Abstract
Capsule Moult of greater coverts in the wintering area is correlated with a decrease in spring arrival condition but not arrival time of yearling male Narcissus Flycatchers.
Understanding the costs and benefits of moulting is important to understand the evolution of the moulting cycle of migrant birds. Moulting is time-consuming and energetically costly owing to the production of new feathers (Svensson & Meria Citation1996) and occurs during other extensive physiological changes (i.e. plasma volume expansion and organ hypertrophy [Ward & D'Cruz Citation1968, Chilgren & DeGraw Citation1977, Hoye & Buttemer Citation2011]). However, during moult, it is essential to maintain plumage functions such as flight ability and social communication (Swaddle & Witter Citation1997). Moulting pattern is especially important for migrating species because the moulting of wing feathers affects flying performance (Ginn & Melville Citation1983).
It is well known that the energy requirements of moulting conflict with the life-cycle strategy of migrants. Simultaneous moulting and breeding has been shown to reduce reproductive success (Hemborg & Lundberg Citation1998, Hemborg Citation1999, Hemborg et al. Citation2001), but delaying post-nuptial moult may decrease feather quality (Dawson Citation2004). In some species, individuals that complete post-juvenile or post-breeding moult earlier store more fat than those in which moult and the autumn migration overlap (Merilä Citation1997). In contrast, the trade-off between pre-breeding moult in the wintering area and spring migration is not yet well understood.
The pre-breeding moult of some migrating species (e.g. the Pied Flycatcher Ficedula hypoleuca and some Motacillidae) in the wintering area involves body plumage and tertials (Ginn & Melville Citation1983). Pre-breeding moulting appears to have evolved for social reasons (i.e. male–male competition and sexual selection by females [Owens et al. Citation1994, Battley et al. Citation2006]), where low-quality plumage obtained during the fast summer moult is replaced (Hedenström et al. Citation1995), especially juvenile feathers (de la Hera et al. Citation2010). This would also be an advantage for migration, since migration with worn feathers is associated with a high cost of flight (Salewski et al. Citation2004). However, accumulation of energy for spring migration at the same time in the wintering area is also necessary because body condition affects arrival time at the breeding site, annual survival, and reproductive success (Alatalo et al. Citation1984, Takaki et al. Citation2001, Saino et al. Citation2004, Mitrus Citation2007). Therefore, individuals face the problem of how to use their energy for both pre-breeding moult and fat accumulation for migration, as well as the possible cost of a pre-breeding moult on spring migration.
If there is a trade-off between the pre-breeding moult and spring arrival condition, birds that moult only a few feathers might be in good condition when they arrive at the breeding area. They might then arrive earlier because of their good condition. Here we evaluate the hypothesis that the pre-breeding moult affects spring arrival condition and arrival date of yearling male Narcissus Flycatchers Ficedula narcissina.
The Narcissus Flycatcher is a sexually dimorphic small passerine which breeds in northeast Asia and winters in southeast Asia (del Hoyo et al. Citation2006, Töpfer Citation2006). Narcissus Flycatchers have a partial moult of body feathers, greater, median, and lesser coverts, and some tertials in spring. They also moult completely in autumn post-breeding (Yamashina Citation1941). The summer plumage colour of yearling males and adults thus differs significantly. The entire upper part of the adult male wing feather is black, and they have white patches on their inner greater coverts and sometimes also on their tertials. The primaries, secondaries, tertials, greater coverts, medium coverts, lesser coverts and alula of yearling males have female-type brown juvenile feathers, prior to any spring moult. Any greater coverts and tertials of yearling males moulted in the wintering area are black with white patches as in adult male feathers (Kuroda Citation1925, Yamashina Citation1941). Therefore, we can easily determine whether each feather of the first summer plumage has been moulted in the wintering area or not.
Data were collected from 15 April to 15 May in 2010, 2011 and 2012 in the study area at Fuji Primitive Forest, central Japan (35°27′N, 138°38′E; 60 ha; 1140 m above sea level). We searched for singing males in the study area every morning (03:30–11:30) during the study. The day that a male was first recorded singing in a territory was assumed to be his arrival day (Mitrus Citation2007). In subsequent statistical analyses to examine the effect of moulting on arrival time, Julian date from 25 April was used because the first flycatcher was observed on 25 April. Each male was lured into mist nets using song playback for up to two weeks from his arrival day, and captured an average of 2.71 ± 4.43 sd days after his arrival within his territory. The age of each bird was determined by plumage characteristics. We recorded the moulting status (feather moulted in the wintering area or juvenile) of each greater covert, primary, secondary, and tertiary.
The following measurements were taken: body mass using a 30 g Pesola spring balance (to the nearest 0.25 g [Mitrus 2007]), wing length (maximum wing chord in mm [Mitrus 2007]), and tarsus length with Mitsutoyo sliding digital callipers (to the nearest 0.1 mm [Svensson Citation1992]). An index of individual body condition was calculated using the residual from the regression of body mass on tarsus length (Veiga Citation1993, Merilä Citation1996, Mitrus Citation2007). Males were uniquely ringed using a combination of aluminium (authorized by the Japan Environment Agency) and colour-coded rings.
All statistical tests were carried out using R version 2.12.2. (R development core team Citation2011). To examine the effect of pre-breeding moult on arrival body condition, we used general linear models (GLMs) with Gaussian error and identity link function. P-values for each factor were obtained from a likelihood F-test of full models. We used the number of moulted greater coverts, the number of moulted tertials, wing length and study year as explanatory variables. Study year was included as a factor because of annual variability of arrival condition; other predictors were continuous.
We also used GLMs with Poisson error and log link function to reveal the effect of pre-breeding moult on arrival date. In this case, P-values were obtained from a likelihood Chi-squared test, and we included the number of moulted greater coverts, the number of moulted tertials, body condition, wing length, and study year as possible predictors. Study year was included as a factor because of annual variability of arrival condition, other predictors were continuous. In both cases, non-significant variables were removed from the final model, but their presence in full models did not change the results.
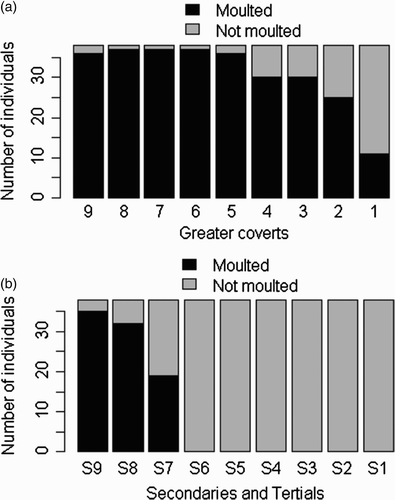
We captured 38 yearlings over 3 years and found individual variability in the pre-breeding moult of Narcissus Flycatchers. Most flycatchers moulted their inner greater coverts, but the number of moulted outer greater coverts was lower than that of inner ones (. The flycatchers never moulted their primaries and outer secondaries, but some individuals moulted their tertials (S7–S9; .
Figure 1. Moulting status of Narcissus Flycatchers. (a) Greater coverts, (b) Secondaries, and Tertials.
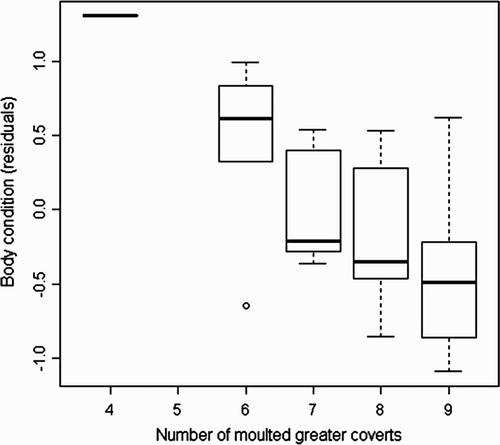
The number of moulted greater coverts had a significant negative effect on arrival body condition; however, the number of moulted tertials had no such effect (): GLM, study year coefficient = –0.38 ± 0.10 se, F 1,37 = 13.1, P < 0.01; wing length coefficient = 0.03 ± 0.06 se, F 1,36 = 0.2, P = 0.66; number of moulted greater coverts coefficient = –0.19 ± 0.07 se, F 1,35 = 8.8, P < 0.01; number of moulted tertials coefficient = 0.05 ± 0.11 se, F 1,34 = 0.2, P = 0.64. In contrast, the moulting status of wing feathers did not affect arrival time: glm, study year coefficient = –4.19 ± 0.85 se, F 1,37 = 26.2, P = 0.001; wing length coefficient = –0.04 ± 0.39 se, F 1,36 = 0.4, P = 0.56; number of moulted greater coverts coefficient = 0.34 ± 0.52 se, F 1,35 = 0.5, P = 0.51; number of moulted tertials coefficient = –1.05 ± 0.79 se, F 1,34 = 2.0, P = 0.17; body condition coefficient = –1.50 ± 1.21 se, F 1,33 = 1.5, P = 0.23).
Figure 2. The number of moulted greater coverts by body condition (residuals from tarsus length on body weight) at arrival. Thick horizontal lines represent medians, boxes represent upper and lower quartiles, and fine lines extending from boxes indicate extremes (maximum and minimum data in 1.5 times the interquartile range, which is the distance between the lower and upper quartiles of the data). Small circles indicate outliers.
Salewski et al. Citation(2004) suggested that moulting tertials and innermost secondaries in the spring would convey an advantage for the migration of Pied Flycatcher because these feathers are the most exposed, and become worn faster than other remiges. If there is a high energy cost of migration associated with worn feathers, body condition or arrival time should be positively correlated with the number of moulted feathers on birds' wings. However, our results suggest that migration of Narcissus Flycatchers with worn feathers may not be associated with a high cost but there may be an energy cost of pre-breeding moult on spring arrival condition. In addition, moulting of primaries is associated with high energy costs for birds (Barta et al. Citation2008) and, therefore, flycatchers may retain their old primaries to cut the energetic costs of moulting.
There is a possibility that the trade-off we observed was not caused by physiological factors, but by social factors, for example, male–male competition in Narcissus Flycatchers. Yearling males that moulted their tertials and greater coverts, and thus have larger white wing patches, may be exposed to more aggression and fights, which may reduce their body condition. Therefore, yearling males that moulted fewer feathers could be in better condition owing to avoidance of aggression and competition from males during migration.
In the present study we found a negative effect of pre-breeding moult on arrival condition but no effect of body condition on arrival time. Studies of post-breeding moulting of migratory birds have shown that moulting of feathers is associated with lower body condition (Merilä Citation1997, Portugal et al. Citation2007). Other studies have shown that early-migrating individuals moult all feather types faster than late-migrating ones (Jenni & Winkler Citation1994, Kjellén Citation1994), and individuals in better condition arrive at the breeding grounds faster than worse condition ones (Mitrus Citation2007). Our contradictory results may thus have arisen because we examined only yearling males. The lack of a relationship between the arrival time of yearling males and body condition has also been observed in the Red-breasted Flycatcher F. parva (Mitrus Citation2007) and in the dusky warbler Phylloscopus fuscatus (Forstmeier Citation2002). These studies also suggested condition-dependent arrival may vary with environmental conditions. For example, in a year of good weather conditions, no relationship between the arrival date and body condition was found (Ninni et al. Citation2004). Moreover, young males that are attempting to breed for the first time may need more time to find high-quality territories (Mitrus Citation2007). Thus, environmental conditions and delayed acquisition of a territory might mask the effect of pre-breeding moult and body condition on spring arrival time.
An interesting finding is that flycatchers still moulted some of their greater coverts, even though this was associated with a poorer spring arrival condition. This might mean that moulting these feathers carries a benefit for flycatchers, even though such sexual trait expression probably decreases body condition (Balmford et al. Citation1993, Mennill et al. Citation2003, Hegyi et al. Citation2010). The white patch on the wing is important for sexual selection and intra-sexual competition in Ficedula flycatchers (Sirkiäa & Laaksonen Citation2009), and mature male Narcissus Flycatchers and yearling males that have moulted have a white patch on their greater coverts and tertials. Therefore, having a larger number of moulted greater coverts and tertials might be important for competition in the breeding area and this requires future study.
ACKNOWLEDGEMENTS
We thank Ayaka Sasaki, Hiromi Konishi, and Narisa Togo for assistance in the field.
REFERENCES
- Alatalo , R. V. , Gustafsson , L. and Lundberg , A. 1984 . Why do young birds have shorter wings than older birds? . Ibis , 126 : 410 – 415 .
- Balmford , A. , Thomas , A. L.R. and Jones , I. L. 1993 . Aerodynamics and the evolution of long tails in birds . Nature , 361 : 628 – 631 .
- Barta , Z. , McNamara , J. M. , Houston , A. I. , Weber , T. P. , Hedenström , A. and Feró , O. 2008 . Optimal moult strategies in migratory birds . Philos. Trans. R. Soc. Lond., B, Biol. Sci. , 363 : 211 – 229 .
- Battley , P. F. , Rogers , D. I. and Hassel , C. J. 2006 . Prebreeding moult, plumage and evidence for a presupplemental moult in the Great Knot Calidoris tenuirostris . Ibis , 148 : 27 – 38 .
- Chilgren , J. D. and DeGraw , W. A. 1977 . Some blood characteristics of White-crowned Sparrows during molt . Auk , 94 : 169 – 171 .
- Dawson , A. 2004 . The effects of delaying the start of moult on the duration of moult, primary feather growth rates and feather mass in Common Starlings Sturnus vulgaris . Ibis , 146 : 493 – 500 .
- de la Hera , I. , Díaz , J. A. , Pérez-Tris , J. and Tellerría , J. L. 2010 . Relationships among timing of moult, moult duration and feather mass in long-distance migratory passerines . J. Avian Biol. , 41 : 609 – 614 .
- del Hoyo , J. , Elliott , A. and Christie , D.A. 2006 . Handbook of the Birds of the World 11. Old World Flycatchers to Old World Warblers Edited by: del Hoyo , J. , Elliott , A. and Christie , D.A. Lynx Edicions, Barcelona
- Forstmeier , W. 2002 . Benefits of early arrival at breeding grounds vary between males . J. Anim. Ecol. , 71 : 1 – 9 .
- Ginn , H. B. and Melville , D. S. 1983 . Moult in Birds , Tring, UK : British Trust for Ornithology .
- Hedenström , A. , Lindström , Å. and Pettersson , J. 1995 . Interrupted moult of adult Willow Warblers Phylloscopus trochilus during autumn migration through Sweden . Ornis Svec. , 5 : 69 – 74 .
- Hegyi , G. , Szöllősi , E. , Jenni-Eiermann , S. , Török , J. , Eens , M. and Garamszegi , L. Z. 2010 . Nutritional correlates and mate acquisition role of multiple sexual traits in male collared flycatchers . Naturwissenschaften , 97 : 567 – 576 .
- Hemborg , C. 1999 . Sexual differences in moult-breeding overlap and female reproductive costs in pied flycatchers, Ficedula hypoleuca . J. Anim. Ecol. , 68 : 429 – 436 .
- Hemborg , C. and Lundberg , A. 1998 . Costs of overlapping reproduction and moult in passerine birds: an experiment with the pied flycatcher . Behav. Ecol. Sociobiol , 43 : 19 – 23 .
- Hemborg , C. , Sanz , J. J. and Lundberg , A. 2001 . Effects of latitude on the tradeoff between reproduction and moult: a long-term study with pied flycatcher . Oecologia , 129 : 206 – 212 .
- Hoye , B. J. and Buttemer , W. A. 2011 . Inexplicable inefficiency of avian molt? Insights from an opportunistically breeding arid-zone species . Lichenostomus penicillatus. PLoS ONE , 6 : e16230
- Jenni , L. and Winkler , R. 1994 . Moult and Ageing of European Passerines , London : Academic Press .
- Kjellén , N. 1994 . Moult in relation to migration in birds – a review . Ornis Svec. , 4 : 1 – 21 .
- Kuroda , N. 1925 . A Contribution to the Knowledge of the Avifauna of the Riu Kiu Islands and Vicinity , Tokyo : N. Kuroda .
- Mennill , D. J. , Doucet , S. M. , Montgomerie , R. and Ratcliffe , L. M. 2003 . Achromatic colour variation in black-capped chickadees, Poecile atricapilla: black and white signals of sex and rank . Behav. Ecol. Sociobiol. , 53 : 350 – 357 .
- Merilä , J. 1996 . Genetic variation in offspring condition: an experiment . Funct. Ecol. , 10 : 465 – 474 .
- Merilä , J. 1997 . Fat reserves and moult–migration overlap in goldcrests, Regulus regulus – a tradeoff? . Ann. Zool. Fenn. , 34 : 229 – 234 .
- Mitrus , C. 2007 . Is the later arrival of young male Red-breasted Flycatchers (Ficedula parva) related to their physical condition? . J. Ornithol. , 148 : 53 – 58 .
- Ninni , P. , de Lope , F. , Saino , N. , Haussy , C. and Møller , A. P. 2004 . Antioxidants and condition-dependence of arrival date in a migratory passerine . Oikos , 105 : 55 – 64 .
- Owens , I. P.F. , Burke , T. and Thompson , D. B.A. 1994 . Extraordinary sex roles in the Eurasian dotterel: female mating arenas, female–female competition, and female mate choice . Am. Nat. , 144 : 76 – 100 .
- Portugal , S. J. , Green , J. A. and Butler , P. J. 2007 . Annual changes in body mass and resting metabolism in captive barnacle geese (Branta leucopsis): the importance of wing moult . J. Exp. Biol. , 210 : 1391 – 1397 .
- R development core team . 2011 . R: a language and environment for statistical computing , Version 2.12.2. R Foundation for Statistical Computing, Vienna
- Saino , N. , Szép , T. , Romano , M. , Rubolini , D. , Spina , F. and Møller , A. P. 2004 . Ecological conditions during winter predict arrival date at the breeding quarters in a trans-Saharan migratory bird . Ecol. Lett. , 7 : 21 – 25 .
- Salewski , V. , Altwegg , R. , Erni , B. , Falk , K. H. , Bairlein , F. and Leisler , B. 2004 . Moult of three Palaearctic migrants in their West African wintering quarters . J. Ornithol. , 145 : 109 – 116 .
- Sirkiäa , P. M. and Laaksonen , T. 2009 . Distinguishing between male and territory quality: females choose multiple traits in the pied flycatcher . Anim. Behav. , 78 : 1051 – 1060 .
- Svensson , E. and Meria , E. 1996 . Moult and migratory condition Blue Tits: a serological study . Condor , 98 : 825 – 831 .
- Svensson , L. 1992 . Identification Guide to European Passerines , 4 , Tring, UK : British Trust for Ornithology .
- Swaddle , J. P. and Witter , M. S. 1997 . The effects of moult on the flight performance, body mass, and behavior of European starlings (Sturnus vulgaris): an experimental approach . Can. J. Zool. , 75 : 1135 – 1146 .
- Takaki , Y. , Eguchi , K. and Nagata , H. 2001 . The growth bars on tail feathers in the male Styan's Grasshopper Warbler may indicate quality . J. Avian Biol. , 32 : 319 – 325 .
- Töpfer , T. 2006 . Systematic notes on Asian birds. 60. Remarks on the systematic position of Ficedula elisae (Weigold, 1922) . Zool. Med. Leiden , 80 : 203 – 212 .
- Veiga , J. P. 1993 . Badge size, phenotypic quality and reproductive success in the house sparrow: a study on honest advertisement . Evolution , 47 : 1161 – 1170 .
- Ward , P. and D'Cruz , D. 1968 . Seasonal changes in thymus gland of a tropical bird . Ibis , 110 : 203 – 205 .
- Yamashina , Y. 1941 . [A Natural History of Japanese Birds 2] , Tokyo (in Japanese) : Iwanami-shoten .