Abstract
Capsule European Nightjars Caprimulgus europaeus breeding in southern England were found to over-winter in the Democratic Republic of Congo.
Aims To ascertain the wintering areas and migration routes of European Nightjars breeding in southern England.
Methods The wintering areas of three Nightjars were mapped using light geolocation tags (two in 2008 and one in 2010). For one of these birds, details of the timing and route of migration were determined. The impact of the birds' behaviour on location accuracy was measured and data on the timing of emergence and roosting was collected.
Results All three Nightjars were found to be wintering in the south and east of the Democratic Republic of Congo, in an area not previously considered to be part of the wintering range of this species. The route of migration differed in each period. Autumn migration was across central Sahara, whereas in spring the route was to the west of the Sahara. Aberrations in the light curve caused by the roosting and emergence of the birds were found to affect the estimated location of the wintering areas, shifting them approximately 1° south, and reducing the estimated accuracy of the locations. The timing of these aberrations showed that roosting and emergence roughly follow the timing of dawn and dusk.
Conclusions Current distribution maps for the wintering areas of Nightjars in Africa probably under-represent the true distribution of the species in the continent. The wide dispersal of birds from the same breeding area in the UK may be an indication of mixing of breeding populations during the wintering period. Further study is needed to understand how these results fit into the larger picture of Nightjar migration both from the UK and the wider Eurasian breeding range, and to determine locations of stopovers.
The European Nightjar (Caprimulgus europaeus) is the UK's only native member of the Caprimulgidae. Nightjars breed mainly in conifer plantation clearings and heaths throughout the UK, with principle population centres in southern and eastern England. While research into the behaviour of Nightjar during the breeding season has provided many insights into both foraging and mating behaviour (Alexander & Cresswell Citation1990, Cresswell & Alexander Citation1990, Citation1992, Cresswell Citation1996), very little is known about the migration and wintering period. Current maps show the possible wintering range to extend along the eastern coast of Africa from Kenya to South Africa and in the western Sub-Saharan region from Senegal to Cameroon (Cleere & Nurney Citation1998, Holyoak Citation2001). However, recovery rates of ringed individuals within Africa are extremely low and data to confirm current distribution maps and migratory routes for the continent are largely lacking (Wernham et al. Citation2002). This problem of a lack of knowledge on migration routes and wintering areas is common across a variety of species and may be a block to successful conservation management and allocation of scarce funds (Luschi Citation2003, Silvano et al Citation2006, Faaborg et al Citation2010, Robinson et al. Citation2010, Kirby Citation2011).
Miniaturized light-geolocation tags are one of a range of options available in the study of migration, allowing long-term tracking of species that is impossible using traditional radiotracking techniques (Fiedler Citation2009). These tags have been deployed successfully in a number of studies investigating long-distance migration and the locations of wintering areas (Croxall et al. Citation2005, Shaffer et al. Citation2006, González-Solís et al. Citation2007, Harris et al. Citation2009, Rodríguez et al. Citation2009, Stutchbury et al. Citation2009). Light-geolocation tags monitor light intensity at regular intervals and use the timing of dawn and dusk to estimate longitude and the length of daylight to estimate latitude (Wilson et al. 2002, Hill Citation1994). Light-geolocation shows greater accuracy in longitude estimates than latitude due to sensitivity to small changes in day length produced by errors in placing the timing of dawn and dusk. Latitude estimation is also impossible during the periods around the spring and autumn equinox because day length is the same at all latitudes (Hill Citation1994, Welch & Eveson Citation1999, Musyl et al. Citation2001). Although geolocation using light levels is not a new idea, it is only relatively recently that use for tracking animal movements has become widespread, mainly due to miniaturization of the devices. Deployment has been largely restricted to marine animals, for whom sea-surface temperature (SST) data can be used to compensate for the inherently poor latitude estimates from light data alone (Teo et al. Citation2004, Shaffer et al. Citation2005, Nielsen et al. 2006). Despite the limitations on the accuracy of latitude estimates, the low weight of these miniaturized tags is a major benefit in the long-term tracking of small birds for which larger GPS or satellite tags are not suitable, as shown in a number of recent studies on small terrestrial species (Rodriguez et al. 2009, Stutchbury et al. Citation2009, Bächler et al. Citation2010).
Most studies to date have used a light-intensity threshold to identify the timing of dawn and dusk (Shaffer et al. Citation2005, Fuller et al. Citation2008, Harris et al. Citation2009, Rodríguez et al. Citation2009). A more sophisticated approach is to fit an idealized curve, created from theoretical calculations, to the light-intensity data as a means of identifying the timing of dawn and dusk. The slope of this fitted curve, in addition to day length and the timing of dawn and dusk, is used to make the location estimates (Ekstrom Citation2002, Citation2004). This ‘template-fit’ approach has a number of advantages to the threshold method. The template does not need to be ‘tuned’ to the data and therefore removes the subjectivity that comes with selecting a light-intensity threshold, and allows separate fitting of the curve for dawn and dusk to accommodate changes in cloudiness across the day. Another advantage to the template-fit method is that it provides an estimate of accuracy of individual locations by assessing the goodness-of-fit to the theoretical twilight curves (Ekstrom Citation2007). Test data from stationary tags and data from the breeding period have shown the latitude error estimate to be linearly related to the true error, although the strength of this relationship appears to change over time (Cresswell & Edwards unpubl. data). During the equinox period this relationship is decoupled and the error estimate does not reflect the true error.
This study aimed to:
| 1. | provide new data on the location wintering grounds of Nightjar breeding in the UK and the route and timings of the pre- and post-nuptial migration using miniaturized light-based geolocation tags; and | ||||
| 2. | assess the utility of ‘template-fit’ location estimation for these tags. | ||||
METHODS
Nightjars were captured using mist-nests and playback of their calls in Wareham Forest, Dorset, southern England in late summer and early autumn of 2008 (five birds, all males) and 2010 (five males and one female). The birds were fitted with geolocation tags (LightBugs) from Biotrack Ltd (Wareham, Dorset BH20 5AX, UK), under licence from the British Trust for Ornithology. The tags used in 2008 were prototypes weighing 2 g with 60-mm flexible light stalks, while those in 2010 were 1.8 g commercially available LightBugs with 15-mm rigid light stalks. In both designs the light stalks contained light fibre that channelled light to a sensor within the tag. The longer stalks used in 2008 gathered more light. Tags were attached with a full-body harness (Kenward et al. Citation2001) made from 4-mm Teflon ribbon (Bally Ribbon Mills, PA 19503, USA). They also contained VHF transmitters to improve chances of recapture upon the birds' return to the UK. Light intensity was measured every 4 minutes in the 2008 tags and every 7 minutes, 12 seconds in the 2010 tags. Of the 11 tags deployed, 3 (27%) were retrieved (all from males), Return rates from the study site for ringed-only birds between 2011 and 2012 was 14% (4 of 28). Therefore, we believe the return rate of instrumented birds to be acceptable for this site.
Tag data were downloaded using TagTalk software (Lotek Wireless Inc., L3Y 7B5, Canada). Template-fit analysis (Ekstrom Citation2004) of the raw light data was carried out using LAT Viewer Studio 2.9.36 (Lotek Wireless Inc.) to create daily location estimates. As in previous studies (Guilford et al. Citation2009, Rodríguez et al. Citation2009, Stutchbury et al. Citation2009) the periods when the birds were in the breeding area, wintering area and on migratory stopovers were taken to be periods of stable longitude. A bird was considered to be migrating when the longitude or latitude of three or more locations was shifted by more than twice the standard deviation from either the winter or summer means. Latitude values were excluded for 20 days either side of the equinoxes, from 3 September to 13 October and from 28 February to 9 April. During these periods day length is too similar at all latitudes, making latitude estimation unreliable. Timings of each period of movement were recorded and used to assess the duration and speed of the migration. During the periods when the birds were not actively migrating, i.e. periods with stable longitude, it was assumed that any movement by the birds would be inconsequential in comparison with the magnitude of error associated with geolocation. Given this assumption, all variation in location estimates was taken to be due to random error in the estimates and therefore the distribution was expected to fit to a normal distribution in both the x- (easting) and y- (northing) axes.
Each of the stationary periods was analysed using home range analysis in Ranges8 v2.2 (Kenward et al. Citation2008). The latitude and longitude estimates produced by LAT Viewer Studio were converted into northing and easting values for a Mercator projection with a false northing of 13 523 000 m and a false easting of 1 403 000 m using GEOTRANS 2.4.2 (US National Geospatial Intelligence Agency). This conversion was necessary because Ranges cannot analyse angular Lat/Lon coordinates or data with negative values. Before carrying out home-range analysis the locations were filtered to remove outliers. This filtering was carried out automatically using the latitude error estimate based upon the goodness of fit of the template that is created as part of the template fit analysis (Ekstrom Citation2007). For the 2008 tags a cut-off latitude error of 4.9° was chosen as this removed all outliers for tag 766 and all but one outlier for tag 757. The choice of cut-off error value for the 2010 tag (125) was more difficult due to a higher average error and a more even spread across the error range, which was caused by the shorter light stalk in this tag. Subsets of the data with varying error thresholds were analysed to assess the accuracy of the location estimate during the breeding period and also the size of ellipses for both breeding and wintering periods, plotted in a similar way to utilisation curves (Kenward et al. Citation2008). From this assessment, a cut-off of latitude error of 7° was chosen to allow the best compromise between the accuracy and size of the data set. The assumption that all variation was due to random error was tested by carrying out Kolmogorov–Smirnov tests for normality on both easting and northing values for the filtered datasets using the nortest package in R (R Development Core Team Citation2010). Simple ellipses were used to assess the location of the wintering areas because the observed variation was assumed to be due to random error and expected to fit a normal distribution (Kenward Citation2001). Distances between the wintering areas, potential stopovers and the breeding area were calculated using great circle distances using the centre points of the ellipses and the point of capture, respectively, and assuming a spherical earth of radius 6372.8 km.
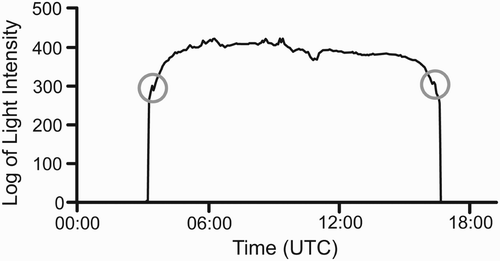
During visual examination of the light curves of tags 757 and 766, a pattern of aberrations in the light curves at dawn and dusk were observed, likely caused by the behaviour of the bird entering and leaving cover; see example curve in . These aberrations would have introduced a bias in the locations produced by template-fit methods by dropping the daytime light levels and reducing the slope of the fitted curve, leading to a poleward transformation in location estimates (Ekstrom pers. comm.). The aberrations were identified visually using LAT Viewer Studio and the timing of the aberrations along with the temperature and light intensity immediately prior to the aberration were collated manually from the raw data. The raw data were then transformed manually in Excel (Microsoft Corp.) by lowering light readings on the night-side of the aberration to remove the impact on template-fit location. The light intensity at the peak of the aberration was decreased such that the slope remained constant with that immediately to the day side of the aberration. All light-intensity values to the night side of the peak were decreased by the same amount as the peak until the curve flattened or reached the minimum sensitivity of the light sensor. Only days with a single aberration in the light curve at dawn and/or dusk were subject to this transformation, days with multiple or no aberrations were not transformed. The data set transformed to remove the aberrations was then analysed using the programme Template Fit Beta Version 7 (Lotek Wireless Inc.) and then transformed, filtered and analysed as above. The timing between the aberrations, thought to represent the time the bird was active, was plotted against a number of variables that might explain variation in the length of the activity period. The variables chosen were potential surrogates for prey availability. These were: average night-time temperature over 1–5 nights and average normalized daytime light level, both of which might be expected to influence the activity of insect prey. The proportion of the night illuminated by the moon was also considered, because Nightjars are known to synchronize their breeding with moon phase (Perrins & Crick Citation1996), indicating moonlight may be important to them.
Figure 1. Light-intensity curve showing aberrations at dawn and dusk (circled) that we believe occur as a result of the birds' roosting behaviour. The light scale is arbitrary because the measurements are not calibrated.
RESULTS
Wintering area
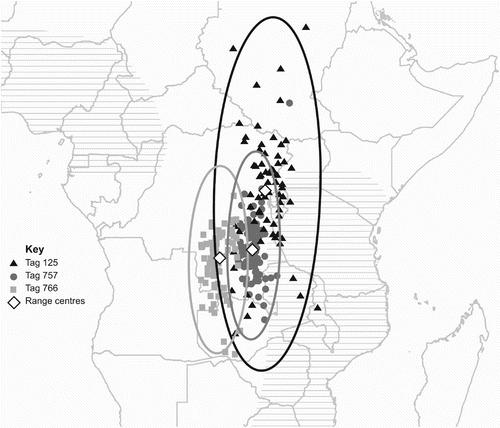
Usable data were downloaded from all three of the retrieved tags, two from the 2008 deployment (tags 757 and 766) and one from the 2010 deployment (tag 125). The tags from the 2008 deployment contained raw light-intensity data from 15 October 2008 until 26 January 2009, after which date they had been programmed to calculate locations on-board due to insufficient memory for further raw data to be stored. The tag from 2010, which had a slower sampling rate, contained raw data for the entire duration of its deployment, from 29 July 2010 to 28 June 2011. The arrival date into the wintering areas was estimated for the three birds as 24 October 2008, 20 October 2008 and 29 October 2010 for tags 757, 766 and 125, respectively. Range analysis of the locations for the wintering period place the wintering range of the two birds tagged in 2008 in the southeast of the Democratic Republic of Congo (DRC) (6.73°S, 27.48°E for tag 757, and 8.12°S, 24.07°E for tag 766) and the bird tagged in 2010 within central eastern DRC, near to the borders with Rwanda and Burundi (1.68°S, 28.83°E); these data can be seen in . The centre of the ellipses for the 2008 birds both fell within the Central Zambezian Miombo woodlands, although for tag 766, the centre is close to the boundary with the Southern Congolian forest savanna mosaic (Olson et al. Citation2001). The centre of the ellipse for the 2010 bird was found in the Albertine Rift montane forests (Olson et al. Citation2001), although this is a relatively small ecoregion in comparison to the size of the ellipse and the wintering area could easily fall in any of the surrounding ecoregions. The locations for tags 125 and 757 both conform to the assumption of normality in both easting and northing, but the data from tag 766 do not conform to normality in either axis showing a northerly and easterly bias (Kolmogorov-Smirnov test for normality: tag 757 easting D = 0.0845, n = 82, P = 0.158, northing D = 0.0843, n = 82, P = 0.160; tag 766 easting D = 0.1271, n = 83, P = 0.002, northing D = 0.1429, n = 83, P = 0.0002; tag 125 easting D = 0.0763, n = 75, P = 0.34, northing D = 0.0769, n = 75, P = 0.33).
Figure 2. Map of Central Africa showing the wintering period for all three birds. Filtered locations for each bird are shown with 99% ellipses and the centres of the ranges are indicated by white diamonds: 125 by black triangles, 757 by dark grey dots and 766 by pale grey squares. The extent of the wintering ground as previously believed is shown by horizontal hatching.
Timing and route of migration
With nearly a full year of data collected in tag 125 it was possible to estimate the timing and route for the whole of the migration to and from the wintering areas, as shown in . Because individual latitude estimates were often poor, especially near to the equinoxes, daily positions of the migrating bird are not shown. Instead, only longitude data have been used, shown as maximum and minimum (range) of longitude in successive 9–10-day periods (except from 11 April to 4 May when successive 10-day periods overlapped almost entirely).
Figure 3. Map showing ranges of longitude occupied by bird 125 for successive 10-day periods during post- and pre-nuptial migration. Between 11 April and 4 May, successive 10-day periods overlapped; therefore, this band includes 24 days/locations. The length of the vertical lines (longitude) indicates realistic latitudinal limits and not the range of the latitudes in each period, which were much greater. Darker lines and text in two longitude ranges are to make the figure clearer. Numbers of days/locations in each longitude band are shown in brackets. Probable routes are suggested by broad arrows between breeding and wintering sites.
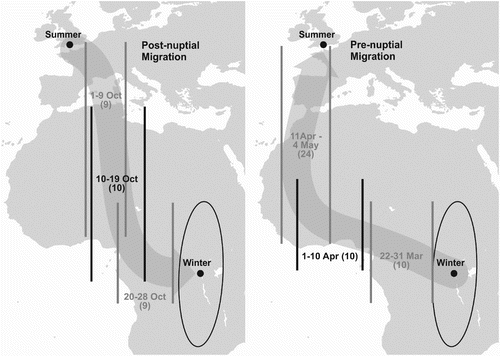
This bird started its post-nuptial migration around 1 October 2010, taking a route across the Mediterranean, and then across the central Sahara. It is possible that the bird stopped over during mid-October 2010, perhaps at more than one site, but as no locations during this time had an estimated latitude error lower than the cut-off value of 7°, it would be too speculative to suggest stopover locations. Ignoring possible stopovers, the minimum distance travelled per day using the straight line distance between the breeding and wintering grounds was 232 km/day.
Nightjar 125 remained in its wintering area until 22 March 2011 when it began its pre-nuptial migration, travelling 2700 km west in 12 days (about 270 km per day). It took about another 32 days to travel a further 6000 km to its breeding area in the UK. This probably included one or more stopovers in West Africa, but because only five locations during this time had a latitude error lower than 7°, the data are not adequate to identify sites. The bird returned to the UK via the western Sahara and Spain, arriving at its breeding grounds on 6 May 2011. Its average speed through West Africa was about 190 km per day, which is a very conservative estimate of a typical daily movement if this period included stopovers.
Impact of aberrations in the light curve
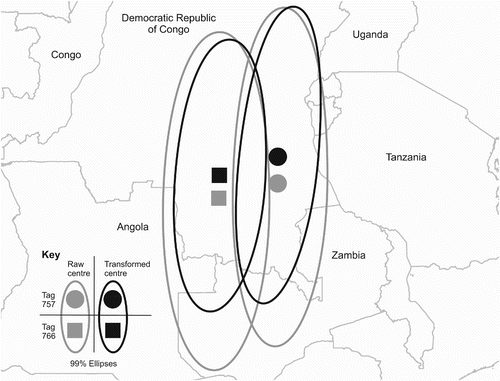
Aberrations were often present in the light curves at dawn and dusk. As shows, the dusk aberration took the form of a sudden jump in intensity during a steep section of the declining light curve, and was thought to be caused by a bird emerging from its roost into the open sky. The aberration was mirrored at dawn, when the light level dropped suddenly during a steeply rising section of the light curve. This is believed to be caused by the Nightjar retiring to its roost. The light curves of 74 dates were transformed by the removal of aberrations at either dawn or dusk for tag 757, of which 29 were transformed at both dawn and dusk. Thirteen dates had no aberration at dawn or dusk and one date had multiple aberrations at both dawn and dusk: these dates received no transformation. For tag 766, 79 dates were transformed, of which 44 dates were transformed at both dawn and dusk. Fourteen dates did not have any aberrations and received no transformation. The removal of the aberrations at dawn and dusk moved the centre of the ellipses northwards for both tags, to 5.60°S, 27.48°E for tag 757 and 6.76°S and 24.25°E, as shown in . This is a movement in latitude of 1.13° for tag 757 and 1.36° for tag 766. This transformation also reduced the overall variation in latitude which can be seen in the smaller ellipses produced for this data set. As would be expected, the removal of these aberrations at dawn and dusk improved the fit of the template curve leading to lower estimated error in the locations. This transformation brought the distribution of points for tag 766 closer to normality in the x-axis and successfully normalized the y-axis (b. easting D = 0.0962, n = 83, P = 0.04, northing D = 0.0922, n = 83, P = 0.06). Tag 125 did not show aberrations, which were probably masked by higher noise levels in the light data due to the shorter light stalk.
Figure 4. Map of Central Africa showing the impact of aberrations in the light curve at dawn and dusk on the placement of 99% ellipses to ascertain the wintering area. Ellipses and their centres derived from raw data are light grey and those derived from transformed data are black. Centres of ellipses for bird 757 are circles and for bird 766 are squares.
Activity periods of Nightjars
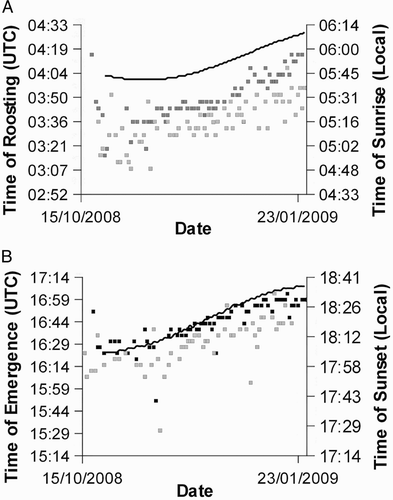
Aberrations at dusk were found on 58 occasions for tag 757 and 71 occasions for tag 766. Aberrations at dawn were found on 71 occasions for tag 757 and 73 occasions for tag 766. From these indicators of arising from and retiring to roost it was possible to calculate the period of activity during 41 nights for tag 757 and 50 nights for tag 766. Plotting the timing of the aberrations showed that the birds' emergence from roost became steadily later over time, broadly in line with changes in the timing of sunset for the region. The timing of roosting also showed a trend broadly in line with the timing of sunrise; both these trends can be seen in . There was no overall trend in total activity time over the course of the wintering period and the mean times active for the two birds showed remarkable similarity, 11 hours 6 minutes 37 seconds (se 14 minutes 20 seconds) and 11 hours 6 minutes 47 seconds (se 11 minutes 30 seconds) for 757 and 766, respectively. None of the surrogates for prey availability (night-time temperature, average normalized daytime light level and proportion of the night illuminated by the moon) were able to describe the variation seen in overall activity times. (anova: night-time temperature: F1,75 = 0.9, P = 0.36; daytime light level: F1,75 = 0.8, P = 0.39; moonlight: F1,75 = 0.1, P = 0.71). Of the five measures of night-time temperature (average temperature over 1–5 nights) we used the average over 3 nights because this best correlated with each of the other measures (Spearman's ρ with 1 night = 0.79, 2 nights = 0.94, 4 nights = 0.91, 5 nights = 0.96) and because biologically this best represents both long-term and short-term temperature.
Figure 5. Plots of the timing of roosting (A) and emergence (B) (UTC) and the timing of sunrise and sunset (solid lines) in the general region of the wintering ranges (local time). Black squares represent bird 766 and grey squares bird 757. Emergence and roosting times of bird 757 were earlier because it was further east than bird 766.
DISCUSSION
The winter locations of these three Nightjars suggest that the British breeding population spreads out widely during the wintering period. Each of the birds in this study were located within 1 km of each other when caught in their breeding grounds, but wintered several hundred km apart (approximately 400 km between birds 757 and 766 and nearly 900 km between 766 and 125). This is the first study of migratory connectivity in European Nightjar and there is clearly much more to discover about links between breeding and wintering areas. An appropriate follow-on to this project would be to track Nightjars from multiple breeding colonies within UK and mainland Europe to determine to what extent widely spaced breeding populations mix in the wintering ground and where first-year birds over-winter in relation to the areas used by their parents. Geolocators have already been used successfully for a similar study in the marine environment (González-Solís et al. Citation2007) and have great potential in this kind of study on terrestrial birds.
The ecoregions that the wintering ranges of birds 757 and 766 fall within all display a mix of scrub, woodland and grassland (Burgess et al. Citation2004), characterized as a forest savanna mosaic. Although the bias in locations caused by aberrations in the light curves at dawn and dusk clearly has some impact on the precision and accuracy of the wintering range, both estimates, from raw and transformed data, fall within these similar ecoregions. While the Albertine Rift montane forest ecoregion that bird 125 wintered within is predominantly montane rainforest, the wider area also includes ericaceous heathland and other moorland ecosystems (Burgess et al. Citation2004, Plumptre et al. Citation2007). The identification of wintering areas for these three Nightjars within the DRC provides evidence that current distribution maps for the African continent (Cleere & Nurney Citation1998, Holyoak Citation2001) under-represent the true range of the species. Given that the ecoregions within which the two 2008 birds wintered extend considerably further west across the DRC and that ecoregions with a similar mixture of scrub, woodland and grassland occur in the neighbouring countries (Olson et al. Citation2001, Burgess et al. Citation2004), this under-representation may cover a significant area.
The tracking of a Nightjar for a whole year is a significant step in understanding the migration, revealing that its pre- and post-nuptial migrations followed different routes. Further study of larger numbers of birds from other breeding localities is needed to show whether this is a common pattern and to identify possible stop-over sites.
The tag used in 2010 showed a higher level of error in its location estimates than those used in 2008. This appears to be caused by an increase in the level of noise in the light-intensity data, probably due to its shorter light stalk. There is potential to improve the signal-to-noise ratio using signal-processing algorithms as investigated in Welch & Eveson Citation(1999), but this was not attempted. Despite the additional estimated error seen in individual locations due to poor fitting of the template, remarkably, the median longitude across the breeding period differed by less than 0.2o from the true position (around 10 km error). Even the latitude error was only 2o (about 220 km). The additional noise is likely to be a reason for no obvious aberrations in the light-intensity curve for tag 125 being seen at dawn and dusk, though the lower sampling rate of light levels may also affect this.
The identification of aberrations within the light curve at dawn and dusk provides a benefit in the potential to collect additional behavioural information from light-intensity data. However, this is balanced by the fact that behaviour may also impact on the accuracy of locations. The impact of behaviour on the accuracy of light-geolocation has already been identified in the effect that rapid directional movement can have by artificially increasing or decreasing apparent day length (Welch & Eveson Citation1999, Shaffer et al. Citation2005) and also with the effect of light attenuation due to diving behaviour in marine species (Musyl et al. Citation2003, Nielsen et al. 2006). The transformation carried out here to assess the impact of the roosting behaviour is somewhat crude, but does show that by understanding the behavioural patterns of study species and taking this into account, it may be possible to improve the accuracy of geolocation estimates. Additionally, if behaviour can be shown to occur systematically at a particular point during dawn and dusk, it may be possible to use this knowledge to assess the accuracy of the timing of dawn and dusk as calculated by the template-fit process. A similar idea has already been used for Macaroni Penguins (Eudyptes chrysolophus), which show an increase in diving depth at dawn and dusk that has been used to measure the timing of sunrise and sunset (Green et al. Citation2009).
ACKNOWLEDGEMENTS
The geolocators used in this study were built with help from Lotek and Biotrack engineers: especially Jude Barkway, Phil Ekstrom, Simon Hardy, Steve Hayward, Djordje Lekovic, Andrew Mahon and Aravind Warrier. Stour Ringing Group members helped to catch Nightjars for deploying and retrieving geolocators: especially Ian Alexander, Jason Fathers, Bob Gifford, Michael Gould and Trevor Squire. Sean Walls and Sarah Levett helped with data analysis, fieldwork and other advice, and the manuscript was improved by two anonymous reviewers. The Forestry Commission kindly allowed access to the land where the study was based.
REFERENCES
- Alexander , I. H. and Cresswell , B. H. 1990 . Foraging by Nightjars Caprimulgus europaeus away from their nesting areas . Ibis , 132 : 568 – 574 .
- Bächler , E. , Hahn , S. , Schaub , M. , Arlettaz , R. , Jenni , L. , Fox. , Afanasyev , V. and Liechti , F. 2010 . Year-round tracking of small trans-Saharan migrants using light-level geolocators . PloS ONE , 5 : e9566
- Burgess , N. , D'Amico Hales , J. , Underwood , E. and Dinerstein , E. 2004 . Terrestrial Ecoregions of Africa and Madagascar: A Conservation Assessment , Washington , DC : Island Press .
- Cleere , N. and Nurney , D. 1998 . A Guide to the Nightjars and Related Nightbirds , Robertsbridge , , UK : Pica Press .
- Cresswell , B. H. 1996 . Nightjars – some aspects of their behaviour and conservation . Br. Wildl. , 7 : 297 – 304 .
- Cresswell , B. H. and Alexander , I. H. 1990 . A case of mate-switching between broods in the Nightjar . Ring. Migr. , 11 : 73 – 75 .
- Cresswell , B. H. and Alexander , I. H. 1992 . “ Activity patterns of foraging Nightjars Caprimulgus europaeus ” . In Wildlife Telemetry , Edited by: Priede , I. G. and Swift , S. M. 642 – 647 . Chichester : Ellis Horwood Ltd .
- Croxall , J. P. , Silk , J. R.D. , Phillips , R. A. , Afanasyev , V. and Briggs , D. R. 2005 . Global circumnavigations: tracking year-round ranges of nonbreeding Albatrosses . Science , 307 : 249 – 250 .
- Ekstrom , P. A. 2002 . “ Blue twilight in a simple atmosphere ” . In Proceedings of SPIE Annual Meeting 2002: Atmospheric Radiation Measurements and Applications in Climate , Edited by: Shaw , J. A. Vol. 4815: 73 – 81 . Bellingham , WA : SPIE .
- Ekstrom , P. A. 2004 . “ An advance in geolocation by light ” . In Proceedings of the International Symposium on Bio-logging Science. Memoirs of the National Institute of Polar Research Special Issue , Edited by: Yasuhiko , N. Vol. 58: 210 – 226 . Tokyo , , Japan : National Institute of Polar Research .
- Ekstrom , P. A. 2007 . Error measures for template-fit geolocation based on light . Deep Sea Res. Pt II , 54 : 392 – 403 .
- Faaborg , J. , Holmes , R. T. , Anders , A. D. , Bildstein , K. L. , Dugger , K. M. , Gauthreaux , S. A. Jr , Heglund , P. , Hobson , K. A. , Jahn , A. E. , Johnson , D. H. , Latta , S. C. , Levey , D. J. , Marra , P. P. , Merkord , C. L. , Nol , E. , Rothstein , S. I. , Sherry , T. W. , Sillett , T. S. , Thompson , F. R. III and Warnock , N. 2010 . Recent advances in understanding migration systems of New World land birds . Ecol. Monogr. , 80 : 3 – 48 .
- Fiedler , W. 2009 . New technologies for monitoring bird migration and behaviour . Ring. Migr. , 24 : 175 – 179 .
- Fuller , W. J. , Broderick , A. C. , Phillips , R. A. , Silk , J. R.D. and Godley , B. J. 2008 . Utility of geolocating light loggers for indicating at-sea movements in sea turtles . Endang. Species Res. , 4 : 139 – 146 .
- González-Solís , J. , Croxall , J. P. , Oro , D. and Ruiz , X. 2007 . Trans-equatorial migration and mixing in the wintering areas of a pelagic seabird . Front. Ecol. Environ. , 5 : 297 – 301 .
- Green , J. A. , Wilson , R. P. , Boyd , I. L. , Woakes , A. J. , Green , C. J. and Butler , P. J. 2009 . Tracking macaroni penguins during long foraging trips using ‘behavioural geolocation’ . Polar Biol. , 32 : 645 – 653 .
- Guilford , T. , Meade , J. , Willis , J. , Phillips , R. A. , Boyle , D. , Roberts , S. , Collet , M. , Freeman , R. and Perrins , C. M. 2009 . Migration and stopover in a small pelagic seabird, the Manx Shearwater Puffinus puffinus: insights from machine learning . Proc. R. Soc. B , 276 : 1215 – 1223 .
- Harris , M. P. , Daunt , F. , Newell , M. , Phillips , R. A. and Wanless , S. 2009 . Wintering areas of adult Atlantic Puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology . Mar. Biol. , 157 : 827 – 836 .
- Hill , R. D. 1994 . “ Theory of geolocation by light levels ” . In Elephant Seals: Population Ecology, Behaviour and Physiology , Edited by: Le Boeuf , B. J. and Laws , R. M. 227 – 235 . Berkeley , CA : University of California Press .
- Holyoak , D. T. 2001 . Nightjars and their Allies , Oxford : Oxford University Press .
- Kenward , R. E. 2001 . A Manual for Wildlife Radiotagging , 2 , London : Academic Press .
- Kenward , R. E. , Pfeffer , R. H. , Al-Bowardi , M. A. , Fox , N. C. , Riddle , K. E. , Bragin , E. A. , Levin , A. , Walls, and Hodder , K. H. 2001 . Setting harness sizes and other marking techniques for a falcon with strong sexual dimorphism . J. Field Ornithol. , 72 : 244 – 257 .
- Kenward , R. E. , Walls , S. S. , South , A. B. and Casey , N. M. 2008 . Ranges 7: For the Analysis and Tracking of location Data. Online Manual , Wareham , , UK : Anatrack Ltd .
- Kirby , K. 2011 . A public servant speaks …. on commuting . The Bulletin , 42 ( 4 ) : 54 – 55 .
- Luschi , P. 2003 . “ Migration and sea turtles ” . In Animal Behaviour and Wildlife C onservation , Edited by: Festa-Bianchet , M. and Apollonio , M. 49 – 62 . Washington , DC : Island Press .
- Musyl , M. K. , Brill , R. W. , Curran , D. S. , Gunn , J. S. , Hartog , J. R. , Hill , R. D. , Welch , D. W. , Eveson , J. P. , Boggs , C. H. and Brainard , R. E. 2001 . “ Ability of archival tags to provide estimates of geographical position based on light intensity ” . In Electronic Tagging and Tracking in Marine fisheries , Edited by: Sibert , J. R. and Nielson , J. L. 343 – 347 . Dordrecht : Kluwer .
- Musyl , M. K. , Brill , R. W. , Boggs , C. H. , Curran , D. S. , Kazama , T. K. and Seki , M. P. 2003 . Vertical movement of Bigeye Tuna (Thunnus obesus) associated with islands, buoys, and seamounts near the main Hawaiian Islands from archival tagging data . Fish. Oceanogr. , 12 : 152 – 169 .
- Nielson , A. , Bigelow , K. A. , Musyl , M. K. and Sibert , J. R. 2006 . Improving light-based geolocation by including sea-surface temperature . Fish. Oceanogr. , 15 : 314 – 325 .
- Olson , D. , Dinerstein , E. , Wikramanayake , E. D. , Burgess , N. D. , Powell. , G. V.N. , Underwood , E. C. , D'Amico , J. A. , Itoua , I. , Strand , H. E. , Morrison , J. C. , Loucks , C. J. , Allnutt , T. F. , Ricketts , T H. , Kura , Y. , Lamoreux , J. F. , Wettengel , W. W. , Hedao , P. and Kassem , K. R. 2001 . Terrestrial ecoregions of the world: a new map of life on earth . Bioscience , 51 : 933 – 938 .
- Perrins , C. M. and Crick , H. Q.P. 1996 . Influence of lunar cycle on laying dates of European Nightjars (Caprimulgus europaeus) . Auk , 113 : 705 – 708 .
- Plumptre , A. J. , Davenport , T. R.B. , Behangana , M. , Kityo , R. , Eilu , G. , Ssegawa , P. , Corneille , E. , Meirte , D. , Kahindo , C. , Herremans , M. , Peterhans , J. K. , Pilgrim , J. D. , Wilson , M. , Languy , M. and Moyer , D. 2007 . The biodiversity of the Albertine Rift . Biol. Conserv. , 134 : 178 – 194 .
- R Development Core Team . 2010 . R: A Language and Environment for Statistical Computing , Vienna , , Austria : R Foundation for Statistical Computing .
- Robinson , W. D. , Bowlin , M. S. , Bisson , I. , Shamoun-Baranes , J. , Thorup , K. , Diehl , R. H. , Kunz , T. H. , Mabey , S. and Winkler , D. W. 2010 . Integrating concepts and technologies to advance the study of bird migration . Front. Ecol. Environ. , 8 : 354 – 361 .
- Rodríguez , A. , Negro , J. J. , Bustamante , J. , Fox , J. W. and Afanasyev , V. 2009 . Geolocators map the wintering grounds of threatened Lesser Kestrels in Africa . Divers. Distrib. , 15 : 1010 – 1016 .
- Shaffer , S. A. , Tremblay , Y. , Awkerman , J. A. , Henrey , R. W. , Teo , S. L.H. , Anderson , D. J. , Croll , D. A. , Block , B. A. and Costa , D. P. 2005 . Comparison of light- and SST-based geolocation with satellite telemetry in free-ranging albatrosses . Mar. Biol. , 147 : 833 – 843 .
- Shaffer , S. A. , Tremblay , Y. , Weimerskirch , H. , Scott , D. , Thompson , D. R. , Sagar , P. M. , Moller , H. , Taylor , G. A. , Foley , D. G. , Block , B. A. and Costa , D. P. 2006 . Migratory Shearwaters integrate oceanic resources across the Pacific Ocean in and endless summer . Proc. Natl Acad. Sci. USA , 103 : 12799 – 12802 .
- Silvano , R. , MacCord , P. , Lima , R. and Begossi , A. 2006 . When does this fish spawn? Fishermen's local knowledge of migration and reproduction of Brazilian coastal fishes . Environ. Biol. Fish. , 76 : 371 – 386 .
- Stutchbury , B. J.M. , Tarof , S. A. , Done , T. , Gow , E. , Kramer , P. M. , Tautin , J. , Fox , J. W. and Afanasyev , V. 2009 . Tracking long-distance songbird migration by using geolocators . Science , 323 : 896
- Teo , S. L.H. , Boustany , A. , Blackwell , S. , Walli , A. , Weng , K. C. and Block , B. A. 2004 . Validation of geolocation estimates based on light level and sea surface temperature from electronic tags . Mar. Ecol. Progr. Ser. , 283 : 81 – 98 .
- Welch , D. W. and Eveson , J. P. 1999 . An assessment of light-based geo-position estimates from archival tags . Can. J. Fish. Aquat. Sci. , 56 : 1317 – 1327 .
- Wernham , C. , Siriwardena , G. M. , Toms , M. , Marchant , J. , Clark , J. A. and Baillie , S. 2002 . The Migration Atlas: Movements of the Birds of Britain and Ireland , Edited by: Wernham , C. , Siriwardena , G. M. , Toms , M. , Marchant , J. , Clark , J. A. and Baillie , S. London : Christopher Helm .
- Wilson , R. P. , Ducamp , J. J. , Rees , G. , Culik , B. M. and Niekamp , K. 1992 . “ Estimation of location: global coverage using light intensity ” . In Wildlife Telemetry: Remote Monitoring and Tracking of Animals , Edited by: Priede , I. M. and Swift , S. M. 131 – 134 . Chichester , , UK : Ellis Horwood .