a few birds are tracked (e.g. Urios et al. Citation2010). In the case of the Harpy Eagle, this is exacerbated by the secretive nature of the species, the difficulties in finding their nests, as well as carrying out the fieldwork in remote dense forests. Results presented here indicate that territories occupied by adults are also important for juvenile birds for at least 2 years after hatching. Therefore, conservation actions at these areas should not only be focused during the pre-fledging period, but also throughout the year, given that the juveniles are actively using the same areas used by parents for a long time. Juvenile survival is one of the most important parameters determining population trends in long-lived raptors (Soutullo et al. Citation2008a). Hence, protecting these adult territories would also not only directly affect adult survival but also juvenile survival, which would be of paramount importance for conservation of the Harpy Eagle.
ACKNOWLEDGEMENTS
We are indebted to Fundación Terra Natura for funding the Project on satellite tracking of Harpy Eagles in Ecuador. We would like to thank Alexander Blanco for his support and help in capturing wild Harpy Eagles. We are also thankful to the indigenous communities of the study area for their essential work with the species. The Environment Ministry of Ecuador and EcoFondo funded the fieldwork. A. Soutullo and P. López-López provided insightful comments on an earlier draft of the manuscript. We are also grateful to P. Whitfield and W. Cresswell for their comments, which substantially improved the manuscript. R. Limiñana had a postdoctoral grant (reference 10/12-C) co-funded by ‘Consejería de Educación y Ciencia’ (JCCM) and the European Social Fund. Gonzalo D. Cortés is supported by a ‘Beca de Maestría’ of the Sistema Nacional de Becas of the Agencia Nacional de Investigación e Innovación (reference POSNAC 2011 POS-2011-1-3383). This article is part of the PhD thesis of R. Muñiz-López at the University of Alicante.
REFERENCES
- Álvarez-Cordero, E. 1996. Biology and conservation of the Harpy Eagle in Venezuela and Panama. PhD Thesis, University of Florida.
- Banhos , A. , Hrbek , T. , Gravena , W. , Sanaiotti , T. and Farias , P. 2008 . Genomic resources for the conservation and management of the harpy eagle (Harpia harpyja Falconiformes, Accipitridae) . Genet. Mol. Biol. , 31 : 146 – 154 . (doi:10.1590/S1415-47572008000100025)
- BirdLife International . 2012 . Species factsheet: Harpia harpyja . Downloaded from http://www.birdlife.org (accessed 15 March 2012)
- Burnham , K. P. and Anderson , D. R. 2002 . Model Selection and Multimodel Inference. A Practical Information–Theoretic Approach, 2nd ed , New York : Springer .
- Cadahía , L. , López-López , P. , Urios , V. and Negro , J. J. 2008 . Estimating the onset of dispersal in endangered Bonelli's Eagle Hieraaetus fasciatus tracked by satellite telemetry: a comparison among methods . Ibis , 150 : 416 – 420 . (doi:10.1111/j.1474-919X.2007.00781.x)
- Cadahía , L. , López-López , P. , Urios , V. and Negro , J. J. 2009 . Natal dispersal and recruitment of two Bonelli's eagles Aquila fasciata: a four year satellite tracking study . Acta Ornithol. , 44 : 193 – 198 . (doi:10.3161/000164509X482777)
- Cadahía , L. , López-López , P. , Urios , V. and Negro , J. J. 2010 . Satellite telemetry reveals individual variation in juvenile Bonelli's eagle dispersal areas . Eur. J. Wildl. Res. , 56 : 923 – 930 . (doi:10.1007/s10344-010-0391-z)
- Cerón , C. , Palacios , W. , Valencia , R. and Sierra , R. 1999 . “ Las formaciones naturales de la Costa del Ecuador ” . In Propuesta preliminar de un sistema de clasificación de Vegetación para el Ecuador Continental , Edited by: Sierra , R. Quito : Ministerio del Medio Ambiente, Proyecto INEFAN/GEF-BIRF y EcoCiencia .
- Clobert , J. , Danchin , E. , Dhont , A. A. and Nichols , J . 2001 . Dispersal, Causes, Consequences and Mechanisms of Dispersal at the Individual, Population and Community Level , Edited by: Clobert , J. , Danchin , E. , Dhont , A. A. and Nichols , J . Oxford : Oxford University Press .
- del Hoyo , J. , Elliot , A. and Sargatal , J. 1994 . Handbook of the Birds of the World , Vol. 2 , Barcelona : Lynx Editions .
- Ferguson-Lees , J. and Christie , D. A. 2001 . Raptors of the World , London : Christopher Helm .
- Ferrer , M. 1993 . Juvenile dispersal behavior and natal philopatry of a long lived raptor, the Spanish Imperial Eagle Aquila adalberti . Ibis , 135 : 132 – 138 . (doi:10.1111/j.1474-919X.1993.tb02824.x)
- Ferrer , M. 2001 . The Spanish Imperial Eagle , Barcelona : Lynx Editions .
- Fowler , J. and Cope , J. 1964 . Notes on the Harpy Eagle in British Guiana . Auk , 81 : 257 – 273 . (doi:10.2307/4082683)
- Galetti , M. and de Carvalho Jr . 2000 . Sloths in the diet of a Harpy Eagle nestling in Eastern Amazon . Wilson Bull. , 112 : 535 – 536 . (doi:10.1676/0043-5643(2000)112[0535:SITDOA]2.0.CO;2)
- Granizo , T. , Pacheco , C. , Ribadeneira , M. B. , Guerrero , M. and Suárez , L. 2002 . Libro rojo de las aves de Ecuador , Quito : SIMBIOE-Conservación Internacional-Ecociencia-Ministerio del Ambiente-UICN .
- Kenward , R. E. 2001 . A Manual for Wildlife Radio Tagging , London : Academic Press .
- Kitowski , I. 2009 . Social learning of hunting skills in juvenile marsh harriers Circus aeruginosus . J. Ethol. , 27 : 327 – 332 . (doi:10.1007/s10164-008-0123-y)
- Lenz , B. B. and Marajó Dos Reis , A. 2011 . Harpy Eagle–primate interactions in the Central Amazon . Wilson J. Ornithol. , 123 : 404 – 408 . (doi:10.1676/10-171.1)
- Lerner , H. R.L , Johnson , J. A. , Lindsay , A. R. , Kiff , L. F. and Mindell , D. P. 2009 . It's not too late for the Harpy Eagle (Harpia harpyja): high levels of genetic diversity and differentiation can fuel conservation programs . PLoS ONE , 4 ( 10 ) : e7336 (doi:10.1371/journal.pone.0007336)
- Limiñana , R. , Soutullo , A. and Urios , V. 2007 . Autumn migration of Montagu's harriers Circus pygargus tracked by satellite telemetry . J. Ornithol. , 148 : 517 – 523 . (doi:10.1007/s10336-007-0182-9)
- Limiñana , R. , Romero , M. , Mellone , U. and Urios , V. 2012a . Mapping the migratory routes and wintering areas of Lesser Kestrels Falco naumanni: new insights from satellite telemetry . Ibis , 154 : 389 – 399 . (doi:10.1111/j.1474-919X.2011.01210.x)
- Limiñana , R. , Soutullo , A. , Urios , V. and Reig-Ferrer , A. 2012b . Migration and wintering areas of adult Montagu's Harriers (Circus pygargus) breeding in Spain . J. Ornithol. , 153 : 85 – 93 . (doi:10.1007/s10336-011-0698-x)
- López-López , P. , Limiñana , R. , Mellone , U. and Urios , V. 2010 . From the Mediterranean Sea to Madagascar. Are there ecological barriers for the long-distance migrant Eleonora's falcon? . Landscape Ecol. , 25 : 803 – 813 . (doi:10.1007/s10980-010-9460-7)
- Mellone , U. , Yáñez , B. , Limiñana , R. , Román-Muñoz , A. , Pavón , D. , González , J. M. , Urios , V. and Ferrer , M. 2011 . Summer staging areas of non-breeding Short-toed Snake Eagles Circaetus gallicus . Bird Study , 58 : 516 – 521 . (doi:10.1080/00063657.2011.598914)
- Muñiz-López , R. 2007 . “ Ecología, biología y hábitat del Águila Harpía (Harpia harpyja) ” . In Cunsi Pindo. Conservación del Águila Harpía en Ecuador , Edited by: Tufiño , P. Quito : SIMBIOE .
- Muñiz-López , R. 2008 . Revisión de la situación del águila Harpía Harpia harpyja en Ecuador . Cotinga , 29 : 42 – 47 .
- O'Toole , L. T. , Kennedy , P. L. , Knight , R. L. and McEwen , L. C. 1999 . Postfledging behavior of golden eagles . Wilson Bull. , 111 : 472 – 477 .
- Palacios , W. , Cerón , C. , Valencia , R. and Sierra , R. 1999 . “ Las formaciones naturales de la Costa del Ecuador ” . In Propuesta preliminar de un sistema de clasificación de Vegetación para el Ecuador Continental , Edited by: Sierra , R. Quito : Ministerio del Medio Ambiente, Proyecto INEFAN/GEF-BIRF y EcoCiencia .
- Penteriani , V. , Otalora , F. and Ferrer , M. 2006 . Floater dynamics can explain positive patterns of density-dependent fecundity in animal populations . Am. Nat. , 168 : 697 – 703 . (doi:10.1086/507995)
- Penterani , V. and Delgado , M. M. 2009 . Thoughts on natal dispersal . J. Raptor Res. , 43 : 90 – 98 . (doi:10.3356/JRR-08-39.1)
- Penteriani , V. , Ferrer , M. and Delgado , M. M. 2011 . Floater strategies and dynamics in birds, and their importance in conservation biology: towards an understanding of nonbreeders in avian populations . Anim. Conserv. , 14 : 233 – 241 . (doi:10.1111/j.1469-1795.2010.00433.x)
- Piana , R. P. 2007 . Anidamiento y dieta de Harpia harpyja Linnaeus en la Comunidad Nativa de Infierno, Madre de Dios, Perú . Rev. Peru. Biolol. , 14 : 135 – 138 .
- Pinheiro , J. and Bates , D. 2000 . Mixed Effects Models in S and S-Plus , New York : Springer-Verlag .
- Pinheiro , P. , Bates , D. , DebRoy , S. , Sarkar , D. and the R Development Core Team . 2011 . nlme: Linear and Nonlinear Mixed Effects Models. , R package version 3.1-102
- R Development Core Team . 2011 . R: A Language and Environment for Statistical Computing , Vienna , , Austria : R Foundation for Statistical Computing .
- Rettig , N. L. 1978 . Breeding behaviour of the Harpy Eagle (Harpia harpyja) . Auk , 95 : 629 – 643 .
- Russel , E. M. and Rowley , I. 2000 . Demography and social organization of the red-winged fairy-wren Marulus elegans . Aust. J. Zool. , 48 : 161 – 200 . (doi:10.1071/ZO99066)
- Shaner , K. 2011 . Harpia harpyja , Animal Diversity Web. Available at: http://animaldiversity.ummz.umich.edu/site/accounts/information/Harpia_harpyja.html (accessed 10 April 2012)
- Skutch , A. F. 1976 . Parent Birds and Their Young , Austin : University of Texas Press .
- Soutullo , A. , Urios , V. , Ferrer , M. and Peñarrubia , S. G. 2006a . Post-fledging behaviour in Golden eagles Aquila chrysaetos: onset of juvenile dispersal and progressive distancing from the nest . Ibis , 148 : 307 – 312 . (doi:10.1111/j.1474-919X.2006.00530.x)
- Soutullo , A. , Urios , V. , Ferrer , M. and Peñarrubia , S. G. 2006b . Dispersal of Golden Eagles Aquila chrysaetos during their first year of life . Bird Study , 53 : 258 – 264 . (doi:10.1080/00063650609461441)
- Soutullo , A. , López-López , P. and Urios , V. 2008a . Incorporating spatial structure and stochasticity in endangered Bonelli's eagle's population models: implications for conservation and management . Biol. Conserv. , 141 : 1013 – 1020 . (doi:10.1016/j.biocon.2008.01.011)
- Soutullo , A. , Urios , V. , Ferrer , M. and López-López , P. 2008b . Habitat use by juvenile golden eagles in Spain . Bird Study , 55 : 236 – 240 . (doi:10.1080/00063650809461528)
- Springer , M. T. , Carver , A. D. and Correa , N. J. 2011 . Harpy Eagle (Harpia harpyja) feeding behavior on a Brown-throated Three-toed Sloth (Bradypus variegates) . J. Raptor Res. , 45 : 100 – 103 . (doi:10.3356/JRR-10-45.1)
- Strandberg , R. , Klaassen , R. H.G. , Hake , M. and Alestam , T. 2010 . How hazardous is the Sahara Desert crossing for migratory birds? Indications from satellite tracking of raptors . Biol. Lett. , 6 : 297 – 300 . (doi:10.1098/rsbl.2009.0785)
- Urios , V. , Soutullo , A. , López-López , P. , Cadahía , L. , Limiñana , R. and Ferrer , M. 2007 . First case of recruitment of a juvenile Golden Eagle (Aquila chrysaetos) tracked by satellite telemetry . Acta Ornithol. , 42 : 205 – 209 .
- Urios , V. , López-López , P. , Limiñana , R. and Godino , A. 2010 . Ranging behaviour of a juvenile Bearded Vulture (Gypaetus barbatus meridionalis) in South Africa revealed by satellite telemetry . Ornis Fenn. , 84 : 114 – 118 .
- Vargas , J. J. , Whitacre , D. , Mosquera , R. , Albuquerque , J. , Piana , R. P. , Thiollay , J. M. , Márquez , C. , Sánchez , J. E. , Lezama-López , M. , Midence , S. , Matola , S. , Aguilar , S. , Rettig , N. and Sanaiotti , T. 2006 . Estado y distribución actual del águila arpía (Harpia harpyja) en Centro y Sur America . Ornitol. Neotrop. , 17 : 39 – 55 . (doi:10.1590/S0074-02762006000100008)
- Vargas , J. J. and Vargas , F. H. 2011 . Nesting density of Harpy Eagles in Darien with population size estimates for Panama . J. Raptor Res. , 45 : 199 – 210 . (doi:10.3356/JRR-10-57.1)
- Whitfield , D. P. , Fielding , A. H. , McLeod , D. R.A. and Haworth , P. F. 2004 . The effects of persecution on age of breeding and territory occupation in golden eagles in Scotland . Biol. Conserv. , 118 : 249 – 259 . (doi:10.1016/j.biocon.2003.09.003)
- Whitfield , D. P. , Douse , A. , Evans , R. J. , Grant , J. , Love , J. , McLeod , D. R.A. , Reid , R. and Wilson , J. D. 2009 . Natal and breeding dispersal in a reintroduced population of White-tailed Eagles Haliaeetus albicilla . Bird Study , 56 : 177 – 186 . (doi:10.1080/00063650902792023)
- Young , B. E. 1996 . An experimental analysis of small clutch size in tropical house wrens . Ecology , 77 : 472 – 488 . (doi:10.2307/2265623)
- Zuur , A. F. , Ieno , E. N. , Walker , N. J. , Saveliev , A. A. and Smith , G. 2009 . Mixed Effects Models and Extensions in Ecology with R , New York : Springer .
- Zuur , A. F. , Ieno , E. N. and Elphick , C. S. 2010 . A protocol for data exploration to avoid common statistical problems . Meth. Ecol. Evol. , 1 : 3 – 14 . (doi:10.1111/j.2041-210X.2009.00001.x)