
Abstract
Capsule Bird and seed abundances decreased in both standard and enhanced Agri-Environment Scheme seed mixtures (plots of seed-bearing crops) between November and February.
Aims We analysed effects of Agri-Environment seed mixture type (‘standard’ or ‘enhanced’), plot age and non-target species on seed and bird abundance.
Methods Surveys were conducted in November/December and January/February on 36 standard and 19 enhanced seed mixtures to test effect of plot type and age on abundance of buntings, finches and seeds.
Results Bunting and finch densities decreased between January/February and November/December. Seed decreased by approximately 70% between surveys, plot type did not affect depletion. Second-year plots contained nearly 90% less seed, and non-significantly lower bunting and finch densities. Bunting densities were positively related to cereal seed abundance, and finch densities to non-cereal seed; these relationships did not differ between plot types. There was no evidence that non-target species affected use of seed mixtures by target species.
Conclusion There was no evidence that plot type affected seed or bird abundances. Increased quality and uptake of Agri-Environment Scheme seed-rich habitats (seed mixtures and over-winter stubbles, possibly alongside well-implemented supplementary feeding) is needed to provide late-winter seed, especially cereals for species of conservation concern.
Lack of seed food in winter is one of the key causes of farmland bird declines in the UK and across Europe, alongside loss of nest sites, and insect food in summer (Wilson et al. Citation2009). Low over-winter survival rates explain the decline of a suite of granivorous farmland birds, including Yellowhammer Emberiza citrinella, Reed Bunting Emberiza shoeniclus and Chaffinch Fringilla coelebs (Peach et al. Citation1999, Siriwardena et al. Citation1999, Citation2000). Availability of seed food has decreased on farmland in England for a number of reasons, such as increased herbicide use, fewer over-winter stubbles, more efficient crop harvesting and storage and, more recently, the end of the set-aside policy in 2008 (Siriwardena et al. Citation2008, Morris et al. Citation2011). Seed food is particularly scarce in late winter (Siriwardena et al. Citation2008).
Two main types of options that increase seed abundance are widely available in Agri-Environment Schemes in the UK: seed mixtures and over-winter stubbles. In addition to these, a supplementary feeding option has recently been added. Seed mixtures involve cultivation of seed-bearing crops for one or two years, with restricted pesticide applications (Stoate et al. Citation2004). These can be rotational or non-rotational (Natural England Citation2013a). Seed mixtures are associated with increased bird abundances in the seed mixture plots (Stoate et al. Citation2003, Perkins et al. Citation2008, Field et al. Citation2011), with some evidence of benefits at the farm (Stoate et al. Citation2003) and landscape (Baker et al., Citation2012) scale. More generally, Agri-Environment Scheme options that provide winter seed (seed mixtures and over-winter stubbles) can have a positive effect on population growth rates of some species of granivorous farmland bird at the landscape scale (Baker et al. Citation2012).
Around 70% of farmland in England is currently managed under an Agri-Environment Scheme (Natural England Citation2013a); the main ones being Entry Level Stewardship, a ‘broad and shallow’ scheme open to all farmers, and Higher Level Stewardship, a targeted scheme with competitive entry between farmers. Two seed mixture options are available under these schemes, a standard seed mixture (‘EF2’) under Entry Level Stewardship and an enhanced seed mixture (‘HF12’) under Higher Level Stewardship. EF2 involves sowing at least three of the following crops: barley, triticale, kale, quinoa, linseed, millet, mustard, fodder radish and sunflower (Natural England Citation2013b), whilst HF12 is tailored to the bird species present (Natural England Citation2013c), for example, an annual mix for granivores consisting mostly of cereals and millet, an annual mix for tree sparrow Passer montanus containing 40% millet, or a two-year mix for grey partridge Perdix perdix containing 40% kale (P. Grice pers. comm.). Approximately 18% of Agri-Environment Scheme agreements in England contain seed mixture options, covering just under 0.2% of the option area (P. Grice, pers. comm.).
Standard EF2 and enhanced HF12 seed mixture plots differ in that there is no requirement for standard plots to contain cereals, despite the importance of cereal seed in the diet of many farmland birds of conservation concern, for example, buntings such as Yellowhammers and Reed Buntings (Wilson et al. Citation1996). As a result, Field et al. (Citation2011) found that enhanced plots contained more cereals than standard plots, and that there was a non-significant trend for enhanced plots to contain more Yellowhammers, Corn Buntings Emberiza calandra and Tree Sparrows, and standard plots to contain more Carduelis finches, most of which are of lesser conservation concern. Seed mixture plots are either sown annually or every two years and this is another factor affecting abundance of cereal seed in plots, as two-year plots will lack the annual cereal component in their second year (excluding the small amount due to self-sown cereal ‘volunteers’ from the previous crop, Perkins et al. Citation2008). This creates a particular problem if two-year plots are established simultaneously across a farm when no plots contain cereals in the second year. Perkins et al. (Citation2008) found that plot age affected bird use with buntings, Tree Sparrows and Skylarks Alauda arvensis more likely to be found in first-year plots, but occurrence of finch species either unrelated to plot age, or more likely in second-year plots.
It has been suggested that the effectiveness of seed mixtures in benefiting target species may be limited by competition or predation from other species (Pywell et al. Citation2007, Perkins et al. Citation2008, Baker et al. Citation2012). Direct competition by dominant species of bird may limit use of plots by smaller granivorous passerines (Krams Citation2001), non-target bird species such as corvids, columbids and gamebirds (except grey partridge), could also limit use of seed mixture plots by indirect competition, i.e. depletion of seed within plots (Pywell et al. Citation2007, Perkins et al. Citation2008), and concentrations of birds in plots could lead to increased predation pressure (Bro et al. Citation2004, Baker et al. Citation2012).
In the present study, data from bird, seed and vegetation surveys in standard EF2 seed mixture plots on Entry Level Stewardship farms, EF2 plot equivalents on Higher Level Stewardship farms (‘HF2’ to investigate farm context of EF2) and enhanced HF12 seed mixture plots during early and late winter of 2010/2011 were used to test: (1) whether type or age of seed mixture plot affected their use by buntings or finches, and whether this was affected by the presence of non-target bird species; (2) whether vegetation structure, composition and abundance or retention of seeds differed between types or ages of seed mixture plots; (3) whether these differences in vegetation explained differences in use by buntings and finches (4) whether non-target bird species significantly depleted seed resources available to buntings or finches.
METHODS
Surveys were conducted at ten EF2 plots, eight HF2 plots and ten HF12 plots centring on East Anglia an arable-dominated landscape in eastern England, and at nine EF2, nine HF2 and nine HF12 plots centring on Oxfordshire a mixed arable/pastoral landscape in central England (). These regions were identified as having relatively high rates of uptake of the different seed mixture options. Where possible, first-year plots were selected so that plot types of similar ages could be compared, but in East Anglia, eight EF2, four HF2 and four HF12 plots were surveyed in their second-year due to low availability of first-year plots (this was controlled for in analyses). Survey plots were selected using a GIS database of Agri-Environment Scheme options, on farms that had been under Agri-Environment Scheme agreements for at least two years. One of each of the three plot types were selected as close as possible to each other (usually within 10 km), but at a minimum distance of 2 km, to increase the likelihood of the same species assemblages being present, whilst trying to maintain sampling independence among individual birds. Only one plot of each option type was surveyed on any individual farm (with one exception), to ensure that replicates were independent in terms of location and management. Plots of different option types were also surveyed on the same farm in one case. Seed mixture plots that were re-sown in the autumn prior to surveys were not surveyed.
Figure 1. Locations of seed mixture plots surveyed.
Each plot was surveyed twice; once in November/December 2010 (‘early winter’), and once in January/February 2011 (‘late winter’). Bird surveys were not conducted in the first hour after dawn or late in the evening, to avoid including birds that were travelling to or from roost sites (Stoate et al. Citation2003). Birds seen or heard using the plot were recorded during a one-hour watch from a car or portable hide. To reduce effects of disturbance, surveys did not start until 15 minutes after entering the hide. Birds were recorded that were in the plot on arrival, and during a diagonal transect walked through the plot at the end of the survey. The maximum number of birds in the plot at any one time was then calculated for each species to avoid double counting. If plots were too long to be viewed from a single vantage point then surveys were either conducted in two parts from different vantage points, or a subsample of the plot marked by canes was surveyed, in which case the area of this subsample was used in analysis. Effect of disturbance from predators was controlled for by noting presence of any raptor or corvid avian predators in, adjacent to, or flying over the plot during bird surveys. Surveys were not conducted in persistent rain, winds over Beaufort force 5, heavy fog or snow cover.
Vegetation surveys were conducted at each visit. Ten 0.5 m × 0.5 m vegetation quadrats were spaced at equal distances along a diagonal transect of the plot, a minimum of 2 m from the plot edge. Percentage cover of sown cereals, sown non-cereals and bare ground, and vegetation height at the four corners of the quadrat were noted. Seed-heads were taken from the top left 20 cm × 20 cm of each quadrat. Seeds were later separated into sown cereals and sown non-cereals and dried at 45°C for 48 hours to stabilize mass before weighing. Mean values from the ten quadrats were used in analyses. Seed samples were also taken from the upper 5 mm of soil, but amounts of seed in these were too small to include in analyses (also found by Gruar et al. Citation2013).
Statistical analysis
Analyses were conducted in SAS v 9.1 (SAS Institute Citation2003). REGION (East Anglia/Oxfordshire) was controlled for in all models. Results presented are for full models. Significant factor effects were investigated by comparing predicted means (using the LS Means function in SAS, Littell et al. Citation1996).
Effect of plot type and age on bird densities
Bird counts were modelled using generalized linear mixed models (GLMMs), with Poisson distribution and logarithmic link function, PLOT as a random effect to control for repeat surveys, and the log of plot area (or survey area) used as an offset so that densities were being modelled. Overdispersion was accounted for by multiplying variances by the observed overdispersion parameter (using the random ‘residual’ term in PROC GLIMMIX of SAS, Littell et al. Citation1996). If models did not converge with this adjustment, a unique survey identifier was specified as a random effect (equivalent to assuming a Poisson lognormal distribution). Combined groups of ‘buntings’ (Yellowhammer and Reed Bunting; Corn Bunting were not included in this category because they were only recorded in 8/110 surveys), and ‘finches’ (Chaffinch, Linnet, Greenfinch Carduelis chloris and Goldfinch Carduelis carduelis) were used in analyses. Too few of the granivorous species specifically targeted by Higher Level Stewardship (Corn Bunting, Tree Sparrow and Grey Partridge) were recorded to include them in analyses.
Bunting or finch counts were specified as dependent variables, and modelled in relation to the fixed effects REGION (East Anglia and Oxfordshire), SURVEY (1 or 2), PLOT TYPE (EF2, HF2 or HF12), PLOT AGE (first- or second-year), presence of predators (presence or absence of any avian predator species during the survey) and aggregate count of non-target species, along with all two-way interactions between SURVEY, PLOT TYPE and PLOT AGE. The ‘non-target species’ category was defined as potential corvid, gamebird and columbid competitor species, and included Jackdaw Corvus monedula, Magpie Pica pica, Rook Corvus frugilegus, Pheasant Phasianus colchicus, Red-legged Partridge Alectoris rufa and Woodpigeon Columba palumbus (no Carrion Crows Corvus corone or Collared Doves Streptopelia decaocto were recorded in plots). Predators were recorded during 36% of surveys (n = 110) and non-target species during 40% (n = 110). The range of aggregate count of non-target species was 0–100 (median = 0; median if only the 44 surveys where non-target species were present are considered = 5).
Effect of plot type and age on vegetation and seed
General linear models (GLM) with normal distribution were run using PROC GENMOD in SAS with vegetation height, % cover of bare ground, % cover of sown cereals, % cover of sown non-cereals, and mass of cereal seed and non-cereal seed included as dependent variables. PLOT TYPE and PLOT AGE were included as fixed effects. Values of vegetation height and percentage cover of different vegetation categories were not expected to change much between surveys, and so these models were run for survey 1 only, to avoid pseudoreplication by including two very similar values for each seed mixture. For models of seed mass, whether plot type or age affected rate of depletion between surveys was of interest, and so mixed models with normal distribution were run using PROC GENMOD, with PLOT as a random effect, SURVEY was included as a fixed effect, along with all two-way interactions between SURVEY, PLOT TYPE and PLOT AGE. Seed masses were log transformed to improve model fit.
Effect of vegetation and seed on bird densities
GLMMs were run with bunting or finch counts as dependent variables, with vegetation height, % cover of bare ground, mass of cereal and non-cereal seed as fixed effects.
Effect of non-target species on seed depletion
GLMs were run with the mass of cereal and non-cereal seed in late winter as the dependent variable. The density of the aggregate number of non-target species in early winter and PLOT TYPE were included as fixed effects plus their interaction term, to investigate whether plots with higher densities of non-target species in early winter contained less seed by late winter, and whether this depended on type of plot. Plot age was not included because only two of the samples from second-year plots contained sown cereal seed and only four contained sown non-cereal seed. Seed masses were log transformed to improve model fit.
RESULTS
Effect of plot type and age on bird densities
Densities of buntings and finches in seed mixture plots were higher in early winter than late winter (, ) and there was a non-significant trend for densities to be higher in first-year than second-year plots (). Densities of buntings and finches were unrelated to plot type, the presence of predators during surveys or the intensity of use by non-target species ().
Figure 2. Mean predicted densities (+/− se) of buntings and finches in seed mixture plots in relation to survey.
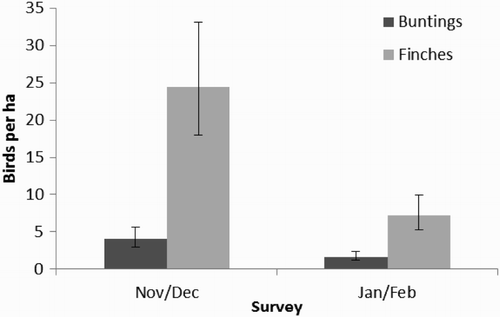
Table 1. Results of GLMM analyses relating bunting and finch densities to features of seed mixture plots recorded during surveys. Significant terms highlighted in bold.
Effect of plot type and age on vegetation structure and seed
Seed became depleted in plots between early and late winter (, ), but mass of seed (cereal and non-cereal), and rate of seed depletion did not differ between plot types (). Although standard seed mixtures are not required to contain a cereal component, which may affect the bird species using them, information provided by 43 farmers on mixtures sown showed that all except one (an EF2 plot) contained a cereal component (information was provided for 14/19 EF2 plots, 16/19 HF2 plots and 13/17 HF12 plots). Second-year plots contained less bare ground and lower cover of sown cereal and non-cereal plants than first-year plots, and less cereal and non-cereal seed (although this was of borderline significance for non-cereal seed, P = 0.068, , ).
Figure 3. Mean predicted densities of cereal and non-cereal seed samples from seed mixture plots in relation to (a) survey and (b) plot age.
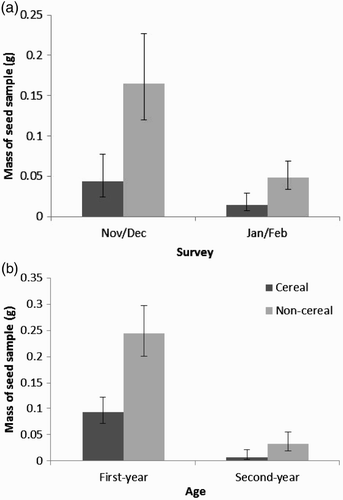
Table 2. Results of GLMM analyses relating vegetation attributes in relation to features of seed mixture plots. Significant terms highlighted in bold.
Effect of vegetation and seed on bird densities
Bunting densities were positively related to mass of cereal seed and negatively related to vegetation height (). Finch densities were positively related to mass of non-cereal seed ().
Table 3. Results of GLMM analyses relating vegetation attributes of seed mixture plots to densities of buntings and finches. Estimates shown on modelled log scale. Significant terms highlighted in bold.
Effect of non-target species on seed depletion
There was no relationship between the density of non-target species during early winter and the mass of seed in seed mixture plots in late winter ().
Table 4. Results of GLM analyses relating mass of seed in January/February 2011 to numbers of buntings, finches and non-target species in November/December 2010. Significant terms highlighted in bold.
DISCUSSION
Bunting densities were positively associated with cereal seed, and finch densities with non-cereal seed, and both types of seed became depleted by late winter (by 75% for seed of sown cereals and 69% for seed of sown non-cereals) as found by previous studies (Stoate et al. Citation2003, Citation2004, Perkins et al. Citation2008, Siriwardena et al. Citation2008). There was no evidence that high densities of non-target birds in the early winter led to this depletion, or that direct competition by non-target species had a negative effect on use of plots by buntings or finches, despite the study being conducted during a relatively cold winter with more frequent snow cover than usual. Other non-target species, such as mammals and insects, are likely to have had an effect on depletion but were not investigated in this study. There was no evidence that enhanced seed mixtures were used more, or retained more seed into late winter, than their standard equivalents. Bunting densities were higher in plots with lower vegetation height, this is likely to reflect greater accessibility of seed resources.
Second-year plots contained 89% less sown cereal seed than first-year plots, 88% less seed of sown non-cereals, less than a quarter of the cover of sown species and less than half the amount of bare ground, which is likely to affect accessibility of seed (Moorcroft et al. Citation2002). There was a trend for bunting and finch densities to be lower in second-year plot than first-year plots, but this was not significant despite the much lower amount of seed in second-year plots, possibly due to the low sample size for second-year plots. It could also be due to higher availability of weed seeds in second-year plots, Henderson et al. (Citation2004) found some evidence of higher bird densities in weedy game crops for some species. The lower cover of sown species in second-year plots suggests problems with establishment of biennial brassicas such as kale, which has been shown to attract more birds than most other seed mixture crops, particularly in late winter due to better seed retention. Most studies suggest that a mixture of kale, quinoa and cereals is most likely to attract a range of farmland birds and that establishment and seed production in kale can be enhanced by the application of low rates of nitrogen fertilizer (Stoate et al. Citation2003, Citation2004, Henderson et al. Citation2004). The factors affecting the poor establishment second-year plots noted in this study could reflect insufficient nitrogen fertilizer application and may warrant further investigation.
The low sample size of second-year plots was due to the fact that we had set out to investigate effects of seed mixture plot type (standard or ‘enhanced’ seed mixtures) rather than age; second-year plots were added in East Anglia due to low availability of first-year plots on study sites. Perkins et al. (Citation2008) found that buntings, Tree Sparrows and Skylarks A. arvensis were more likely to be found in first-year than second-year plots, but occurrence of finches was either unrelated to plot age, or more likely in second-year plots, depending on species. These results were attributed to cereal abundance. The cereal-feeding buntings and sparrows predominantly relied on first-year plots, whilst finches fed on oil seed rape and mustard in first-year plots and kale in second-year plots.
The lack of effect of plot type on seed abundance or bird use is in contrast to Field et al.’s (Citation2011) findings that standard EF2 plots contained less cereals, and tended to contain more Carduelis finches and Reed Buntings, with enhanced plots being used more by Yellowhammers, Corn Buntings and Tree Sparrows. Although we also found positive relationships between bunting densities and abundance of cereal seed, and finch densities and abundance of non-cereal seed, the amount of these did not differ between different plot types. This may reflect improved advice or better mixes available to Entry Level Stewardship farmers increasing the prevalence of cereals in EF2 plots, although as Entry Level Stewardship is designed to operate with little or no advisory support this is perhaps more likely that sample sizes were not large enough to detect an effect. This follows a recent study showing that type of Agri-Environment over-winter stubble did not affect bird densities (Bright et al. Citation2014), although the extended and low-input crop over-winter stubble options investigated were designed primarily to provide spring/summer foraging habitats, and nesting habitats for in-field nesters.
Amount of seed and its spatial arrangement in the landscape is another factor that will affect bird use of seed mixture plots and potentially mask effects of plot type or age on use by birds; resources did not allow habitat data to be collected at a sufficient spatial scale to investigate this.
Seed-rich habitats such as seed mixtures are a critical part of Agri-Environment Schemes, and can lead to more positive population growth for granivorous farmland birds at the landscape scale (Baker et al. Citation2012). Their ability to lead to more positive population growth at the national scale is limited by current low uptake of these options (Baker et al. Citation2012). Another limiting factor is the lack of provision of seed into the late-winter/early spring, sometimes termed the ‘hungry gap’ (Siriwardena et al. Citation2008), as highlighted by this study. Current farm management, including Agri-Environment Schemes, are unlikely to provide enough seed food during this time (Siriwardena et al. Citation2008); most Agri-Environment Scheme over-winter stubbles can be ploughed from mid-February. Standard seed mixture plots have to be retained until 1 March, by which time the seed is likely to be depleted, and are then only ploughed if they are being replaced with new mixtures, their position rotated around the farm or the Agri-Environment Scheme agreement has expired.
Given their importance for farmland bird populations, higher uptake of seed-rich habitats in Agri-Environment Schemes should be encouraged, to address the late winter ‘hungry-gap’ (Siriwardena et al. Citation2008). The inclusion in seed mixtures of crops that attract high densities of birds and retain some seed into late winter is important. For buntings, cereals are favoured, with triticale being particularly useful for grain retention. Kale is used by a wide range of species and also has good late winter seed retention (Stoate et al. Citation2004, Perkins et al. Citation2008). Fodder beet and fodder radish, the latter of which was commonly grown in the seed mixtures in this study, also retains seed well, although access to the seed within radish pods may be difficult for species with small, less powerful bills (Hinsley et al. Citation2010). However, with up to 90% seed depletion by mid-January (Hinsley et al. Citation2010) in some cases seed mixtures may need to be deployed in combination with well-implemented supplementary feeding. From 2013, an option in the Environmental Stewardship Scheme in England has enabled supplementation of food by the spreading a mixture of seeds (cereals, oilseed rape and canary grass) close to existing areas of over-wintered stubbles or Wild Bird Seed Mixtures at least once a week from 1 January until 31 March. Care must be taken to minimize the impact of vermin and disease transmission when spreading supplementary seed (Natural England Citation2013b, Stockdale et al. Citationin press).
There was no evidence that enhanced plots contained more seed or were used more than standard plots, although the extra cost associated with them is relatively low and they do allow for more control over targeting of seed mixtures, which may confer site-specific benefits for granivorous species specifically targeted by Higher Level Stewardship that were too scarce for this study to analyse. Improved quality and uptake of existing Agri-Environment Scheme options is needed to provide seed into late winter, especially cereal seeds needed by species of high conservation concern. Recent work has highlighted the potential value of seeded ryegrass as a source of late winter seed for farmland birds (Buckingham et al. Citation2011) and this option has been included as a new option in the English Environmental Stewardship Scheme, along with late winter supplementary feeding.
ACKNOWLEDGEMENTS
This work was jointly funded by the RSPB and Natural England through the Action for Birds in England (AfBIE) partnership. Surveys were conducted by JF, Chris De Ruyck and Alice Davey. Andrew Bladon, Alexander Rowell and Rosie Lennon provided invaluable assistance with data extraction. Many farmers and land-owners allowed us access to their land, and provided us with useful information.
REFERENCES
- Baker, D.J., Freeman, S.N., Grice, P.G. & Siriwardena, G.S. 2012. Landscape-scale responses of birds to agri-environment management: a test of the English Environmental Stewardship scheme. J. Appl. Ecol. 49: 871–882.
- Bright, J.A., Field, R.H., Morris, A.J., Cooke, A.I., Fern, J., Grice, P.V. & Peach, W. 2014. Influence of crop type and agri-environment management on the use of over-winter stubbles by farmland birds in England. Bird Study 61: 332–339.
- Bro, E., Mayot, P., Corda, E. & Reitz, F. 2004. Impact of habitat management on grey partridge populations: assessing wildlife cover using a multisite BACI experiment. J. Appl. Ecol. 41: 846–857.
- Buckingham, D.L., Bentley, S., Dodd, S. & Peach, W.J. 2011. Seeded ryegrass swards allow granivorous birds to winter in agriculturally improved grassland landscapes. Agr. Ecosyst. Environ. 142: 256–265.
- Field, R.H., Morris, A.J., Grice, P.V. & Cooke, A. 2011. The provision of winter bird food by the English Environmental Stewardship scheme. Ibis 153: 14–26.
- Gruar, D.J., Morris, A.J. & Dillon, I.A. 2013. Evaluating the efficacy of winter seed provision by different agri-environment scheme options. Asp. Appl. Biol. 118: Environmental Management on Farmland, 259–264.
- Henderson, I.G., Vickery, J.A. & Carter, N. 2004. The use of winter bird crops by farmland birds in lowland England. Biol. Conserv. 118: 21–32.
- Hinsley, S.A., Novakowski, M., Heard, M., Bellamy, P.E., Broughton, R.K., Hulmes, S., Hulmes, L., Peyton, J. & Pywell, R.F. 2010. Performance and effectiveness of winter bird food patches established under Environmental Stewardship: Results from the Hillesden experiment. Aspects of Applied Biology 100, Agri-environment schemes – what have they achieved and where do we go from here?, pp. 151–158.
- Krams, I.I. 2001. Seeing without being seen: a removal experiment with mixed flocks of Willow and Crested tits Parus montanus and cristatus. Ibis 143: 476–481.
- Littell, R.C., Milliken, G.A., Stoup, W.W. & Wolfinger, R.D. 1996. SAS System for Mixed Models. SAS Institute Inc., Cary, NC.
- Moorcroft, D., Whittingham, M.J., Bradbury, R.B., & Wilson, J.D. 2002. The selection of stubble fields by wintering granivorous birds reflects vegetation cover and food abundance. J. Appl. Ecol. 39: 535–547.
- Morris, A.J., Baldi, A., Hegarty, J. & Robijns, T. 2011. Setting aside farmland in Europe: the wider context. Agr. Ecosyst. Environ. 143: 1–2.
- Natural England. 2013a. LM update October 2013. Available to download from: http://www.naturalengland.org.uk/Images/lmupdate-13_tcm6-36807.pdf (Accessed 5 February 2014).
- Natural England. 2013b. Entry Level Stewardship. Environmental Stewardship Handbook, 4th edn. Natural England, Peterborough. Available to download from http://publications.naturalengland.org.uk/publication/2798159?category=45001 (Accessed 5 February 2014).
- Natural England. 2013c. Higher Level Stewardship. Environmental Stewardship Handbook, 4th edn. Natural England, Peterborough. Available to download from http://publications.naturalengland.org.uk/publication/2827091 (Accessed 5 February 2014).
- Peach, W.J., Siriwardena, G.M. & Gregory, R. 1999. Long-term changes in over-winter survival rates explain the decline of Reed Buntings Emberiza schoeniclus in Britain. J. Appl. Ecol. 36: 798–811.
- Perkins, A.J., Maggs, H.E. & Wilson, J.D. 2008. Winter bird use of seed-rich habitats in agri-environment schemes. Agr. Ecosyst. Environ. 126: 189–194.
- Pywell, R.F., Shaw, L., Meek, W., Turk, A., Shore, R.F. & Nowakowski, M. 2007. Do wild bird seed mixtures benefit other taxa? Asp. Appl. Biol. 81: 69–76.
- SAS Institute. 2003. The SAS System for Windows. Version 9.1. SAS Institute Inc, Cary, NC.
- Siriwardena, G.M., Baillie, S.R. & Wilson, J.D. 1999. Temporal variation in the annual survival rates of six granivorous birds with contrasting population trends. Ibis 141: 621–636.
- Siriwardena, G.M., Baillie, S.R., Crick, H.Q.P. & Wilson, J.D. 2000. The importance of variation in the breeding performance of seed-eating birds in determining their population trends on farmland. J. Appl. Ecol. 37: 128–148.
- Siriwardena, G.M., Calbrade, N.A. & Vickery, J.A. 2008. Farmland birds and late winter food: does seed supply fail to meet demand? Ibis 150: 585–595.
- Stoate, C., Szczur, J. & Aebischer, N.J. 2003. Winter use of wild bird cover crops by passerines on farmland in northeast England: declining farmland species were more abundant in these crops which can be matched to the birds’ requirements. Bird Study 50: 15–21.
- Stoate, C., Henderson, I.G. & Parish, D.M.B. 2004. Development of an agri-environment scheme option: seed-bearing crops for farmland birds. Ibis 146: 203–209.
- Stockdale, J.E., Dunn, J.C., Goodman, S.J., Morris, A.J., Sheehan, D.K., Grice, P.V. & Hamer, K.C. in press. The protozoan parasite Trichomonas gallinae causes adult and nestling mortality in a declining population of European Turtle Doves Streptopelia turtur. Parasitology. doi:10.1017/S0031182014001474
- Wilson, J.D., Arroyo, B.E. & Clark, S.C. 1996. The diet of birds species of lowland farmland: a literature review. Unpublished report to the Department of the Environment and Joint Nature Conservation Committee. University of Oxford and Royal Society for the Protection of Birds, Sandy.
- Wilson, J.D., Evans, A.D. & Grice, P.V. 2009. Bird Conservation and Agriculture. Cambridge University Press, Cambridge.