Abstract
Capsule The provision of supplementary food in early spring led to an advance in laying date and increased productivity for the Great Spotted Woodpecker.
Aims To use the experimental provision of supplementary food in the prebreeding period to investigate the role of food supply in determining laying date, subsequent synchrony with natural food availability and the impact on productivity for the Great Spotted Woodpecker.
Methods Supplementary food in the form of fat blocks was provided at 13 feeding stations distributed over half of a 100-ha study wood from early February until late April 2011 with the other half of the wood left unfed. The breeding parameters (first egg date, clutch size, number of young fledged per nest and nest success) of Great Spotted Woodpeckers were monitored throughout the wood and data from the ‘fed’ and ‘unfed’ zones compared. The same breeding parameters collected throughout the wood in the five preceding years (2006–10) when there was no supplementary feeding were used to provide control data. The temporal pattern in the abundance of the main natural prey was monitored using caterpillar frass traps.
Results In 2011, the mean first egg date for woodpecker nests in the fed zone of the wood was 4–5 days earlier than for those nests in the unfed zone. Nests in the fed zone were almost twice as productive as those in the unfed zone even though the supplementary feeding stopped before the main period of chick rearing. There were no differences in the breeding parameters in the two zones of the wood in the control years when there was no supplementary feeding. 2011 was a very warm spring and the natural prey abundance peaked very early so that none of the woodpeckers were well synchronized with their main breeding season prey. However, the small advancement in first egg date meant the supplemented birds were better synchronized than the controls.
Conclusion The response of Great Spotted Woodpeckers to supplementary feeding suggests they may be limited in their ability to shift their breeding period to maintain synchrony with their natural prey in the breeding season. This has important implications for the response of the birds to warm springs which are expected to be more frequent under future climate change. Use of garden feeders by Great Spotted Woodpeckers has the potential to increase breeding success and may be one of the many factors contributing to their current population increase.
The provision of supplementary food has been used widely as an experimental tool to investigate the role of food in the life history traits of birds (see Newton Citation1998 for a review) and increasingly as a conservation measure for species under threat (Castro et al. Citation2003, Houston et al. Citation2007, Schoech et al. Citation2008). Most early studies looked at the impacts of supplementary feeding, usually outside the breeding season, on the demography of birds (Newton Citation1998). More recently studies have looked at the impact of feeding at various times of year on survival and breeding parameters (Arcese & Smith Citation1988, Källander & Karlsson Citation1993, Soler & Soler Citation1996, Kelly & Van Horne Citation1997, Robb et al. Citation2008, Harrison et al. Citation2010) in order to isolate the role of food in determining these life-history processes.
In this article we describe an experiment to look at the impact of the provision of supplementary food in late winter and early spring on the subsequent breeding performance of the Great Spotted Woodpecker Dendrocopos major. The primary purpose of the experiment was to explore the degree to which the onset of breeding in this species is affected by the food supply early in the season and the impact this might have on breeding productivity. The population of Great Spotted Woodpeckers in Britain is increasing rapidly (Baillie et al. Citation2012) and they are now frequent visitors to garden feeders (Cannon et al. Citation2005). This utilization of garden feeders has been suggested as a possible contributing factor to the population increase (Fuller et al. Citation2005, Smith Citation2005) and this study helps to inform this discussion.
METHODS
The study was carried out in Wormley Wood, Hertfordshire, southern England (51o44ʹN, 0o6ʹW), a mature Oak Quercus petraea/robor and Hornbeam Carpinus betula wood and a long-term study site for Great Spotted Woodpeckers (Smith Citation2005, Citation2006). The wood was selected because of its large extent (100 ha), relatively uniform habitat, good numbers of breeding woodpeckers (around 25 pairs in recent years), lack of history of any supplementary feeding and its relative isolation from houses and gardens where supplementary feeding may have already been taking place. There is only one private house and garden within 200 m of the boundary of the wood and the nearest village is 1 km away. Great Spotted Woodpeckers can easily move over such distances (Smith, pers. comm.) so our aim was to minimize the confounding influence of other local feeding sites rather than eliminating it entirely.
For the purposes of the experiment, the wood was divided approximately in half on a north–south line. In the western section supplementary food in the form of fat blocks (RSPB ‘Super Suet Cakes’, insect and mealworm flavours; 11% protein, 29% fat, 50% carbohydrate) in wire mesh holders was provided at 13 feeding stations set out on a 200 m grid. The 200 m grid was selected on the basis of practicality and with the aim of providing a feeding station within the expected area around each woodpecker nest/territory (4 ha) (Smith Citation2006). The feeding stations were set up in early February 2011 and kept supplied with food until the end of April, thus covering the prebreeding and egg laying period of the birds but not the young rearing phase (Smith Citation2005, Citation2006). The fat blocks were weighed (to give a measure of consumption rate) and replenished every 5 days or so. As the rate of food consumption increased during the course of the experiment, additional blocks were provided at each feeding station to ensure that the food never ran out in the 5-day interval. No attempt was made to exclude other birds such as tits and Nuthatches Sitta europea from the feeders and indeed these species were probably taking the majority of the food. However, throughout the study, Great Spotted Woodpeckers were seen regularly using the feeders. The feeders were hung on long wires to limit their accessibility to Grey Squirrels Sciurus carolinensis. We were reasonably successful in excluding the squirrels, although in one case we needed to hang the feeder from a long wire stretched between trees to keep them off. We excluded the data from two trap/intervals where it was clear that the squirrels had taken a considerable fraction of the food.
In these woodlands, the natural food for woodpeckers and other species in the breeding season is dominated by defoliating caterpillars which have a short peak of abundance in April/May (Smith et al. Citation2011). To establish the timing of this peak we ran four simple frass trays in the wood following methods described in Smith et al. Citation(2011): two trays each in the fed and unfed zones. The tray contents were collected every 5 days, dried, sorted and weighed to the nearest mg with the figures being converted to g m−2 day−1.
Great Spotted Woodpecker nesting abundance and breeding success was monitored using methods established over 25 years (Smith Citation2005, Citation2006). Nests were searched for from early April onwards and once located the status was determined by visual cues (Jackson Citation1976) and the contents inspected every few days using a video camera on a telescopic pole (see Smith & Charman Citation2012 for details). All nesting attempts were followed to completion and checked after fledging for any dead unfledged young. First egg dates were established from observation of incomplete clutches or, in most cases, by back-calculating from known hatching or fledging dates. Clutch size was determined from nests discovered and inspected during incubation and the numbers of fledged young from nests inspected within 3–4 days of the fledging day when young can be counted easily. In Great Spotted Woodpeckers, unhatched eggs are broken or removed from the nest within a few days and small young can be difficult to count on video images, so it was not appropriate to analyse hatching success. Great Spotted Woodpeckers are essentially single-brooded, only laying replacement clutches if they fail during egg laying or early in incubation (Smith Citation2005, Citation2006), so we had no need to consider first and second broods. The birds were nesting in natural sites, usually in cavities freshly excavated that spring, so inevitably we did not discover all nesting attempts at the outset and so our sample sizes vary for the different nesting parameters.
The boundary between the fed and unfed zones of the wood was set at 100 m to the east of the most easterly line of feeders to provide a substantial buffer zone. We could not discount the possibility of birds from the unfed zone having access to feeders, but there was a substantial contrast in the proximity of feeders to nests between the two zones. The median nearest distance to a feeder for nests in the fed zone was 71 m (range 45–122 m) compared with 453 m (range 147–771 m) in the unfed zone.
We used nesting parameters collected in the same way in the same two areas of the wood over the period 2006–2010 as control data when no supplementary food was provided and a Generalized Linear Model (GLM) framework to analyse the impact of supplementary feeding. The proportions of nests in the fed and unfed zones have been analysed using binomial errors with a logit link function with each year as a replicate. First egg date, clutch size and numbers of young fledged per nest have been analysed in GLMs with year, feeding zone and their interactions as categorical variables with the expectation that the effect of feeding in 2011 would be shown by a significant year*feeding zone interaction in that year. For first egg date, normal errors with identity link have been used whereas for clutch size and number of young fledged per nest we have used Poisson errors and log link function. Nest failure rates were analysed within the same basic framework but using the logistic exposure link (Shaffer 2004a). Because the numbers of failures were low, we have combined the data from egg and young stages in these analyses. Long-term studies have shown seasonal declines in both clutch size and the numbers of young fledged per nest for the Great Spotted Woodpecker (Smith Citation2006). To investigate whether such patterns occurred in this study we have calculated the egg difference for each nest (first egg date for the nest – median first egg date for the year) and modelled clutch size and the number of young fledged per nest in a GLM framework with Poisson errors and a log link function with egg difference and year as the predictive variables. Statistical analyses were carried out in Minitab16 (Minitab 2010) and R (R Core Team Citation2012). The logistic exposure link (Shaffer Citation2004a) was implemented in R by Shaffer and Herzog (Shaffer Citation2004b).
RESULTS
Consumption of supplementary food and natural prey abundance
During the three-month period of food supplementation a total of 90 kg of food was consumed. The rate of consumption started low () but increased to a peak in early April (days 90–105), falling rapidly thereafter. The feeders were last replenished on 27 April but were left in place until 11 May, although most were already empty by then.
Figure 1. The mean rate of fat consumption (left axis and open symbols) and frass fall (right axis and closed symbols) versus date. The periods of egg laying and incubation (1) and chick rearing (2) are indicated at the base of the graph for the fed (upper line) and unfed zones (lower line).
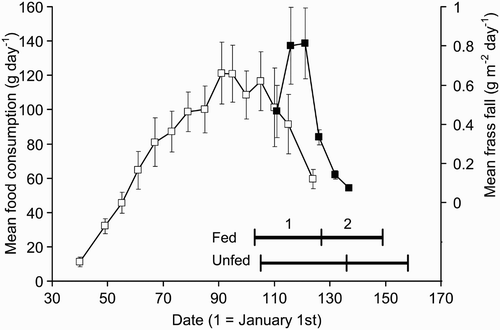
The data from the frass traps showed that the caterpillar abundance increased as the consumption of supplementary food fell, reaching its peak in late April/early May and falling rapidly thereafter (). Although based on a small number of traps, there was no difference in the date of peak frass fall in the two zones (fed zone mean = day 118.8 ± 1.3 se, unfed mean = day 120.0 ± 2.0 se; t = 0.53, P = 0.69).
Distribution of nests
There was no evidence of any change in the distribution of nests between the fed and unfed zones in the year when supplementary feeding was carried out. Between 2006 and 2010, 69 of the 121 nests found were in the fed zone (proportion = 0.57) whereas in 2011, 16 of 27 nests were in that zone (proportion = 0.59) (Binomial GLM of annual proportions: z = 0.21, P = 0.83).
First egg dates
There was considerable annual variation in first egg date but in no year except 2011 was there a significant difference between the two zones of the wood (; GLM, parameter estimate year 2011*feeding zone = 5.7, t = 2.3, P = 0.03, no other year*feeding zone interactions significant). In 2011 eggs were laid in nests in the fed zone around 5 days earlier than in the unfed zone. It is notable that 2011 was a very early nesting year matched only by 2007 in these data.
Table 1. The first egg dates for nests in the fed and unfed zones of Wormley Wood from 2006 to 2011. Supplementary food was provided in the fed zone only in 2011. In a model of all years' data there was a significant difference between the fed and unfed zones in 2011 only (in bold).
Clutch size
Although the sample sizes were small, there were no significant differences between the clutch sizes in the fed and unfed zones in the year of the experiment, 2011 or any of the other years (; Poisson GLM parameter estimate, year 2011*feeding zone = 0.06, z = 0.2, P = 0.87, no other year*feeding zone interactions significant).
Table 2. The clutch size and numbers of young fledged for nests in the fed and unfed zones. Supplementary food was provided in the fed zone in 2011. 2006–2010 were the control years when no supplementary food was provided in either zone. In models including all years' data there was no significant difference in clutch size between the fed and unfed zones in any year including 2011, but there was a significant difference in the number of fledged young between the fed and unfed zones in 2011 only (in bold).
Nest success
The overall nest failure rate was extremely low. In 2011 there were only two nest failures from the 27 nests while from 2006 to 2010 there were six failures from 121 nests. The GLM logistic exposure models gave no significant terms including the year 2011*feed zone interaction (parameter estimate = 36.3, z = 0.01, P = 1.00).
Numbers of young fledged
The numbers of young fledged from nests in the two zones are summarized in . There were significantly more young fledged from nests in the fed zone in 2011, but no other significant differences between the zones (Poisson GLM parameter estimate, year 2011*feeding zone = 0.80, z = 2.3, P = 0.02, no other year*feeding zone interactions significant). The number of young fledged was lower in the unfed zone in 2011 than in any other year/treatment.
Seasonal patterns of clutch size and numbers of young fledged
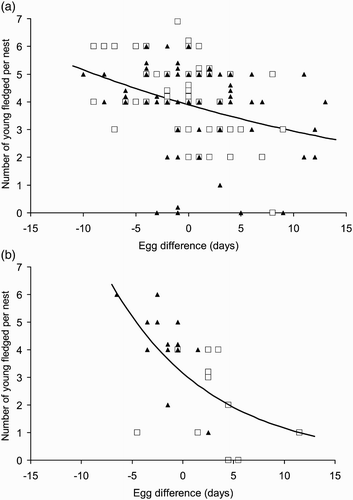
There was no significant relationship between clutch size and egg difference for the overall 2006–2011 data (Poisson GLM parameter estimate, egg difference = 0.017, z = 1.37, P = 0.17) and no significant differences between years (no year*egg difference parameters significant). The numbers of young fledged per nest decreased significantly with egg difference for the overall 2006–2011 data (Poisson GLM parameter estimate, egg difference = 0.035, z = 3.5, P < 0.001), but there were no significant year*egg difference interactions ().
Figure 2. The numbers of young fledged per nest versus egg difference (first egg date for the nest – median laying date for the year). Closed triangles for nests in the fed zone, open squares for nests in the unfed zone. Points are ‘stacked’ where the values overlap. (a) Nests in 2006–2010 when no supplementary food was provided. (b) Nests in 2011 when supplementary food was provided in the fed zone.
DISCUSSION
The results of this experiment are striking. The provision of supplementary food in 2011 resulted in the advancement of laying by 4–5 days and substantially higher productivity. In all years of the study there was a decrease in the number of young fledged per nest with season but in the experimental year the supplemented nests were the earliest ones.
The possible mechanisms involved in the advancement of laying data have been comprehensively reviewed by Harrison et al. Citation(2010). The ‘constraint’ hypothesis suggests that birds are constrained by food supply in their ability to start breeding so that the provision of supplementary food allows them to start earlier. In the case of the Great Spotted Woodpecker, the birds spend a few weeks excavating a nesting cavity before they are able to lay eggs, so this is likely to be a period of particularly high energy demand. The suet blocks used in this study were high in fat and carbohydrate and low in protein and so would be good for supporting the energy demands of cavity excavation, but less so for supplying the amino acids required for egg formation. In our experiment it is difficult to distinguish the predictions of the ‘constraint’ hypothesis from the ‘cue’ hypothesis where the birds are said to be using the availability of food early in the season as a predictor for the timing of the future food peak. However, it is difficult to understand how the ad libitum supply of supplementary food from early February could be used as a predictor of the timing of the invertebrate peak in April and May.
Supplementary feeding may affect the timing of settlement onto a territory or lead to dominant birds settling in the areas with supplementary food. Although they cannot be ruled out entirely, neither of these mechanisms are thought to be important for the Great Spotted Woodpecker in this experiment. There was no evidence of any change in distribution of nests in our wood in response to the supplementary feeding and limited evidence from individually colour-ringed birds suggests they are faithful to their breeding territory from one year to the next (Smith pers. comm.).
Harrison et al. Citation(2010) reported that supplementary feeding led to a reduction in the incubation period of Blue Tits Cyanistes caeruleus and Great Tits Parus major of around one day. If such an effect occurred in our study, our shift in first egg date could have been partly an artefact of back-calculating the first egg date from the fledging date of some nests. To check for any such effects we have calculated the total duration of incubation and chick rearing for the nests found before and during egg laying in 2011. Although the sample sizes are small, there was no significant difference between nests in the fed and unfed zones, indicating that we did not suffer from such an effect (unfed zone, days duration of incubation and chick rearing = 32.8 ± 0.2 se (n = 4); fed zone, days duration = 33.3 ± 0.2 se (n = 7); t = 1.72, P = 0.14).
There are a number of hypotheses to account for the increased productivity of nests in the fed zone (Harrison et al. 2012). One possibility is that the adults are in better condition as a result of the supplementary feeding earlier in the season and so are better able to provision their young. The other explanation is that, by nesting earlier, the birds in the fed zone are better synchronized with the peak in caterpillars, their main prey. It is clear in that in 2011 the caterpillars reached their peak before the woodpeckers were feeding their young, but the birds in the fed zone were better synchronized than those in the unfed. The seasonal decline in productivity observed in all years () lends support to the view that better synchrony was a key driver of the results.
The spring was very warm in 2011 with an April Central England Temperature (Parker et al. Citation1992) of 11.8°C, the highest ever recorded since the series started in 1772. This resulted in a very early and short peak in the caterpillar abundance. In 2011 the frass peak was around 30 April, whereas in Wormley Wood between 2008 and 2010, the years for which the data are available, the peak date ranged from 12 May until 23 May, with a mean of 17 May (Smith et al. Citation2011). Great Spotted Woodpeckers nest earlier in warm years (Smith Citation2006), but the slope of the relationship between median first egg day and Central England Temperature is less steep than that for the date of peak caterpillar abundance (Smith et al. Citation2011). This implies that any mismatch between first egg date and caterpillar peak will tend to be greater as spring temperature increases.
The small advancement in laying date found in this study is similar to those reported in other supplementation studies of temperate woodland birds. Robb et al. Citation(2008) and Harrison et al. Citation(2010) reported advances of a few days for Blue Tits and Great Tits. However, these two studies reported only small effects on productivity (positive in the case of Robb et al. but negative for Harrison et al.). The difference in our study is that the small advancement of laying date has resulted in a big increase in breeding success. This may have been the result of the particularly warm spring in 2011, but our study also shows the importance of understanding both the abundance and temporal patterns of natural food resources in interpreting the results of supplementary feeding experiments.
The subject of the phenology mismatch between the timing of breeding and prey availability has received considerable attention in relation to the effects of climate change on breeding productivity (see Both Citation2010 for a review). The impact of the mismatch is thought to be particularly important for long-distance migrants which may be unable to adjust their annual cycle to respond to temporal shifts in prey availability in the breeding areas. For resident species the results are more mixed, with some showing evidence of disjuncture and others not. For instance, Great Tit populations in different regions show different responses (Visser et al. Citation1998, Citation2006, Charmantier et al. Citation2008). The results of our experiment suggest that the degree to which the resident Great Spotted Woodpecker can advance its laying date is constrained by food availability so that in some years with high spring temperatures it will suffer from mismatch.
Great Spotted Woodpeckers are increasing in abundance and one of the suggested mechanisms is increased survival or breeding success as the result of supplementary feeding in gardens. The frequency of birds visiting gardens has increased (Cannon et al. Citation2005) and, although they peak in June, they are present throughout the year, being reported from around 22% of BTO Garden Birdwatch sites in May (http://www.bto.org/volunteer-surveys/gbw/results). This experiment has shown the potential for there to be a positive effect on the timing of breeding and breeding success but it is not clear whether sufficient birds have access to and visit garden feeders for there to be a population level response. It is probable that Great Spotted Woodpeckers breeding within woods will visit feeders in adjacent gardens and it is also apparent that the birds are able to breed in suburban areas provided there are mature trees to provide nest sites. Although they show strong selection for excavating nest cavities in standing dead trees, even in woodlands, about half of the nests are in live mature trees (Smith Citation1997). Further work is needed, not only to quantify the fraction of the woodpecker population that accesses garden feeders during the breeding season, but also to look at other aspects of this question such as the impact of supplementary feeding at different times of year on over winter survival and juvenile recruitment, which may be even more important factors.
ACKNOWLEDGEMENTS
Wormley Wood is owned and managed by the Woodland Trust and we thank them and their local officer Louise Neicho for support and encouragement throughout this study. Tim Harrison, Will Cresswell and an anonymous referee are thanked for their constructive comments on an earlier draft of this paper.
REFERENCES
- Arcese , P. and Smith , J. N.M. 1988 . Effects of population density and supplemental food on reproduction in song sparrows . J. Anim. Ecol , 57 : 119 – 136 . (doi:10.2307/4768)
- Baillie , S. R. , Marchant , J. H. , Leech , D. I. , Renwick , A. R. , Eglington , S. M. , Joys , A. C. , Noble , D. G. , Barimore , C. , Conway , G. J. , Downie , I. S. , Risely , K. and Robinson , R. A. 2012 . Bird Trends 2011 , BTO Research Report No. 609. BTO, Thetford
- Both , C. 2010 . “ Food availability, mistiming and climatic change ” . In Effects of Climate Change on Birds , Edited by: Moller , A. P. , Fiedler , W. and Berthold , P. Oxford : Oxford University Press .
- Cannon , A. R. , Chamberlain , D. E. , Toms , M. P. , Hatchwell , B. J. and Gaston , K. J. 2005 . Trends in the use of private gardens by wild birds in Great Britain 1995–2002 . J. Appl. Ecol. , 42 : 659 – 671 . (doi:10.1111/j.1365-2664.2005.01050.x)
- Castro , I. , Brunton , D. H. , Mason , K. M. , Ebert , B. and Griffiths , R. 2003 . Life history traits and food supplementation affect productivity in a translocated population of the endangered hihi (stitchbird, Notiomystis cincta) . Biol. Conserv. , 114 : 271 – 280 . (doi:10.1016/S0006-3207(03)00046-6)
- Charmantier , A. , McCleery , R. H. , Cole , L. R. , Perrins , C. , Kruuk , L. E.B. and Sheldon , B. C. 2008 . Adaptive phenotypic plasticity in response to climate change in a wild bird population . Science , 320 : 800 – 803 . (doi:10.1126/science.1157174)
- Fuller , R. J. , Noble , D. G. , Smith , K. W. and Vanhinsbergh , D. 2005 . Recent declines in populations of woodland birds in Britain: a review of possible causes . Br. Birds , 98 : 116 – 143 .
- Harrison , T. J.E , Smith , J. A. , Martin , G. R. , Chamberlain , D. E. , Bearhop , S. , Robb , G. N. and Reynolds , S. J. 2010 . Does food supplementation really enhance productivity of breeding birds? . Oecologia , 164 : 311 – 320 . (doi:10.1007/s00442-010-1645-x)
- Houston , D. , McInnes , K. , Elliot , G. , Eason , D. , Moorhouse , R. and Cockrem , J. 2007 . The use of a nutritional supplement to improve egg production in the endangered kakapoo . Biol. Conserv , 138 : 248 – 255 . (doi:10.1016/j.biocon.2007.04.023)
- Jackson , J. A. 1976 . How to determine the status of a woodpecker nest . Living Bird , 15 : 205 – 221 .
- Källander , K. and Karlsson , J. 1993 . Supplemental food and laying date in the European starling . Condor , 95 : 1031 – 1034 . (doi:10.2307/1369440)
- Kelly , J. F. and Van Horne , B. 1997 . Effects of food supplementation on the timing of nest initiation in belted kingfishers . Ecology , 78 : 2504 – 2511 . (doi:10.1890/0012-9658(1997)078[2504:EOFSOT]2.0.CO;2)
- Minitab 16 Statistical Software . 2010 . State College, PA: Minitab, Inc. (www.minitab.com)
- Newton , I. 1998 . Population Limitation in Birds , London : Academic Press .
- Parker , D. E. , Legg , T. P. and Folland , C. K. 1992 . A new daily Central England temperature series, 1772–1991 . Int. J. Climatol. , 12 : 317 – 342 . (doi:10.1002/joc.3370120402)
- R Core Team . 2012 . R: A Language and Environment for Statistical Computing , R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/
- Robb , G. N. , McDonald , R. A. , Chamberlain , D. E. , Reynolds , S. J. , Harrison , T. J.E. and Bearhop , S. 2008 . Winter feeding of birds increases productivity in the subsequent breeding season . Biol. Lett , 4 : 220 – 223 . (doi:10.1098/rsbl.2007.0622)
- Shaffer , T. L. 2004a . A unified approach to analysing nest success . The Auk , 121 : 526 – 540 .
- Shaffer , T. L. 2004b . Logistic-Exposure Analyses of Nest Survival , Jamestown , ND : Northern Prairie Wildlife Research Center Online . http://www.npwrc.usgs.gov/resource/birds/nestsurv/index.htm (Version 28AUG2008)
- Schoech , S. J. , Bridge , E. S. , Boughton , R. K. , Reynolds , S. J. , Atwell , J. W. and Bowman , R. 2008 . Food supplementation: a tool to increase reproductive output? A case study in the threatened Florida scrub jay . Biol. Conserv , 141 : 162 – 173 . (doi:10.1016/j.biocon.2007.09.009)
- Smith , K. W. 1997 . Nest site selection of the great spotted woodpecker Dendrocopos major in two oak woods in southern England and its implications for woodland management . Biol. Conserv , 80 : 283 – 288 . (doi:10.1016/S0006-3207(96)00038-9)
- Smith , K. W. 2005 . Has the reduction in nest-site competition from Starlings Sturnus vulgaris been a factor in the recent increase of Great Spotted Woodpecker Dendrocopos major numbers in Britain? . Bird Study , 52 : 307 – 313 . (doi:10.1080/00063650509461404)
- Smith , K. W. 2006 . The implications of nest site competition from starlings Sturnus vulgaris and the effect of spring temperatures on the timing and breeding performance of great spotted woodpeckers Dendrocopos major in southern England . Ann. Zool. Fennici , 43 : 177 – 185 .
- Smith , K. W. , Smith , L. , Charman , E. , Briggs , K. , Burgess , M. , Dennis , C. , Harding , M. , Isherwood , C. , Isherwood , I. and Mallord , J. 2011 . Large-scale variation in the temporal patterns of the frass fall of defoliating caterpillars in oak woodlands in Britain: implications for nesting woodland birds . Bird Study , 58 : 506 – 511 . (doi:10.1080/00063657.2011.616186)
- Smith , K. W. and Charman , E. C. 2012 . The ecology and conservation of the Lesser Spotted Woodpecker . Br. Birds , 105 : 294 – 307 .
- Soler , M. and Soler , J. J. 1996 . Effects of experimental food provisioning on reproduction in the Jackdaw Corvus monedula, a semi-colonial species . Ibis , 138 : 377 – 383 . (doi:10.1111/j.1474-919X.1996.tb08054.x)
- Visser , M. E. , van Noordwijk , A. J. , Tinbergen , J. M. and Lessells , C. M. 1998 . Warmer springs lead to mistimed reproduction in great tits (Parus major) . Proc. R. Soc. Lond. B , 265 : 1867 – 1870 . (doi:10.1098/rspb.1998.0514)
- Visser , M. E. , Holleman , L. J.M. and Gienapp , P. 2006 . Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird . Oecologia , 147 : 167 – 172 . (doi:10.1007/s00442-005-0299-6)