Abstract
Capsule The value of farmland for birds at a site in Nigeria, West Africa, was highly dependent upon the habitat structure, with wider and more continuous hedges supporting more bird species.
Aim To understand the value of hedges in Nigerian farmland to bird species richness.
Methods We used line transects to survey birds and vegetation on 166 systematically selected hedgerows on fallow, cultivated and harvested fields in Jos East, Nigeria.
Results Wider hedges supported higher species richness. Plant species composition was a weak predictor of bird species richness compared to hedge width. There was no observed difference in distribution of bird richness across field types, suggesting that the value of hedgerows for biodiversity is comparable, regardless of farming method. Continuous hedgerows (hedgerows with vegetation gap < 1.5 m) held higher species richness compared with discontinuous (hedgerows with vegetation gap > 1.5 m) hedges.
Conclusion This result suggests that hedge size and structure, rather than plant species composition, is an important predictor of avian species richness in hedgerows surrounding farmland. Therefore, the value of farmland for birds in this part of Africa is highly dependent upon the habitat structure, with wider and more continuous hedges supporting more bird species.
Tropical deforestation represents the greatest single threat to global biodiversity because tropical forest may support about 70% of the planet's plant and animal species (e.g. Mayaux et al. Citation2005, Chomitz Citation2007). Although the rate of deforestation has slowed down since the 1990s, Africa loses 0.6% of its forest annually, with the greatest loss occurring in heavily forested countries (Kelatwang & Garzuglia Citation2006, UNEP Citation2008). Throughout sub-Saharan West Africa, habitat loss has been rapid through human clearance for fuel wood, grazing and conversion to intensive agriculture as a consequence of population growth (e.g. Grimmett Citation1987, Chomitz Citation2007, Wilson et al. 2007, Norris et al. Citation2010, Riggio et al. Citation2012). These practices result in a reduction in the natural habitat available for biodiversity and also a negative impact on biodiversity due to the effect of habitat fragmentation (Dunn Citation2004, Manu et al. Citation2007, Norris et al. Citation2010). Much effort has gone into designating protected areas in Africa with the hope of saving these areas of crucial habitat (UNEP Citation2008). As a result, conservation effort has been directed to protected areas with little or no attention given to the potential of the replacement anthropogenic habitats such as farmland to conserve species.
Loss of biodiversity in agricultural environments is considered a high conservation priority in Europe (e.g. Tucker & Evans Citation1997, Bradbury et al. Citation2000, Butler et al. 2009, Wilson et al. Citation2009, Wretenberg 2010) and the previous UK government set a public service agreement target to reverse the decline in farmland bird populations by 2020 (Robinson & Sutherland Citation2002, Butler et al. 2007a,b), although this has since been abolished. The USA has also developed a programme that encourages farming in a more holistic, environmentally sound manner in order to enhance biodiversity and provide habitat for wildlife (Sullivan et al. Citation2004, Stamps et al. 2008). However, the value of farmland habitats to bird species is rarely assessed in Africa (but see Wilson et al. Citation1997, Mangnall & Crowe Citation2003, Bolwig et al. Citation2006, Mulwa et al. Citation2012), despite much native habitat being destroyed to make way for farmland, especially in Nigeria. With this pressure on land in Africa, particularly in the light of a growing human population and many people living below the poverty line, optimizing replacement farmland habitat to promote productivity while still maintaining biodiversity is a key problem.
Natural hedgerows usually arise as a result of clearing natural continuous habitats for agricultural encroachment, industrial development and unsustainable logging practices. Hedges have been shown to be important to animal and plant species diversity in Europe (Parish et al. Citation1994, Sparks et al. Citation1996, Hinsley & Bellamy Citation2000, Wehling & Diekmann Citation2009, Pereira & Rodriquez Citation2010, Vickery & Arlettaz Citation2012). In lowland-farming landscapes in Britain, hedgerows comprise one of the most important surviving elements of semi-natural habitat for birds (Hinsley & Bellamy Citation2000). Agricultural field boundaries such as hedgerows in Europe, Asia and North America have beneficial impacts on adjoining cultivated land; however, many farmlands are cleared of such habitats to increase space for cultivation. Similarly, natural hedges are destroyed because many farmers view hedges as a threat to their crops since they attract birds (Jobin et al. Citation2001). In Great Britain, increased use of machinery, and resulting demand for large fields, resulted in removal of 50% of hedgerow stock (Robinson & Sutherland Citation2002).
One major variable that accounts for bird diversity in any habitat is vegetation structure, which is further enhanced by the plant species composition (Wilson et al. Citation1997, Hinsley & Bellamy Citation2000, Jobin et al. Citation2001, Mulwa et al. Citation2012). Effects of habitat fragmentation, patch size and connectivity on species diversity, abundance and distribution have also been considered in many studies (Saunders et al. Citation1991; Wiens Citation1997; Manu et al. Citation2007).
The aim of this study was to investigate the effects of hedgerow structure and plant species composition on bird species richness in farmlands in Jos-East, Nigeria. The objectives were to determine the effect of hedge size, hedge continuity, and field condition, number of trees in the hedge, inter-hedge distance and plant composition on bird species richness. As a starting point and given the relative unstudied nature of farmed landscape and impact of agricultural practices (e.g. cattle grazing, hedgerow removal or thinning) on bird populations in farmland habitats in this region, this paper describes the species composition and bird community structure of hedgerows in our study region and evaluates the effects of hedgerow characteristics on overall bird species richness.
METHODS
Surveys were carried out on agricultural fields with hedgerows 15 km east of Jos, north-central Nigeria (N9°52ʹ E8°58ʹ). The Jos landscape comprises of high plains with scattered rock outcrops ranging from 1220 to 1450 m asl and a number of granite hill ranges that rise to 1781 m. The vegetation is scattered bush and grass, grazed by cattle, sheep and goats, with riparian forest fragments and extensive cultivation (Hulme Citation2007). Commonly cultivated crops include Guinea Corn Sorghum bicolor, Millet Pennisetum glaucum, Acha Digitaria exilis, Maize Zea mays, Rice Oryza sativa, Irish potato Solanum tuberosum, Mango Mangifera indica, Cucumber Cucumis sativus and Lettuce Lactuca sativa. Mixed cropping is the general farming practice in this area where some fields have trees and/or hedgerows. Other fields are open without trees or hedgerows, while some plots are left fallow some years. On average, intensive rain-fed farming begins late April to early May and lasts until early September. Few fields are cultivated during the dry season (late September to early April). Bird and vegetation data were collected from hedgerows in three field types: fallow (n = 72), cultivated (n = 21) and harvested fields (n = 73). A hedge was considered to be on a given field type if fields on both sides of the hedge were in the same condition, i.e. both sides of the hedge were cultivated. One hundred and sixty-six hedgerows each measuring 100 m (hereafter referred to as ‘hedgerow plot’) were systematically selected (Watt Citation1997), with hedgerow plots spaced at intervals of at least 100 m to ensure that bird counts were reasonably independent. No two plots were sampled from the same field. The entire area covered was approximately 100 km2.
Bird data
Bird surveys were carried out from May to June 2008 at the beginning of the wet season. Hedgerows were surveyed between 0600 and 1000 hours each day. Line transects (Bibby et al. Citation2000) were used to census birds. Bird counts were all completed by one observer (HU). During each visit, a transect was walked along the length of the hedgerow plot and the total number of birds detected by sight or sound in the hedgerow or within 15 m on the adjacent field was recorded. Each start and end point of each hedgerow plot was marked with a GPS (GPS Garmin eTrex®, version 3.10).
Hedge data
Each hedgerow plot was divided into four 25-m sections so as to obtain estimates of key features of the habitat. At each section, hedgerow height and width (horizontal spread of vegetation) were measured and recorded. Similarly foliage density index was determined using a 50 × 50 cm black and white chequered board with 10-cm subdivisions, recording the distance at which 50% of board was obscured by vegetation (Bibby et al. Citation2000). Within each section, a 2 × 2 m quadrat was placed centrally and ground cover (taken as amount of dry litter on the ground) and bare ground were estimated by eye to the nearest 5% (Bullock Citation2006). Plant species composition was determined by recording presence/absence and identified using Arbonnier Citation(2004). The number of trees in each hedgerow plot (trees with circumference at breast height over 30 cm) was recorded. Hedgerow continuity was determined by observing whether a hedgerow plot was continuous (hedgerows with vegetation gap = 0 or < 1.5 m) or discontinuous (hedgerows with vegetation gap > 1.5 m). Distance of the nearest hedge to a focal hedgerow was estimated by eye to the nearest 5 m. The number of shrubs (bushes) and trees within each adjacent field, and the field status (cultivated, harvested and fallow) were recorded for each field.
Data analysis
R version 2.15.0 was used for statistical analyses. Means are given with ± one standard error.
For each 100 m hedge, bird species richness was calculated as the total number of species recorded. We summarized bird species according to guilds by considering the major food resource consumed (based on species food habits described in Fry et al. 1988, Borrow & Demey 2010). This resulted in six functional groups. The total number of species in each feeding guild was summed per hedge. The presence/absence of each species and guild for each hedgerow plot was determined and proportion frequency of occurrence over 166 hedgerow plots was calculated ().
Table 1. Frequency of occurrence of bird species in hedges. Frequency is the number of occurrences of a species or guild over 166 hedgerow plots; proportion is the proportion of hedgerow plots where the species or guild was recorded.
Shrub density for each 100 m hedgerow was calculated using MacArthur foliage density index, K:
K = ln2/D
where D is the distance from the observer to the chequered board (McArthur & McArthur Citation1961, Bibby et al. Citation2000).
To control for non-orthogonality in our model, Spearman's correlation was used to determine significant associations between habitat variables: hedge height, groundcover and foliage density index showed strong and significant correlation with hedge width (coefficient of correlation, r s > 0.50, P < 0.05). Consequently we omitted these three predictor variables from our model while retaining hedge width (we considered hedge width to be more biologically important). We fitted Generalized Linear Models to the data using Poisson error and log link, with bird species richness as the response variable; explanatory variables incorporated in the model were hedge width, hedge continuity, number of trees on hedge, number of plant species in the hedge, distance to nearest hedge and field condition. Models for all possible parameter subsets were compared in terms of parsimony and prediction through the calculation of AIC (Burnham & Anderson 2002). These analyses were performed with the MuMIn package in R which returns all possible models in order of importance and plausibility with the lower AIC values representing better models. For the purpose of effect size, we standardized covariates (i.e. converted as mean = 0, sd = 1).
RESULTS
As a group, seed- and insect-eating birds occurred in at least 90% of surveyed hedges, with higher densities in wider and continuous hedges; frugivores and nectarivores occurred in more than 50% of surveyed hedgerow plots; carnivores (raptors) occurred in 2% of surveyed hedges (). Ninety-five species belonging to 39 families were recorded from 166 hedgerow plots, with Estrildidae, Sylviididae, Ploceidae, Columbididae, Pycnonotidae and Nectariniidae occurring in at least 50% of surveyed hedges. As species, Common Bulbul Pycnonotus barbatus, Red-cheeked Cordon-Bleu Uraeginthus angolensis and Scarlet-chested Sunbird Chalcomitra senegalensis were recorded in more than half of surveyed hedges; Variable Sunbird Cinnyris venustus, African Thrush Turdus pelios, Grey-backed Camaroptera Camaroptera brachyura and Tawny-flanked Prinia Prinia subflava were recorded in at least 30% of hedges (). Fifty-six plant species were identified in the hedgerow plots.
Bird–habitat relations
The model selection procedure showed that there were five top models (). Discontinuous hedges (negative effect) and hedge width (standardized coefficient, sc = 0.111) were included in all five best models (). Distance to nearest hedge (sc = 0.057) and number of trees in the hedge (sc = 0.045) were included in four and three of the five best models, respectively, and both had positive effects on species richness. The mean (± se) distance between hedges was 63.1 (± 2.8) m (n = 152). Within farms the mean number of shrub and trees were 4.35 ± 0.36 (n = 165) and 3.13 ± 0.26 (n = 165), respectively.
Table 2. Top Generalized Linear Models (GLMs) of variables predicting bird species richness on hedgerows on West African farmlands. Estimates of variables not included in the models are indicated by blank spaces. ∆ i is difference in AIC values of each model relative to the best model. Cultivated field and continuous hedge are reference categories. The table reports all models with ∆ i < 5. *significant predictors in GLM.
Figure 1. Relationship between bird species richness and hedge width for different hedge structures. The predicted lines are based on the best model: solid line (continuous hedge) and dashed line (discontinuous hedge).
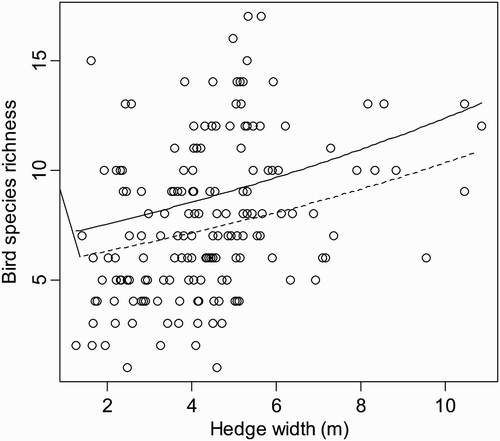
Although hedgerow plots adjacent to cultivated fields had the highest (mean ± se) bird species richness (1.91 ± 0.09) compared with fallow (1.86 ± 0.04) and harvested fields (1.69 ± 0.06), these differences were not important in predicting bird richness.
DISCUSSION
The results presented here show that hedgerows in West Africa seem to play similar role to those in Europe. Bird species richness showed a positive relationship with hedgerow width. Parish et al. Citation(1994) also found a similar relationship with hedge width, indicating though that taller hedges were more important for bird species richness. Similarly, larger hedgerows (greater width) contain more species, presumably because they meet the varied requirements of foraging, roosting, nesting and protection or refuge from predators (Sparks et al. Citation1996, Hinsley & Bellamy Citation2000, Whittingham & Evans Citation2004). In Britain, Hinsley et al. Citation(1999) found that bird species richness and abundance showed a better relationship with hedge width than with hedge height; a narrow hedge will be generally unfavourable to most birds probably due to a combination of lack of resources and exposure to weather and predators.
Plant species composition was a weaker predictor of bird species richness in hedgerows compared to hedge width. This result contradicts the prediction that higher plant species diversity within hedges should result in greater potential food resources and hence more birds (Hinsley & Bellamy Citation2000). It suggests, therefore, that habitat structure may be more important and probably indicates that hedges on fields serve more to provide cover or protection for birds (Hinsley & Bellamy Citation2000) and nests (Sparks et al. Citation1996, Perkins et al. Citation2002) and also as connection between habitats (Fahrig & Merriam Citation1994). The botanic composition of a hedge may become less important if the width of the hedge is narrow. Besides, most of the species recorded here were not specialists. However, we note that in this study, the use of 2 × 2 m quadrats in four equidistant sections to sample plant species in a length of 100 m may mean that some species may have been missed. Similarly, the effect of the presence of individual species – for example, plants with a high production of fruit – might be greater than the number of species per se, so plant composition cannot be ruled out entirely as an important factor.
Bird species richness was not affected negatively by distance to the nearest hedge, possibly because the maximum hedge isolation distance was insufficient in this study to act as much of a constraint. Similarly, most of the farms contained trees or bushes that probably served as connections between the hedges, thereby further precluding any negative effect of hedge isolation (Fahrig & Merriam Citation1994).
Bird species richness increased with number of trees in the hedge. This result is consistent with other studies (e.g. Parish et al. Citation1994, Hinsley et al. 2000); increasing number of trees in hedges probably indicates an increase in niche opportunities and habitat types which affects bird abundance positively. However, it should be noted that the UK studies referred to involve much smaller total numbers of species (20–30), with the same core species likely to be found in all survey plots compared with an average richness of 8 species per plot with a total pool of 95 for this study. This implies that there must be many replacement species within guilds playing the same roles in different plots. In a review of value of hedgerows to birds in lowland farming landscape in Britain, Hinsley et al. (2000) discussed that the presence of well-grown, dead or decaying trees is beneficial to many species, providing nest-holes, foraging sites and perches. Tall hedges with many trees are probably attractive to many woodland bird species because of the resemblance of this type of structure to woodland edge. Wilson et al. Citation(2007) reported that number of bird species, total number of birds and the diversity of birds decreased significantly with a decrease in tree density, whereas there was no similar decline on adjacent farmland habitat that had already been deforested.
Field condition, i.e. whether a field was cultivated, harvested or fallow, did not significantly influence bird species richness, suggesting that the value of hedgerows for biodiversity is comparable, regardless of farming type employed. However, hedgerows adjacent to cultivated and fallow fields had a higher number of bird species compared to harvested fields. This might reflect higher food availability and a complex habitat structure around fallow fields. Fallow fields would also be favoured especially by granivorous species, in this case weavers, that seek grains and weed seed (Bradbury et al. Citation2000) as well as insects because of no artificial chemical input and low disturbance. Chamberlain et al. Citation(1999), in a study of the effects of habitat type and management on the abundance of breeding skylarks in Britain, showed that set aside (equivalent to fallow) was consistently associated with high densities of birds. The cultivated habitat had a semblance of continuous vegetation which graded into the hedge itself (see Krebs et al. Citation1991) to increase its spread and created habitat for more species and may have provided some level of cover from predators.
Continuity in hedgerow vegetation was important because continuous hedges held higher bird species compared with discontinuous hedges. This pattern is probably an indication that a continuous matrix provides more space, reduces predation risk and creates corridors for movement along the hedge to adjoining suitable habitats. Ninety-five percent of the bird species encountered in this study were edge species (cf. Manu Citation2002, Borrow & Demey Citation2004) and they respond differently to differences in hedge continuity or fragmentation, suggesting that detrimental edge effects have a disproportionately greater influence in small, discontinuous habitat patches (Tattersall et al. 2002). A discontinuous hedge will function less as a corridor in terms of aiding birds moving between habitats. Jobin et al. Citation(2001) also found a negative association between the proportion of gaps in margins and bird communities. It has been suggested generally that farm vegetation structure and predation risk may influence the demography of farmland bird populations (Whittingham & Evans Citation2004).
CONCLUSIONS
Our results suggest that hedge size rather than plant species composition is an important predictor of avian species richness in hedgerows surrounding farmlands. Similarly, discontinuity in hedge vegetation greatly affects bird species richness. This suggests that, as in Europe, the value of farmland for birds in African farmland is highly dependent upon the habitat structure (Sparks et al. Citation1996, Robinson & Sutherland Citation1999, Hinsley & Bellamy Citation2000, Whittingham & Evans Citation2004, Wretenberg et al. Citation2010, Mulwa et al. Citation2012), with wider and more continuous hedges supporting more bird species. Therefore, sustainable conservation efforts should be aimed at promoting fields with more edge habitat, so that farmland can play a dual role in supporting biodiversity and man, particularly if the conservation goal is to maintain high local species richness (see also Mulwa et al. Citation2012).
However, as Africa expands and intensifies its agriculture (Wilson et al. 2007, Norris et al. Citation2010, Deininger & Byerlee 2011), with local farmers increasingly having access to bulldozers to clear and tractors to prepare fields, hedgerow removal, as well as shrubs and small bushes within fields, is likely to increase. Such removal is one of the drivers of biodiversity decline in European farming systems (Robinson & Sutherland Citation2002, Newton Citation2004, Donald et al. 2001, Whittingham et al. Citation2006, Butler et al. 2007a,b, Wilson et al. Citation2009, Norris et al. Citation2010) and so Africa may experience a similar trend in loss of biodiversity, but little of this has been documented to date. This is one of the few studies in Africa, particularly in West Africa, that has investigated aspects of agricultural practices that are perceived to cause declines in biodiversity. It is therefore important to expand research in understanding the impact of man-managed habitat to biodiversity in this region because it appears that agricultural landscape will continue to increase, especially in developing countries as world food demand is expected to more than double its present rate in the next 40 years (Tilman et al. Citation2002, Green et al. Citation2005, Godfray et al. Citation2010).
ACKNOWLEDGEMENTS
This work was carried out during an MSc to Hope Usieta funded by the A.P. Leventis Conservation Foundation. Afan Ajan provided assistance in the field. We thank Katherine Jones and Tatsuya Amano who advised on statistical analysis and William J. Sutherland for helpful comments and suggestions to earlier versions of this paper. Gavin Siriwardena and a second anonymous reviewer provided useful comments and suggestions. This is publication 60 from the AP Leventis ornithological Research Institute.
REFERENCES
- Arbonnier , M. 2004 . Trees, Shrubs and Lianas of West African Dry Zones , Weikersheim, Germany : CIRAD, MARGRAF Publishers .
- Bibby , C. J. , Burgess , N. D. , Hill , D. A. and Mustoe , S. H. 2000 . Bird Census Techniques , 2 , London : Academic Press .
- Bolwig , S. , Pomeroy , D. , Tushabe , H. and Mushabe , D. 2006 . Crops, trees, and birds: Biodiversity change under agricultural intensification in Uganda's farmed landscapes . Dan. J. Geogr. , 106 : 115 – 130 .
- Borrow , N. and Demey , R. 2004 . Birds of Western Africa , London : Christopher Helm .
- Bradbury , R. B. , Kyrkos , A. , Morris , A. J. , Clark , S. C. , Perkins , A. J. and Wilson , J. D. 2000 . Habitat associations and breeding success of yellowhammers on lowland farmland . J. Appl. Ecol. , 37 : 789 – 805 .
- Bullock , M. J. 2006 . “ Plants ” . In Ecological Census Techniques: A Hand Book , 2 , Edited by: Sutherland , W. J. New York : Cambridge University Press .
- Burnham, K.P. & Anderson, D.R. 2002. Model selection and multimodel inference: A practical information - theoretic approach. 2nd ed. Springer, New York.
- Butler , S. , Vickery , J. and Norris , K. 2007a . A risk assessment framework for evaluating progress towards sustainability targets . Aspects Appl. Biol. , 81 : 317 – 323 .
- Butler , S. J. , Vickery , J. A. and Norris , K. 2007b . Farmland biodiversity and the footprint of agriculture . Science , 315 : 381 – 383 .
- Butler, S., Brooks, D., Feber, R.E., Storkey, J., Vickery, J. & Norris, K. 2009. A cross taxonomic index for quantifying the health of farmland biodiversity. J. App. Ecol. 46: 1154–1162.
- Chamberlain , D. E. , Wilson , A. M. , Browne , S. J. and Vickery , J. A. 1999 . Effects of habitat type and management of Skylarks in the breeding season . J. Appl. Ecol. , 36 : 856 – 870 .
- Chomitz , K. M. 2007 . At Loggerheads? Agricultural Expansion, Poverty Reduction and Environment in the Tropical Forests World Bank Policy Research Report, Washington, DC
- Donald, P.F., Green, R.E. & Heath, M.F. 2001. Agricultural intensification and the collapse of Europe's farmland bird populations. Proc. R. Soc. B 268: 25–29.
- Deininger, K. & Byerlee, D. 2011. Rising global interest in farmland: can it yield sustainable and equitable benefits? The World Bank, Washington D.C. DOI: 10.1596/978-0-8213-8591-3.
- Dunn , R. R. 2004 . Managing the tropical landscape: a comparison of effects of logging and forest to agriculture on ants, birds and Lepidoptera . For. Ecol. Manage. , 191 : 215 – 224 .
- Fahrig , L. and Merriam , G. 1994 . Conservation of fragmented populations . Conserv. Biol. , 8 : 50 – 59 .
- Fry, C.H., Keith, S. & Urban, E.K. 1988. The birds of Africa. Vol. 1-6. Academic press, London.
- Godfray , C. J. , Beddington , J. R. , Crute , I. R. , Haddad , L. , Muir , J. F. , Pretty , J. , Robinson , S. , Thomas , S. M. and Toulmin , C. 2010 . Food security: the challenge of feeding 9 billion people . Science , 327 : 812 – 818 .
- Green , R. E. , Cornell , S. J. , Scharlemann , J. P.W. and Balmford , A. 2005 . Farming and the fate of wild nature . Science , 307 : 550 – 555 .
- Grimmet , R. 1987 . A Review of the Problems Affecting Paleartic Migratory Birds in Africa , ICBP Study Report No. 22. International Council for Bird Preservation, Cambridge
- Hinsley , S. A. and Bellamy , P. E. 2000 . The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review . J. Environ. Manage. , 60 : 33 – 49 .
- Hinsley , S. A. , Bellamy , P. E. , Sparks , T. H. and Rothery , P. 1999 . “ A field comparison of habitat characteristics and diversity of birds, butterflies and plants between game and nongame areas ” . In Lowland Game Shooting Study Edited by: Firbank , L. G. 69 – 116 . ITE final report to the British Association for Shooting and Conservation. ITE, Cumbria
- Hulme , M. 2007 . “ Density and diversity of birds on farmland in West Africa ” . PhD Thesis, University of St Andrews, UK
- Jobin , B. , Choinière , L. and Bélanger , L. 2001 . Bird use of three types of field margins in relation to intensive agriculture in Québec, Canada . Agric. Ecosyst. Environ. , 84 : 131 – 143 .
- Kelatwang , S. and Garzuglia , M. 2006 . Changes in forest Area in Africa 1990–2005 . Int. For. Rev. , 8 : 21 – 30 .
- Krebs , J. R. , Wilson , J. D. , Bradbury , R. M. and Siriwardena , G. M. 1999 . The second silent night? . Nature , 400 : 611 – 612 .
- McArthur , R. H. and McArthur , J. W. 1961 . On bird species diversity . Ecology , 42 : 594 – 598 .
- Mangnall , M. J. and Crowe , T. M. 2003 . The effects of agriculture on farmland bird assemblages on the Agulhas Plain, Western Cape, South Africa . Afr. J. Ecol. , 41 : 266 – 276 .
- Manu , S. A. 2002 . Effects of habitat fragmentation on the distribution of forest birds in South Western Nigeria with particular reference to Ibadan Malimbe and other Malimbes . PhD Thesis, University of Oxford, UK
- Manu , S. , Peach , W. and Cresswell , W. 2007 . The effects of edge, fragment size and degree of isolation on avian species richness in highly fragmented forest in West Africa . Ibis , 149 : 287 – 297 .
- Mayaux , P. , Holmgren , P. , Achard , F. , Eva , H. , Stibig , H. and Branthome , A. 2005 . Tropical forest cover change in the 1990's and options for future monitoring . Phil. Trans. R. Soc. B , 360 : 373 – 384 .
- Mulwa , R. K. , Böhning-Gaese , K. and Schleuning , M. 2012 . High bird species diversity in structurally heterogeneous farmland in Western Kenya . Biotropica , 44 : 801 – 809 .
- Newton , I. 2004 . The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions . Ibis , 146 : 579 – 600 .
- Norris , K. , Asase , A. , Collen , B. , Gockowksi , J. , Mason , J. , Phalan , B. and Wade , A. 2010 . Biodiversity in a forest–agriculture mosaic – the changing face of West African rainforests . Biol. Conserv. , 143 : 2341 – 2350 .
- Parish , T. , Lakhani , K. H. and Sparks , T. H. 1994 . Modelling the relationship between bird population variables and hedgerow and other field margin attributes. I. Species richness of winter, summer and breeding birds . J. Appl. Ecol. , 31 : 764 – 775 .
- Pereira , M. and Rodriguez , A. 2010 . Conservation value of linear woody remnants for two forest carnivores in a Mediterranean agricultural landscape . J. Appl. Ecol. , 47 : 611 – 620 .
- Perkins , A. J. , Whittingham , M. J. , Morris , A. J. and Bradbury , B. R. 2002 . Use of field margins by foraging Yellowhammers . Emberiza citronella. Agric. Ecosyst. Environ. , 93 : 413 – 420 .
- R Development Core Team . 2012 . R: A Language and Environment for Statistical Computing , R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org
- Riggio , J. , Jacobson , A. , Dollar , L. , Bauer , H. , Becker , M. , Dickman , A. , Funston , P. , Groom , R. , Henschel , P. , de Iongh , H. , Lichtenfeld , L. and Pimm , S. 2012 . The size of savannah Africa: a lion's (Panthera leo) view . Biodiv. Conserv. , http://dx.doi.org/10.1007/s10531-012-0381-4
- Robinson , R. A. and Sutherland , W. J. 1999 . The winter distribution of seed-eating birds: habitat structure, seed density and seasonal depletion . Ecography , 22 : 447 – 454 .
- Robinson , R. A. and Sutherland , W. J. 2002 . Post-war changes in arable farming and biodiversity in Great Britain . J. Appl. Ecol. , 39 : 157 – 176 .
- Saunders , D. A. , Hobbs , R. J. and Margules , C. R. 1991 . Biological consequence of ecosystem fragmentation: a review . Conserv. Biol. , 5 : 18 – 32 .
- Sparks , T. H. , Parish , T. and Hinsley , S. A. 1996 . Breeding birds in field boundaries in an agricultural landscape . Agric. Ecosyst. Environ. , 60 : 1 – 8 .
- Stamps, W.T., Dailey, T.V., Gruenhagen, N.M. & Linit, M.J. 2008. Soybean yield and resource conservation field border. Agriculture, Ecosystems and Environment B:142–146.
- Sullivan , P. , Hellerstein , D. , Hansen , L. , Johansson , R. , Koenig , S. , Lubowski , R. , McBride , W. , McGranahan , D. , Roberts , M. , Vogel , S. and Bucholtz , S. 2004 . The Conservation Reserve Program: Economic Implications for Rural America AER-834. Economic Research Service, United States Department of Agriculture, Washington, DC
- Tattersal, F.H., MacDonald, D.W., Hart, B.J., Johnson, P., Manley, W. & Feber, R. 2002. Is habitat linearity important for small mammal communities on farmland? J. App. Ecol. 39: 643–652.
- Tilman , D. , Cassman , K. G. , Matson , P. A. , Naylor , R. and Polasky , S. 2002 . Agricultural sustainability and intensive production practices . Nature , 418 : 671 – 677 .
- Tucker , G. M. and Evans , G. I. 1997 . Habitats for Birds in Europe; A Conservation Strategy for the Wider Environment Birdlife Conservation Series No. 6. Birdlife International, Cambridge, UK
- UNEP . 2008 . Africa: Atlas of Our Changing Environment , Division of Early Warning and Assessment (DEWA) United Nations Environment Programme (UNEP)
- Vickery , J. and Arlettaz , R. 2012 . “ The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes ” . In Birds and Habitat: Relationships in Changing Landscapes , Edited by: Fuller , R. J. Cambridge : Cambridge University Press .
- Watt , T. A. 1997 . Introductory Statistics for Biology Students , 2 , London : Chapman and Hall/CRC .
- Wehling , S. and Diekmann , M. 2009 . Importance of hedgerows as habitat corridors for forest plants in agricultural landscapes . Biol. Conserv. , 142 : 2522 – 2530 .
- Whittingham , M. J. and Evans , K. L. 2004 . The effects of habitat structure on predation risk of birds in agricultural landscapes . Ibis , 146 : 210 – 220 .
- Whittingham , M. J. , Devereux , C. L. , Evans , A. D. and Bradbury , R. B. 2006 . Altering perceived predation risk and food availability: management prescriptions to benefit farmland birds on stubble fields . J. Appl. Ecol. , 43 : 640 – 650 .
- Wiens , J. A. 1997 . “ Metapopulation dynamics and landscape ecology ” . In Metapopulation Biology: Ecology, Genetics and Evolution , Edited by: Hanski , I. A. and Gilpin , M. E. 43 – 62 . London : Academic Press .
- Wilson , C. J. , Reid , R. S. , Stanton , N. L. and Perry , B. D. 1997 . Effects of land-use and Tsetse fly control on bird species richness in Southwestern Ethiopia . Conserv. Biol. , 11 : 435 – 447 .
- Wilson , J. D. , Evans , A. D. and Grice , P. V. 2009 . Bird Conservation and Agriculture , Cambridge : Cambridge University Press .
- Wilson , J. M. , Cresswell , W. C. , Vickery , J. , Jones , P. and Holt , S. 2007 . Changes in densities of Sahelian bird species in response to recent habitat degradation . Ostrich , 78 : 247 – 253 .
- Wretenberg , J. , Pärt , T. and Berg , Å. 2010 . Changes in local species richness of farmland birds in relation to land-use changes and landscape structure . Biol. Conserv. , 143 : 375 – 381 .