
Abstract
Capsule Woodland structure, rather than tree species, is the most important determinant of breeding habitat selection by Willow Warblers in North West England.
Aims To examine how habitat characteristics predict the occurrence of male Willow Warbler territories.
Methods Woodland structure (trunk density, trunk diameter, canopy cover and understory cover), tree species and food abundance were compared between woodland areas within and outside of male territories at a site in the UK.
Results Territories contained higher trunk numbers, had a narrow range of trunk diameters, and intermediate canopy cover. Food abundance did not differ with occupancy. Willow and alder were the most common trees within territories, in contrast to birch which has been found in previous studies. The habitat structure matches young woodlands, where birches often grow. However, at the study site the birches were large and mature, and therefore unsuitable. Moreover, woodland structure variables were better predictors of occupancy than any particular tree genera.
Conclusion The results indicate that vegetation structure, but not tree species or food availability, influence breeding habitat selection by Willow Warblers. The preferred structure is similar to coppice woodlands; therefore, the Willow Warbler decline may be linked to the loss of this traditional management across south England.
Willow Warblers (Phylloscopus trochilus) are small, migratory, passerine birds that breed throughout the Western Palearctic (Cramp & Brooks Citation1992). Similarly to other trans-Saharan migratory birds, Willow Warbler populations in the UK have declined over the past few decades and they are now a focus for conservation research (Fuller et al. Citation2005, Morrison et al. Citation2010). However, such declines are not uniform throughout the UK. Willow Warblers are reported to be doing well in northern parts of England and in Scotland whereas the southern UK populations are declining (Amar et al. Citation2006, Risely et al. Citation2008). Although the decline may be due to issues during migration, other aspects of Willow Warbler ecology, including their breeding habitats may be important. Therefore, it is important to identify the characteristics of successful Willow Warbler territories so that appropriate local conservation actions can be implemented.
Willow Warbler habitat is generally outlined as young plantations, forest clearings and woodland edges (Cramp 1992, Holden & Cleeves Citation2010), with broadleaf and deciduous trees being preferred (Tiainen et al. Citation1983, Felton et al. Citation2011). Several other studies report a tendency for specific selection of birch trees (Betula spp.) throughout the UK, compared to a variety of other tree species (Yapp Citation1962, Cramp 1992, Fuller & Henderson Citation1992, Mason Citation1997, Gillings et al. Citation1998). In their winter habitats in Zimbabwe, they seem to prefer Acacia woodland, but they also occur in Miombo (Brachystegia sp.) and Mopane (Colophospermum mopane) woodlands, indicating that they are flexible in the tree species in which they choose to forage (Jones et al. Citation2010).
Alternatively, there are indications of territory choices being made on the basis of vegetation structure alone, rather than tree species in itself. For example, Bellamy et al. (Citation2009) found that Willow Warblers preferred a mean vegetation height of 3.7–5.3 m, regardless of tree species. An extensive survey by Amar et al. (Citation2006) found positive correlations with low vegetation cover, moss cover and bramble cover, yet a negative correlation with canopy cover. Both of the above studies indicated that Willow Warbler males tend to establish territories in young woodland areas. Meanwhile, Arvidsson & Klaesson (Citation1986) found no correlation between male territory size and food abundance, also indicating that males compete mainly for other resources than food, although this was in contrast to earlier research (Nilsson & Ebenman Citation1981). Despite these findings, little is still known about the habitat preference of Willow Warblers in the northern part of its range in the UK (but see Gillings et al. Citation1998). This omission in our knowledge of the species requirements is of potential concern, because species habitat requirements may not be uniform across a species' geographic range due to interspecific competition and changes in resource availability and climatic conditions (Fuller Citation2002). Therefore, it is not altogether clear whether territory selection is made based on (i) the abundance of food, (ii) the structure of the vegetation, or (iii) the tree species composition of an area, and how their importance varies across the Willow Warbler's range. Consequently, this study aimed to investigate the relative influence of these three factors on the selection of breeding territories by male Willow Warblers at a site in the North West of England.
METHODS
Study area
The study was performed in cooperation with Watchtree Ltd., at Watchtree Nature Reserve (WNR), Cumbria, UK (54.88° N, 3.08° W), in May–July 2012. The area was a mass burial site after the Foot and Mouth disease in 2001, and this took up parts of the woodland, which was replanted after the burial event. The nature reserve covers more than 80 ha of varying habitat, with hay meadows, woodland and wetland areas now managed for wildlife conservation. The woodland occupies roughly 25% (20 ha) of the reserve, about half of which is mature woodland from before the burial event and the rest planted as a diverse broadleaf/coniferous mix on the burial grounds after 2001.
Territory borders and sampling quadrats
Forty quadrats, each measuring 5 × 5 m, were placed in the woodland areas of the nature reserve, excluding areas not containing trees. Twenty quadrats were situated in areas which were part of male Willow Warbler territories (‘territory quadrats’); the other 20 were in areas outside Willow Warbler territories (‘non-territory quadrats’). During the breeding season, Willow Warbler territories are annually surveyed and mapped by staff of the nature reserve, following the methodology of the British Trust for Ornithology's Common Bird Census (Marchant Citation1983), using observations of males from nine visits to create maps of territories. All sampling quadrats were marked in the middle of the breeding season May–June in 2012 when most territories were already stabilized. Territory quadrats were selected in areas known to be occupied by birds in the middle of current season, with one quadrat per territory. As some areas can be occupied later in the season by late arriving birds, non-territory quadrats were selected randomly in areas known to have not been occupied by birds in the previous year (Still Citation2011) and confirmed at the end of the season using the territory map for the current season (Still Citation2012).
Habitat surveys
Several habitat variables were measured within each quadrat to assess woodland structure (trunk diameter, trunk density, tree distance, canopy and understory cover) and woodland tree composition (tree species and abundance of each tree species). Trunk diameter was measured as standard diameter at breast height for all trunks larger than 1 cm in diameter and the average was calculated per quadrat. The number of trunks (>1 cm diameter) per quadrat (‘trunk density’) was also recorded. All trunks from a single root were measured separately for trunk diameter and density. Distance between trees was measured with a tape measure, and gives the distance to the nearest tree for all trunks growing from (apparently) separate roots. Distances between trunks of the same tree were not measured. Canopy cover and understory cover for each quadrat were both assessed visually by the same observer in a qualitative scale from 1 to 5, where 1 was no or very little cover and 5 was very dense cover. Canopy cover was defined as the area of the quadrat surface covered by a vertical projection of aboveground vegetation, as in Jennings et al. (1999). Understorey was defined as the density of vegetation below breast height, giving an indication of structural complexity of the vegetation. Finally, all trees present in each quadrat were identified to species level and grouped for analysis by genera and by the type of tree (‘tree type’) to separate broadleaf (angiosperm) trees from coniferous (gymnosperm) trees, because breeding Willow Warblers largely forage on leaf-consuming caterpillars (Nilsson & Ebenman Citation1981). The abundance of each tree genera per quadrat was recorded to assess whether Willow Warbler males select trees of a particular genus when establishing territories.
Caterpillar abundance survey
To quantify food abundance, a caterpillar frass trap was set up under the largest broadleaf tree (or largest coniferous tree when no broadleaf trees were present) in each of 20 quadrats (10 ‘territory’ and 10 ‘non-territory’ quadrats, chosen at random). The largest tree was chosen to maximize the amount of frass sampled, because the quadrats were sufficiently small for Willow Warblers to forage on all trees within it. The traps were constructed using 50 × 50 cm cotton cloths suspended between tent pegs 30 cm above the ground, following the model of Fischbacher et al. (Citation1998). All traps were set up and taken down on the same days (running for 24 hours), during the peak time of breeding for Willow Warblers. The number of frass on the trap was then counted by the same person and used as an estimate of food availability.
Statistical analysis
Differences in habitat variables and food abundance between territory and non-territory quadrats were analysed for continuous variables using a two-sample t-test (trunk density, tree distance and food abundance), or alternatively a Mann–Whitney test in cases where distribution was not normal and variances were not homogenous, according to Levene's test for equality of variances (trunk diameter and abundance of each tree genera). Categorical variables were compared using a Likelihood ratio test and significance assessed by the Monte Carlo method to account for low expected frequencies and small sample sizes.
To assess the contribution of habitat variables in determining Willow Warbler territory occupancy we used generalized linear models with a binomial distribution and a log link function (using the ‘Stats’ and ‘AICcmodavg’ packages in R). We constructed a set of competing likelihood models by testing territory occupancy (0 for non-territory quadrats and 1 for territory quadrats) against each individual habitat variable (represented by linear and quadratic function), as well as all potential combinations of significant main effects. The significance of each habitat variable was tested by assessing changes in deviance between models with and without the individual term using chi-squared (χ2) tests. The Akaike Information Criterion for small samples (AICc) was used to rank and select the best model (the model with the lowest AICc value). All analyses were conducted using the R statistical software (R Core Development Team Citation2012).
RESULTS
Woodland structure
Trunk density per quadrat was significantly higher in territory areas than in non-territory areas (a, ). Average diameter of tree trunks did not significantly differ between territory and non-territory quadrats (), however, the variance of the data was not equal in the two categories (Levene's test, F = 25.5, P < 0.001). In territory areas, the diameters of trees tended to lie within a narrow range of 5–10 cm; in non-territory areas, the variation was large with upper and lower quartiles reaching up to 30 cm and below 3 cm (b). The average distance between trees did not significantly differ between territory and non-territory quadrats ().
Figure 1. Trunk density (a) and average trunk diameter (b) in quadrats within (Yes) and outside (No) of male Willow Warbler territories. The bold line inside the box denotes the median value, the box shows the inter-quartile range of the median and the whiskers denote the largest and smallest values of the data.
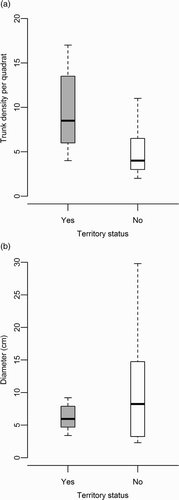
Table 1. Description of habitat variables considered in the study for quadrats within and outside Willow Warbler territories.
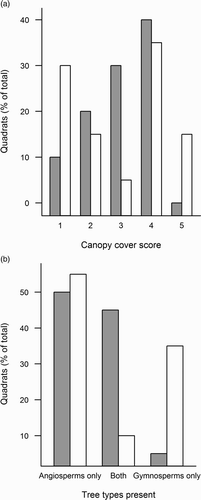
Canopy cover varied between territory and non-territory quadrats, with territory quadrats more likely to have intermediate canopy cover scores (a), however, the difference was marginal (Likelihood ratio = 10.4, df = 4, P = 0.056). No significant differences in understory cover were observed between territory and non-territory quadrats (Likelihood ratio = 8.1, df = 4, P = 0.18).
Figure 2. Proportion of quadrats within (grey bars) and outside (white bars) of Willow Warbler territories for (a) different canopy cover scores (in order of increasing cover) and (b) tree types present: quadrats containing either only broadleaf trees (Angiosperms), only coniferous trees (Gymnosperms) or both.
Woodland tree composition
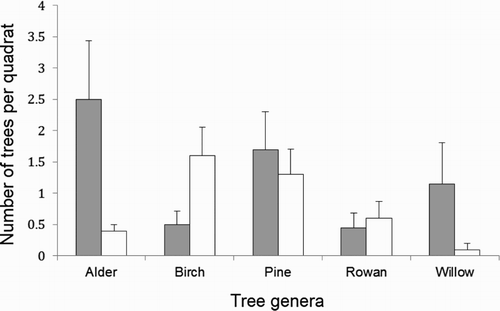
A total of 16 different species of trees were recorded, belonging to the following genera: Acer (Acer pseudoplatanus), Alnus (Alnus glutinosa, Alnus incana), Betula (Betula pendula), Crataegus (Crataegus monogyna), Fraxinus (Fraxinus excelsior), Larix (Larix decidua), Picea (Picea abies), Pinus (Pinus sylvestris), Populus (Populus tremula), Prunus (Prunus padus), Quercus (Quercus robur), Salix (Salix alba, Salix caprea, Salix cinerea) and Sorbus (Sorbus aucuparia). The majority of territory quadrats contained at least one broadleaf (Angiosperm) tree (b) and overall the distribution of quadrats among the tree type categories differed significantly from expected values (Likelihood ratio = 1.0, df = 2, P = 0.010). Tree genus abundance differed between territory and non-territory quadrats for two of the tree genera recorded (). Territory quadrats contained significantly more alder (Alnus sp.) and willow (Salix sp.) trees than non-territory quadrats ().
Figure 3. Abundance of different tree genera (for genera with abundance > 5 trees) within (grey bars) and outside (white bars) Willow Warbler territories. The mean and se are presented.
Caterpillar abundance
The average caterpillar frass counts did not significantly differ between territory and non-territory quadrats ().
Associations between habitat variables and territory occupancy
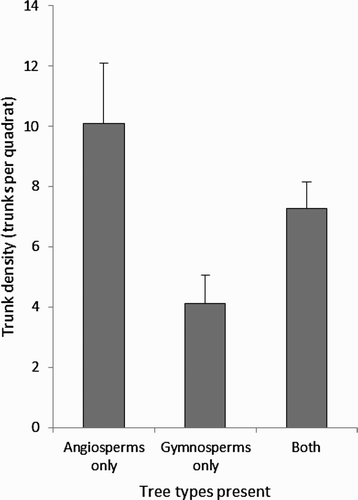
Trunk density was positively correlated with alder (r = 0.41, n = 40, P = 0.008) and willow abundance (r = 0.82, n = 40, P < 0.001). The number of alder trees in a quadrat was significantly different among canopy cover scores (Z = 11.3, n = 40, P = 0.024, ), quadrats with both intermediate and high canopy cover scores contained more alder trees. Note that there was no difference between total numbers of trees (i.e. trunk density) between canopy cover scores, due to the sampling area excluding areas not containing trees. Finally, trunk density was significantly lower in quadrats containing only coniferous (Gymnosperm) trees (Z = 7.7, n = 40, P = 0.021, ). No other significant correlations were observed between habitat variables.
Figure 4. Abundance of alder (Alnus sp.) trees in all quadrats which differ in their canopy cover scores (in order of increasing cover). The mean and se are presented.
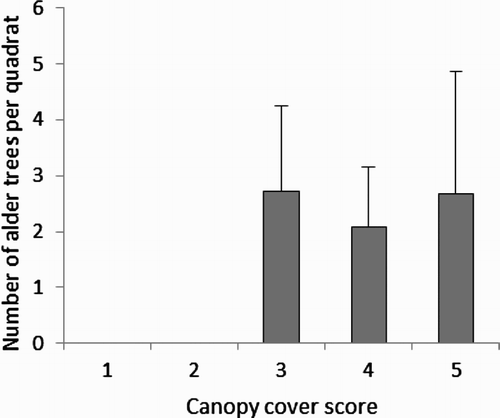
Figure 5. Trunk density in all quadrats containing only broadleaf trees (Angiosperms), only coniferous trees (Gymnosperms) or both. The mean and se are presented.
Territory occupancy by Willow Warbler males was significantly predicted by habitat variables related to both woodland structure and to woodland tree composition (). Trunk diameter was a significant factor when included as a quadratic term (), and was the best single factor in predicting territory occupancy (AICc = 39.7, lowest for any single factor model). Trunk density, canopy cover, tree type and the abundance of alder, willow and birch were all found to have a significant effect on territory occupancy, and were subsequently used for the two- and three-factor models ().
Table 2. Results of generalized linear models of Willow Warbler territory occupancy against habitat variables (Trunk density, Trunk diameter, Tree distance, Canopy cover, Understory cover, Tree type, Alder abundance, Willow abundance, Birch abundance, Pine abundance and Rowan abundance).
Models including habitat variables related to woodland structure (trunk density, trunk diameter and canopy cover) had lower AICc values compared to models including habitat variables related to woodland tree composition (abundance of alder, willow or birch). Territory occupancy was best predicted by a two-factor model incorporating trunk density and trunk diameter (), with a higher probability of territory occupancy for quadrats containing high density of trunks and trees with intermediate trunk diameter. Models including canopy cover and tree type were also good predictors of territory occupancy (difference in AICc with the best model less than 2, ).
DISCUSSION
Willow Warbler males in our study area had a specific habitat structure they selectively prefer to breed in. Territory areas contained higher numbers of trunks, had a narrow range of trunk diameters, and intermediate canopy cover.
Territory areas had significantly higher trunk numbers, and models including trunk density achieved the lowest AICc values, an indication that trunk density was a good predictor of territory occupancy. Willow Warbler presence has been associated with relatively young woodlands which typically contain high numbers of trees (Cramp 1992, Tappeiner et al. Citation1997, Amar et al. Citation2006). The preference for high tree density may be related to higher availability of foraging sites, as it has been found that Willow Warblers forage significantly more in the tree vegetation itself (leaf, twig and trunk) than in the air or on the ground (Salewski et al. Citation2003).
Tree type was a significant predictor of Willow Warbler habitat occupancy, with territory areas containing more broadleaf trees. This is consistent with previous literature (Tiainen et al. Citation1983, Felton et al. Citation2011). In areas with only coniferous trees, trunk number was lower than in areas with broadleaf trees or both tree types, which may have influenced the preference for broadleaf trees, because trunk number was a better predictor of territory occupancy than tree type. It is tempting to state that broadleaf trees were preferred because they are a better host for most caterpillars, i.e. providing the Willow Warblers with increased food, but we found no significant differences in caterpillar frass numbers between territory and non-territory areas, and caterpillar density was a poor predictor of territory occupancy. However, our method measured only one part of the Willow Warbler diet (Jones et al. Citation2010). In addition, the catching time of our traps was limited to 24 hours due to poor weather conditions, and therefore may have underestimated food abundance.
Average trunk diameter did not differ between territory and non-territory areas, but territory occupancy was best predicted by the quadratic term of trunk diameter, indicating that Willow Warblers prefer a range of diameters which is quite well-defined, avoiding areas with very large or very small trees. This is consistent with previous literature; for example, Bellamy et al. (Citation2009) narrowed the preferred tree height in Willow Warbler territories down to a range of only 1.6 m of variation. Similarly, Gillings et al. (Citation1998) reported that males avoided trees that were smaller or larger than 1–4 m in height.
Willow Warblers appeared to also have a preference for certain species of trees. Willows (Salix sp.) were common in the territories, as were alder (Alnus sp.) trees. Interestingly, however, there was no preference for birch (Betula sp.) trees, despite birch being commonly reported in the literature as one of the most used tree species for Willow Warbler habitats (Yapp Citation1962, Mason Citation1997, Gillings et al. Citation1998, Felton et al. Citation2011). At the study site, this preference was not observed. Indeed, there appeared to be a tendency to avoid birch trees with 75% of birches growing in non-territory areas. Willow and alder trees had higher trunk number than the birch trees in our study area, however, trunk number was a better predictor of territory occupancy than the abundance of either alder, willow or birch trees. This indicates that the birds prefer areas with higher trunk densities, regardless of tree identity, and as a consequence, they tend to be associated with tree species which have high trunk numbers. Birches are often found as young re-growth (Gimingham Citation1984), which provide ideal Willow Warbler habitat. However, at our study site, birch trees were generally found in the older, undisturbed parts of the nature reserve, and thus were large and had few trunks, therefore being unsuitable for Willow Warbler territories. Thus, Willow Warblers might at first be observed to select breeding areas with certain tree species, whereas in fact the selection criterion is not the tree species in itself, but instead is the structure of the tree. Indeed, Bellamy et al. (Citation2009) mentions that the observed preference for birch may be confused with structural differences. Our results indicate that these suspicions are likely well founded, as habitat variables associated with woodland structure were better predictors of Willow Warbler territory (lower AICc values) than the abundance of any particular tree genera.
A tendency was observed for territories to cover areas with intermediate canopy cover: models including canopy cover had AICc values similar to the best predictive model. This is in slight contrast to the results presented by Amar et al. (Citation2006) who found a significant negative correlation between canopy cover and Willow Warbler abundance. The preference for some canopy cover may relate to the need for protecting the nest from predators and exposure, as these factors have been observed to impact fledgling rates of the open, ground-nesting Willow Warblers (Bjørnstad & Lifjeld Citation1996).
It appears that the preferred habitat type of the Willow Warblers in WNR resembles that of young and coppiced woodland – high trunk density and a fairly small trunk size, with intermediate canopy cover. This result is important because it suggests that the decline of the Willow Warbler populations in southern England could be linked to habitat deterioration because there has been a decline in coppice management in this area (Natural England Citation2009). Mason (2007) report that after a long-term decline, traditional coppice management ceased almost completely after the 1950s. Coppicing also declined in the north of England throughout the 20th century, but coppice has historically been more widespread in the south (Fuller & Warren Citation1993) and thus the degradation of these habitats might have relatively more impact in the south. In addition, changes to forestry policy have led to a decline in young growth in broadleaf forests, particularly due to the manifestation of continuous cover forestry which avoids clear-felling (Mason 2007). The combination of loss of coppice management and forests dominated by older trees has meant that there is now very little re-growth of young trees to fit the Willow Warbler's habitat requirements. Therefore, due to the transient nature of these ecosystems, active management and planting of new areas would be essential to prevent further habitat deterioration (Bellamy et al. Citation2009).
In conclusion, our results indicate that Willow Warbler males choose their territories on the basis of a specific woodland structure. The woodland structure found here is largely consistent with earlier research. However, this study indicates that the identity of the tree species may not be a selection factor in itself, in contrast to several earlier studies. Mason (Citation1997) suggested that as the bird preferred birch, and there had been no decline in the birch habitat in the south of England, the Willow Warbler decline was unlikely to be due to reduced breeding habitat. Our results do not support this conclusion, and instead suggest the opposite: habitat deterioration may have played a substantial role in the decline of Willow Warblers due to a reduction in coppice management and increasing age of British forests. However, this apparent relationship between forestry trends and Willow Warbler decline is merely an observed pattern, in a limited area, and no causation is proven, providing a potential focus for future investigations.
ACKNOWLEDGEMENTS
We would like to thank Watchtree Limited for providing the opportunity to work in their nature reserve, especially Frank Mawby, Dr Liz Still and Tim Lawrence for their assistance, and for providing useful data. We also thank Dr Ian Hartley for ornithological advice, David Rogers for assistance with field data collection and Mark Mainwaring, Will Cresswell, Simon Gillings and an anonymous referee for invaluable comments on the manuscript.
REFERENCES
- Amar, A., Hewson, C.M., Thewlis, R.M., Smith, K.W., Fuller, R.J., Lindsell, J.A., Conway, G., Butler, S. & MacDonald, M.A. 2006. What's happening to our woodland birds? Long-term changes in the populations of woodland birds. BTO Research Report 169. BTO, Thetford.
- Arvidsson, B. & Klaesson, P. 1986. Territory size in a Willow warbler Phylloscopus trochilus population in Mountain Birch Forest in Swedish Lapland. Ornis Scand 17: 24–30. doi: 10.2307/3676749
- Bellamy, P.E., Hill, R.A., Rothery, P., Hinsley, S.A., Fuller, R.J. & Broughton, R.K. 2009. Willow warbler Phylloscopus trochilus habitat in woods with different structure and management in southern England. Bird Study 56: 338–348. doi: 10.1080/00063650902806914
- Bjørnstad, G. & Lifjeld, J.T. 1996. Male parental care promotes early fledging in an open-nester, the Willow Warbler Phylloscopus trochilus. Ibis 138: 229–235. doi: 10.1111/j.1474-919X.1996.tb04333.x
- Cramp, S. & Brooks, J.D. (eds) 1992. Birds of the Western Palearctic, Vol. 6. Oxford University Press, Oxford.
- Felton, A., Andersson, E., Ventorp, D. & Lindbladh, M. 2011. A comparison of avian diversity in spruce monocultures and spruce–birch polycultures in southern Sweden. Silva Fennica 45: 1143–1150. doi: 10.14214/sf.92
- Fischbacher, M., Naef-Daenzer, B. & Naef-Daenzer, L. 1998. Estimating caterpillar density on trees by collection of frass droppings. Ardea 86: 121–129.
- Fuller, R.J. 2002. Spatial differences in habitat selection and occupancy by woodland bird species in Europe: a neglected aspect of bird–habitat relationships. In Chamberlain, D. & Wilson, A. (eds) Avian Landscape Ecology: Pure and Applied Issues in the Large-scale Ecology of Birds. Proceedings of the 11th Annual IALE (UK) Conference: 101–111. IALE (UK), Norwich.
- Fuller, R.J. & Henderson, A.C.B. 1992. Distribution of breeding songbirds in Bradfield Woods, Suffolk, in relation to vegetation and coppice management. Bird Study 39: 73–88. doi: 10.1080/00063659209477103
- Fuller, R.J. & Warren, M.S. 1993. Coppiced Woodlands: Their Management for Wildlife, 2nd edn. Joint Nature Conservation Committee, Peterborough.
- Fuller, R.J., Noble, D.G., Smith, K.W. & Vanhinsbergh, D. 2005. Recent declines in populations of woodland birds in Britain: a review of possible causes. British Birds 98: 116–143.
- Gillings, S., Fuller, R.J. & Henderson, A.C.B. 1998. Avian community composition and patterns of bird distribution within birch-heath mosaics in north-east Scotland. Ornis Fennica 75: 27–37.
- Gimingham, C.H. 1984. Ecological aspects of birch. Proc. R. Soc. Edinburgh Sect. B 85: 65–72.
- Holden, P. & Cleeves, T. 2010. RSPB Handbook of British Birds, 3rd edn. Christopher Helm, London.
- Jennings, S.B., Brown, N.D. & Sheil, D. 1999. Assessing forest canopies and understorey illumination: canopy closure, canopy cover and other measures. Forestry 72: 59–74.
- Jones, P., Salewski, V., Vickery, J. & Mapaure, I. 2010. Habitat use and densities of co-existing migrant Willow warblers Phylloscopus trochilus and resident eremomelas Eremomela spp. in Zimbabwe. Bird Study 57: 44–55. doi: 10.1080/00063650903071401
- Marchant, J.H. 1983. BTO Common Bird Census Instructions. British Trust for Ornithology, Tring.
- Mason, C.F. 1997. Association between Willow Warbler Phylloscopus trochilus territories and birch in woodlands in southeastern England. Ibis 139: 411–412. doi: 10.1111/j.1474-919X.1997.tb04648.x
- Mason, W.L. 2007. Changes in the management of British forests between 1945 and 2000 and possible future trends. Ibis 149: 41–52.
- Morrison, C.A., Robinson, R.A., Clark, J.A. & Gill, J.A. 2010. Spatial and temporal variation in population trends in a long-distance migratory bird. Diversity. Distrib. 16: 620–627. doi: 10.1111/j.1472-4642.2010.00663.x
- Natural England. 2009. South West Future Landscapes Programme: Phase 1 Technical Report. Available from: http://www.naturalengland.org.uk
- Nilsson, S.G. & Ebenman, B. 1981. Density changes and niche differences in Island and Mainland Willow warblers Phylloscopus trochilus at a Lake in Southern Sweden. Ornis Scand 12: 62–67. doi: 10.2307/3675906
- R Core Development Team. 2012. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. Available from: http://www.R-project.org/
- Risely, K., Noble, D.G. & Baillie, S.R. 2008. The breeding bird survey 2007. BTO Research Report 508. BTO, Thetford.
- Salewski, V., Bairlein, F. & Leisler, B. 2003. Niche partitioning of two Palearctic passerine migrants with Afrotropical residents in their West African winter quarters. Behav. Ecol. 14: 493–502. doi: 10.1093/beheco/arg021
- Still, L. 2011. Watchtree nature reserve ornithological monitoring 2011. Unpublished report.
- Still, L. 2012. Watchtree nature reserve ornithological monitoring 2012. Unpublished report.
- Tappeiner, J.C., Huffman, D.W., Marshall, D., Spies, T.A. & Bailey, J.D. 1997. Density, ages, and growth rates in old-growth and young-growth forests in coastal Oregon. Can. J. For. Res 27: 638–648. doi: 10.1139/x97-015
- Tiainen, J., Wickholm, M., Pakkala, T., Piiroinen, J. & Virolainen, E. 1983. The habitat and spatial relations of breeding Phylloscopus warblers and the goldcrest Regulus Regulus in southern Finland. Annales Zoologici Fennici 20: 1–12.
- Yapp, W.B. 1962. Birds and Woods. Oxford University Press, Oxford.