
Abstract
Capsule Skylarks and Yellowhammers generally did not preferentially establish territories in flax relative to autumn-sown crops on lowland farmland.
Aims To investigate the biodiversity value of flax using farmland birds as indicator species.
Methods Field surveys took place annually between 2009 and 2011 in Northern France, during the breeding season. Fifteen 1 km grid squares were surveyed using a grid-transect method during the periods of greatest avian activity. Territories were identified by the presence of singing males, which were assigned to crop types. Pairwise habitat comparisons were performed.
Results Our results suggest that flax is not universally beneficial for Yellowhammers and Skylarks in our study area. However there were inter-specific differences in its use: Skylarks established more territories in flax than in oilseed rape, ploughed fields or improved grassland, whereas Yellowhammers mostly avoided flax fields in preference for grassland, ploughed fields and barley.
Conclusion Overall, Skylarks and Yellowhammers did not preferentially establish territories in flax relative to autumn-sown crops, although some selection for flax by Skylarks was evident. Further work is required to: (a) quantify breeding success in flax relative to other crops and (b) investigate the effect of flax on territory densities of other species.
Loss of biodiversity on agricultural land is a major European conservation issue (Gregory et al. Citation2004). Birds are charismatic and easily surveyed, and simultaneously represent both targets and indicators of environmental change, particularly of agri-systems (Ormerod & Watkinson Citation2000). A dramatic decline in common farmland bird populations across Northern and Western Europe over the last few decades is well documented, as detailed by the European Union Farmland Bird Index that lists 37 species of which 20 currently have unfavourable conservation status (European Bird Census Council (EBCC) Citation2008). There is substantial and compelling evidence to link the declines to various aspects of agricultural intensification (Aebischer et al. Citation2003, Vickery et al. Citation2004, Donald et al. Citation2006) including a shift from spring to autumn-sown crops (Donald et al. Citation2001), lower diversity of crops and arable weeds (Chamberlain et al. Citation2000), and reduced winter seed availability (Evans et al. Citation2010). However, effects may be ecologically complex and contradictory between species due to specialist requirements. Agri-environment schemes aimed at halting the negative effects of modern farming practices on bird conservation status have not been overwhelmingly successful, and their effects have been shown to vary regionally (Davey et al. Citation2010a, Citationb, Field et al. Citation2011, Princé et al. Citation2012).
Flax Linum usitatissimum is a hardy herbaceous annual cultivated for food and fibre and extensively grown as a cash crop in Northern and Eastern Europe, Russia, Belgium and the Normandy region of France. Most varieties of flax are spring-sown, and in Normandy sowing generally occurs by late April. Flax has a short growing cycle: harvested fibres are left to rot on the ground for 10–20 days during late summer/early autumn (a process known as retting), after which fields lie fallow or are replanted with other crops. In general flax has a low fertilizer, herbicide and pesticide requirement relative to crops more commonly grown across Europe such as wheat, barley and oilseed rape (Defra Citation2005). It has been suggested that the inclusion of flax in arable rotations could be beneficial for farmland birds (Defra Citation2005) because it provides a nesting environment that is less dense than autumn-sown crops during mid-summer when pairs are attempting a second brood. Furthermore, seed residues during the retting period may provide high-energy food for granivorous birds when traditional stubble crops are no longer available. However, early ground-nesting attempts may be disrupted by the sowing of flax, especially if this occurs late due to unseasonal weather. In addition, retted flax fibres may provide inadequate cover relative to cereal stubbles.
Numerous published studies, particularly from the UK, report bird habitat selection within an agricultural landscape (for a review see Ormerod & Watkinson Citation2000) and the value of un-cropped land in arable rotations has been well demonstrated (see Filippi-Codaccioni et al. Citation2010, Henderson et al. Citation2012). However, to our knowledge, no studies have explicitly focused on the role of ‘unusual’ crops of European farmland such as flax, which are nonetheless integral to the landscape. Such crops may provide spatio-temporal niches that differ from other habitat patches and as such may be important components of farmland ecology at the local and regional scale.
Habitat selection is commonly measured using relative density of breeding territories. Here we use relative territory density of two common but declining songbirds: Skylark Alauda arvensis and Yellowhammer Emberiza citrinella in mixed lowland farmland in Normandy, to provide a preliminary investigation into the relative biodiversity value of flax cultivation. We chose these species because they are conspicuous, sedentary and mono-territorial and are therefore easily monitored in relation to habitat use. Specifically, we test the hypothesis that territories will occur disproportionately frequently in flax relative to its availability as compared to autumn-sown cereals. The study forms part of a preliminary environmental impact assessment for a feasibility study into the expansion of flax production in the Southeast of the UK, which is of similar latitude to Normandy and is comparable in terms of geology, climate and socio-economics.
METHODS
Study sites
The study area encompassed an area of approximately 30 km2 of lowland farmland situated in the department of Seine-Maritime in the region of Haute-Normandie, Northern France (49.6667° N, 0.8333° E). Throughout the area flax is incorporated into arable rotations on a three to five yearly basis. Selection of a total of 15 1 km grid squares was conducted using 1:25 000 maps (Carte de Randonnée, Série bleue, Institut Geographique National) according to the following criteria: (a) at the time of survey the area planted with flax totalled at least 20% of the square and; (b) flax cultivated within the square was subject to a similar regime in terms of fertilizer and herbicide/pesticide applications. All sites comprised mixed farmland bounded by hedgerows, small copses and deciduous or mixed woodland. Arable land was planted with flax, cereals (wheat, barley, oats) maize, sugar beet, brassicas (oilseed rape, cabbage) and legumes (beans, peas) inter-dispersed with improved or semi-improved (ley) pasture predominantly grazed by cattle and horses, plus smaller areas of fallow land and game cover.
Survey protocol
Two observers conducted field surveys in the early summer of 2009, 2010 and 2011. Surveys took place three months after flax was sown, ranging from 1 June to 7 July, because sowing date varied annually according to weather conditions. Five different 1 km grid squares were surveyed every year, over a period of three to five days, representing approximately 30 man-hours of survey effort per year, and totalling 90 man-hours and 15 1 km squares over the whole study period. Surveys were conducted during the periods of greatest avian activity, i.e. between 07:00 and 10:00 and 18:00 and 20:00 hours, using the grid-transect method described by Brown & Shepherd (Citation1993) adapted for lowland farmland. A single observer followed a randomized route on foot through each survey square, such that all parts of the square could be observed and/or approached to within 100 m, maintaining a constant search effort of approximately 30 minutes spent in each 250 m2 section. The observer regularly (approximately every 100 m) scanned the terrain using binoculars, and listened for calls. Surveys were not conducted in winds of > 4 on the Beaufort scale or in very wet or cold weather, or conditions of poor visibility. Where sampling sessions commenced during favourable weather conditions that subsequently deteriorated, data were excluded for the period of poor weather.
Positions of territories were inferred by the presence of singing male Skylarks (Chamberlain et al. Citation1999) and Yellowhammers (Bradbury et al. Citation2000) and recorded onto 1:25 000 maps. Each map had previously been annotated into classifications of habitat and crop type (e.g. arable, barley) during land use surveys, normally conducted the day before each bird survey. Classifications were defined according to the UK Phase 1 habitat survey guidelines (Joint Nature Conservation Committee (JNCC) Citation2010). During year three of the study, the height of all crop types was additionally recorded (as estimated by eye), because crop preferences may be associated with vegetation height, e.g. Skylarks tend to avoid tall crops (Wilson et al. Citation1997). Crop height was assigned to five categories, as modified from Chamberlain et al. Citation1999 (< 10 cm; 10–30 cm; 30–50 cm; 50–1 m; > 1 m).
Because Yellowhammers are known to sing from hedgerows and trees, we assigned Yellowhammer territories to the crop type within which, or adjacent to where occupied hedgerows/tree stands were located. Where occupied hedgerows or trees bordered two or more crop types, a territory was assigned to both. Although it was possible that males displaying over a given crop type held territories that also contained multiple other habitats thereby biasing the results, small mean territory sizes are reported for Skylarks – e.g. 2.62 ha (Poulsen et al. Citation1998) or two fields (Wilson et al. Citation1997) hence we could be confident that the majority of territories predominantly contained the recorded habitat.
Data analysis
All statistical analyses were computed using R (Version 2.13: R Foundation for Statistical Computing, Vienna, Austria).
Counts of number of breeding territories for each species were regressed against habitat type using multiple log-linear regression analyses within generalized linear models (GLM). Year of study (with 3 levels) was included as a fixed factor. Crop height (with 5 levels) was incorporated into a separate, reduced GLM for year 3 only that also contained all other variables. Site (with 15 levels) was additionally included as a fixed factor to account for the variance in the response between sample locations that could be explained by factors other than habitat (see Chamberlain et al. Citation1999). To control for variation in crop area and thus differential availability of habitats within the squares, the total proportion covered by each crop/habitat was entered into the models as an offset (see Chamberlain et al. Citation1999). This allowed any disproportional occupancy of habitat patches to be assumed to represent selection. Two-way interaction terms between all of the main effects were initially included in the models, however these were removed during the model selection procedure because they did not reach convergence and so were not considered further.
The ratio between residual degrees of freedom and residual mean deviance did not exceed 2 for any of the analyses hence response variables were assumed to be best-fitted to a Poisson distribution.
Screening for collinearity between pairs of explanatory variables (e.g. bivariate combinations of site, year, habitat type and crop height) was conducted using Cramér's V test. Where any two variables were collinear and both retained significance at the 5% level in subsequent regression analysis, the variable accounting for the least variance in the response was excluded from the final model. Backwards selection was then used to determine the minimum adequate model (MAM) that included significant terms at the 5% level, identified using Akaike's Information Criterion (AIC) and percentage change in deviance values. Analysis of variance was used to determine whether the MAM and the next best-fit model were significantly different for each species. Because stepwise regression can generate conflicting results according to the direction in which the procedure is computed (James & McCulloch Citation1990), we also used forwards selection, and compared the results. The MAM for both the full and reduced dataset retained identical terms when generated using forwards and backwards selection, hence we conclude that our methods were robust.
Territory counts for Skylarks from site 10 (Bras d'Or) surveyed in 2011 were anomalously high (11 in flax in one grid square) relative to other sites and previous years. The flax field in this square was unusually large (> 50% of the square) and was subject to extensive excavation work creating numerous wide tramlines and mounds of soil. Subsequently, reduced GLMs were re-run with this site excluded to determine its relative influence.
RESULTS
A total of 93 Skylark and 52 Yellowhammer breeding territories were recorded within 13 crop types of differing patch sizes and cropping regimes (). For the full Skylark dataset 24% of males displayed over flax (n = 22); 20% over each of wheat and barley (n = 22 for both); 13% over sugar beet (n = 12) and 10% over maize (n = 9), with the remainder located in beans, pasture, potatoes or ploughed fields. With site 10 removed, only 13% displayed over flax, leaving larger proportions in wheat and barley (22%), sugar beet (15%) and maize (11%). 62% of male Yellowhammers sang from hedgerows or tree stands (n = 32), with the remainder singing from the ground within crops (n = 3), telegraph wires (n = 4), the edge of deciduous or mixed woodland (n = 3), fence-posts (n = 4), excavation mounds (n = 5) and an electricity box (n = 1).
Table 1. Crops and other habitat types in which Skylark Alauda arvensis and Yellowhammer Emberiza citrinella established breeding territories, showing mean size of habitat patch and cropping regime for each.
Habitat significantly predicted number of territories of breeding Skylark for both datasets. Skylarks were significantly more likely to hold a territory in flax than in oilseed rape (full dataset only), semi-improved grassland and improved grassland, although all other comparisons were equal (; ). Site remained in the MAM for the full dataset but became non-significant following removal of site 10. The MAM was significantly different to the next best-fit model for the full dataset (GLM, one-way ANOVA, df = 1, Δ deviance χ2 = 32.5, P < 0.01). Habitat type and site significantly predicted the number of territories for Yellowhammers (), and the MAM was significantly different to the next best-fit model (GLM, one-way ANOVA, df = 1, Δ deviance χ2 = 23.3, P < 0.05). Yellowhammer territories were significantly less likely to occur in flax than in barley, semi-improved and improved grassland or ploughed fields, but other comparisons were equal (; ).
Figure 1. Densities of breeding territories of Skylarks Alauda arvensis in different crop/habitat types. SI = semi-improved, I = improved. Numbers with asterisks indicate significant differences relative to flax: *P < 0.05; **P < 0.01;***P < 0.001. Sample sizes (n=) represent number of grid squares containing each habitat type.
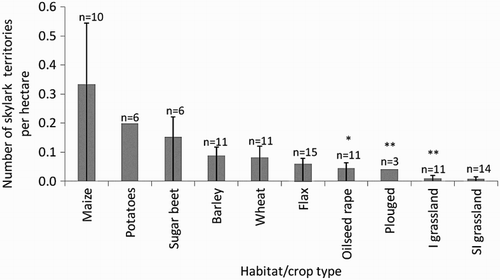
Figure 2. Densities of breeding territories of Yellowhammers Emberiza citrinella in different crop/habitat types. SI = semi-improved, I = improved. Numbers with asterisks indicate significant differences relative to flax: *P < 0.05; **P < 0.01;***P < 0.001. Sample sizes (n=) represent number of grid squares containing each habitat type.
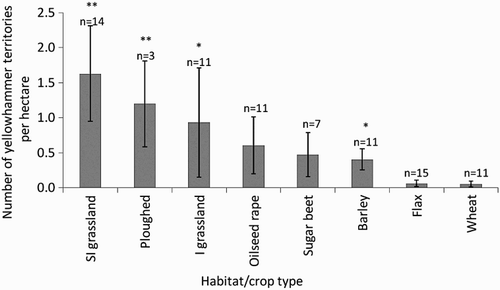
Table 2. Results of multiple log-linear regression analysis (GLM) predicting number of Skylark (F = full dataset and R = reduced dataset) and Yellowhammer breeding territories. Δ deviance values represent the change in deviance after each term is removed from the model. MLPE from post-hoc tests. Positive and negative MLPE values indicate the direction of the difference between each of the habitat categories listed (I grassland = Improved grassland and SI grassland = Semi-improved grassland), and flax (e.g. for Yellowhammers there were more territories in Improved grassland than in Flax). Data for flax in year 2 site 10 (Bras d’Or).
For the crop height data subset, there were no effects of either crop height (GLM, maximum likelihood parameter estimate (MLPE) = 2.6, df = 4, P > 0.05) or site (GLM, MLPE = 11.3, df = 7, P > 0.05) on number of Yellowhammer territories, although habitat remained significant (GLM, MLPE = 24.2, df = 10, P < 0.01). Site, crop height and habitat predicted number of Skylark territories, but crop height and habitat were collinear (Cramér's V test, V2 = 0.73, df = 1, P < 0.01). Crop height was removed from the model on the basis that it had a weak effect, explaining only 8.4% of the deviance in the response, compared to 32.0% attributed to habitat differences. There was no effect of year of study on territory numbers for either species (GLM, Δ deviance = 6.3, df = 2, P > 0.05)
DISCUSSION
Skylarks and Yellowhammers generally did not preferentially establish territories in flax relative to autumn-sown crops. Flax was selected more frequently by Skylarks than oilseed rape but was less preferred by Yellowhammers than barley, with all other comparisons being equal. Unlike the results of Bradbury et al. (Citation2000), Yellowhammers did not unanimously avoid flax, although territory density was low relative to several other habitat types, particularly non-intensively managed grassland. In contrast, Skylarks were observed to display over flax more often than grassland of any kind, and this relationship persisted when Bras d'Or was excluded from analyses.
The disproportionately high rate of Skylark occupancy at Bras d'Or concurs with Oddeskær et al. (Citation1997) who report a strong preference for tramlines by radio-tracked Skylarks in a barley field in late summer, and a positive correlation between crop height and the proportion of Skylarks located in tramlines. Eggers et al. (Citation2011) report that Skylark preferences for vegetation with shorter swards results in a switch in crop selection from autumn-sown to spring-sown cereals as the summer progresses. However the limited influence of crop height on territory densities in our study suggests that crop selection is not entirely due to sward height, with the caveat that our data are derived from one year only and therefore may include some bias. Additional factors may influence decision-making by ground-nesting species; potentially including crop density (Donald Citation2004) availability of seeds and invertebrates, and degree of anthropogenic disturbance.
Regional pesticide and herbicide regimes are likely to vary substantially, making generalizations about relative input difficult. In the current study flax was sprayed less frequently over the annual cycle than wheat, barley and oilseed rape, but this could be an artefact of its shorter growing season (Laurent Bizet pers. comm.) and may therefore not translate into beneficial effects. The latter may include higher chick survivorship as a consequence of increased availability of invertebrate food items.
Suggestions for further work include: (a) repeat surveys of the study sites to obtain detection histories (i.e. occupancy analysis) and enable territories to be defined in more detail; (b) incorporation of field size into survey protocols; (c) concurrent measurement of breeding success and invertebrate food availability; and (d) surveys of additional species that may occupy different ecological niches on farmland.
FUNDING
This work was funded within the framework of the Interreg IVA France (Manche) – England Operational Programme, project 4044 ‘Flax – Increasing Its Value for Society’.
REFERENCES
- Aebischer, N.J., Bradbury, R., Eaton, M., Henderson, I.G., Siriwardena, G.M. & Vickery, J.A. 2003. Predicting the response of farmland birds to agricultural change. BTO Research Report 289, British Trust for Ornithology, Thetford.
- Bradbury, R.B., Kyrkos, A., Morris, A.J., Clark, S.C., Perkins, A.J. & Wilson, J.D. 2000. Habitat associations and breeding success of yellowhammers on lowland farmland. J. Appl. Ecol. 37: 798–805. doi: 10.1046/j.1365-2664.2000.00552.x
- Brown, A.F. & Shepherd, K.B. 1993. A method for censusing upland breeding waders. Bird Study 40: 89–195.
- Chamberlain, D.E., Wilson, A.M., Browne, S.J. & Vickery, J.A. 1999. Effects of habitat type and management on the abundance of skylarks in the breeding season. J. Appl. Ecol. 36: 856–870. doi: 10.1046/j.1365-2664.1999.00453.x
- Chamberlain, D.E., Fuller, R.J., Bunce, R.G.H., Duckworth, J.C. & Shrubb, M. 2000. Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. J. Appl. Ecol. 37: 771–788. doi: 10.1046/j.1365-2664.2000.00548.x
- Davey, C.M., Vickery, J.M., Boatman, N.D., Chamberlain, D.E., Parry, H.R. & Siriwardena, G.M. 2010a. Assessing the impact of entry level stewardship on lowland farmland birds in England. Ibis 152: 459–474. doi: 10.1111/j.1474-919X.2009.01001.x
- Davey, C.M., Vickery, J.M., Boatman, N.D., Chamberlain, D.E., Parry, H.R. & Siriwardena, G.M. 2010b. Regional variation in the efficacy of entry level stewardship in England. Agr. Ecosyst. Environ. 139: 121–128. doi: 10.1016/j.agee.2010.07.008
- Department for Environment, Food and Rural Affairs (Defra) 2005. ADAS report: UK flax and hemp production: the impact of changes in support measures on the competitiveness and future potential of UK fibre production and industrial use. DEFRA, HMSO, 2005. 65 pp. Accessed 19 November 2013 at: http://archive.defra.gov.uk/foodfarm/growing/crops/industrial/pdf/flaxhemp-report.pdf
- Donald, P.F. 2004. The Skylark. T. & A.D. Poyser, London.
- Donald, P.F., Buckingham, D.L., Moorcroft, D., Muirhead, L.B., Evans, A.D. & Kirkby, W.B. 2001. Habitat use and diet of skylarks Alauda arvensis wintering on lowland farmland in southern Britain. J. Appl. Ecol. 38: 536–547. doi: 10.1046/j.1365-2664.2001.00618.x
- Donald, P.F., Sanderson, F.J., Burfield, I.J. & van Bommel, F.P.J. 2006. Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agr. Ecosyst. Environ. 116: 189–196. doi: 10.1016/j.agee.2006.02.007
- Eggers, S., Unell, M. & Pärt, T. 2011. Autumn-sowing of cereals reduces breeding bird numbers in a heterogeneous agricultural landscape. Biol. Cons. 144: 1137–1144. doi: 10.1016/j.biocon.2010.12.033
- European Bird Census Council (EBCC) 2008. The state of Europe's common birds. Accessed 19 November 2013 at: http://www.ebcc.info/wpimages/video/SECB2008.pdf.
- Evans, D.M., Pocock, M.J., Brooks, J. & Memmott, J. 2010. Seeds in farmland food-webs: resource importance, distribution and the impacts of farm management. Biol. Cons. 144: 2941–2950. doi: 10.1016/j.biocon.2011.08.013
- Field, R.H., Morris, A.J., Grice, P.V. & Cooke, A. 2011. The provision of winter bird food by the English Environmental Stewardship Scheme. Ibis 153: 14–26. doi: 10.1111/j.1474-919X.2010.01087.x
- Filippi-Codaccioni, O.D.V., Bas, Y., Clobert, J. & Julliard, R. 2010. Specialist response to proportion of arable land and pesticide input in agricultural landscapes. Biol. Cons. 143: 883–890. doi: 10.1016/j.biocon.2009.12.035
- Gregory, R.D., Noble, D.G. & Custance, J. 2004. The state of play of farmland birds; population trends and conservation status of lowland farmland birds in the United Kingdom. Ibis 146: 1–14. doi: 10.1111/j.1474-919X.2004.00358.x
- Henderson, I.G., Holland, J.M., Storkey, J., Lutman, P., Orson, J. & Simper, J. 2012. Effects of the proportion and spatial arrangement of un-cropped land on breeding bird abundance in arable rotations. J. Appl. Ecol. 49: 883–891. doi: 10.1111/j.1365-2664.2012.02166.x
- James, F.C. & McCulloch, C.E. 1990. Multivariate analysis in ecology and systematics: panacea or Pandora's box? Annu. Rev. Ecol. Syst. 21: 129–166.
- Joint Nature Conservation Committee (JNCC) 2010. Handbook for phase 1 habitat survey – a technique for environmental audit. Accessed 19 November 2013 at: http://jncc.defra.gov.uk/PDF/pub10_handbookforphase1habitatsurvey.pdf
- Oddeskær, P., Prang, A., Poulsen, J.G., Normark, P., Anderson, C. & Elrnegaard, N. 1997. Skylark (Alauda arvensis) utilisation of micro-habitats in spring barley fields. Agr. Ecosyst. Environ. 62: 21–29. doi: 10.1016/S0167-8809(96)01113-9
- Ormerod, S.J. & Watkinson, A.R. 2000. Special profile: birds and agriculture. Editors’ introduction: birds and agriculture. J. Appl. Ecol. 37: 699–705. doi: 10.1046/j.1365-2664.2000.00576.x
- Poulsen, J.G., Sotherton, N.W. & Aebischer, N.J. 1998. Comparative nestling and feeding ecology of skylarks Alauda arvensis on arable farmland in southern England with special reference to set-aside. J. Appl. Ecol. 35: 131–147. doi: 10.1046/j.1365-2664.1998.00289.x
- Princé, K., Moussus, J.P. & Jiguet, F. 2012. Mixed effectiveness of French agri-environment schemes for nationwide farmland bird conservation. Agr. Ecosyst. Environ. 149: 74–79. doi: 10.1016/j.agee.2011.11.021
- Vickery, J.A., Bradbury, R.B., Henderson, I.G., Eaton, M.A. & Grice, P.V. 2004. The role of agri-environment schemes and farm management practices in reversing the decline of farmland birds in England. Biol. Cons. 119: 19–39. doi: 10.1016/j.biocon.2003.06.004
- Wilson, J.D., Evans, J., Browne, S.J. & King, J.R. 1997. Territory distribution and breeding success of skylarks Alauda arvensis on organic and intensive farmland in southern England. J. Appl. Ecol. 34: 1462–1478. doi: 10.2307/2405262