
Abstract
Capsule Sex-biased dispersal and an age-dependent effect in survival rate accounted for the pattern of first settlement and reproduction in a newly reintroduced Osprey population.
Aims We estimate the survival of translocated individuals, describe juvenile movements and evaluate the success of first breeding events to document the re-establishment of an Osprey breeding population.
Methods Between 2006 and 2010, 32 fledgling Ospreys were reintroduced via hacking techniques in Maremma Regional Park, Italy. We evaluated the effects of age on survival through multistate capture-mark-recapture analyses. Movements were investigated by radiotracking and using records of resightings.
Results Survival was high for juveniles after the release (0.87), markedly decreased during the first winter (0.26), and improved again in subsequent years (annual apparent survival of 0.69 for immatures and 0.93 for adults). Mean distance covered in initial dispersal was greater for females (246.2 km) than for males (38.7 km).
Conclusion Our results provided information on dispersal and survival rate of reintroduced Ospreys in a Mediterranean area. Despite low apparent survival in the first year, the high survival rates found in immatures and adults suggested favourable conditions for this new population. The study of demographic parameters is important for calibrating management actions aimed at the establishment of a self-sustaining Osprey population.
In any reintroduction project, the pre- and post-release management can affect both post-release survival and dispersal of translocated individuals, ultimately influencing their successful establishment and persistence (Armstrong & Seddon Citation2008). The study of the details of reintroduction programmes is therefore important for planning management and conservation strategies aimed at reducing post-fledging mortality (Soutullo et al. Citation2006). This is especially true for those populations in which movements and dispersal strategies are not well known and can affect an individual's survival rate. For example, in migratory populations the high energetic demands required by long non-stop flights during migratory journeys (Alerstam Citation1990) have been proved to be one of the primary causes of death (Sillet & Holmes Citation2002, Lok et al. Citation2013, Klaassen et al. Citation2014).
The Osprey Pandion haliaetus is a distinctive bird of prey widely distributed across different biogeographical regions of the world (Poole Citation1989). During the 19th and early 20th centuries it faced heavy direct persecution (e.g. shooting, egg-collection and habitat destruction; Poole Citation1989, Saurola Citation2005). As a result, many local populations disappeared (Cramp & Simmons Citation1980, Dennis & Dixon Citation2001). It is included in Annex I of the European Directive (2009/147/EC) on the conservation of wild birds and hence, considered a priority species for conservation along its whole distributional range. Around the Mediterranean, the conservation status of the species is nowadays considered unfavourable (Thibault et al. Citation2001, Muriel et al. Citation2010), with <80 breeding pairs distributed between Corsica, the Balearics, Morocco and Algeria (Monti Citation2012). Although direct management actions allowed a partial recovery in Corsica and in the Balearics, the Mediterranean population is still vulnerable (Thibault & Bretagnolle Citation2001, Thibault et al. Citation2001, Triay & Siverio, Citation2008, Monti et al. Citation2013). Furthermore, because of the high philopatry of the species, the natural recolonization of portions of the past range is unlikely to occur within a short time span. Accordingly, recovery of historical breeding sites by means of reintroduction has been considered as a key strategy for Osprey conservation in this area, since reintroductions are one of the best and reliable ways for the re-establishing of populations, once the past causes of extinction have been identified and eliminated (Griffith et al. Citation1989, Seddon et al. Citation2007). Three reintroduction projects for the Osprey were launched in Southern Europe: in 2003 in continental Spain, 2006 in Central Italy and 2011 in Portugal (Muriel et al. Citation2006, CIBIO Citation2011, Monti et al. Citation2012).
In Italy, the Osprey became extinct as a breeding species during the last years of the 1960s mainly due to direct persecution (Bulgarini et al. Citation1998, Brichetti & Fracasso Citation2003). The last reproduction events were reported for the islands of Sicily and Sardinia in 1968–1969 and continental Apulia region in 1955 (Frugis & Frugis Citation1963, Thibault & Patrimonio Citation1992, Brichetti & Fracasso Citation2003). The last known breeding occurrence for Central Italy was in Tuscany (Montecristo Island in 1929; Arrigoni degli Oddi Citation1929). In 2006, thanks to collaboration between the Natural Regional Park of Corsica (France) and the Maremma Regional Park (MRP) (Italy), a reintroduction programme was launched in Central Italy, aiming at re-establishing a breeding population that would potentially inter-connect with the nearby Corsican breeding population (Sforzi et al. Citation2007).
Since the Osprey population of the Mediterranean is thought to be mostly sedentary and/or characterized by individuals that perform reduced movements within the basin (Thibault et al. Citation1996), here we first report the movements and dispersal patterns of translocated birds to assess whether they remained in the area or moved far away. Second, we estimate apparent survival through mark-recapture models. Considering that the hacking technique used for reintroduction may affect the survival rates of young birds because of the lack of direct parental care during the first stages of life, we expected lower values of survival compared to those found in wild populations. In particular, we hypothesized that a high mortality rate in reintroduced birds may reduce the chances of reaching adulthood and in turn affect the success of the reintroduction programme. Finally, we would predict a greater survival rate for this relatively sedentary population compared to Osprey populations of North and Central Europe that perform hazardous long-distance migrations (Klaassen et al. Citation2014).
METHODS
Study site and field methods
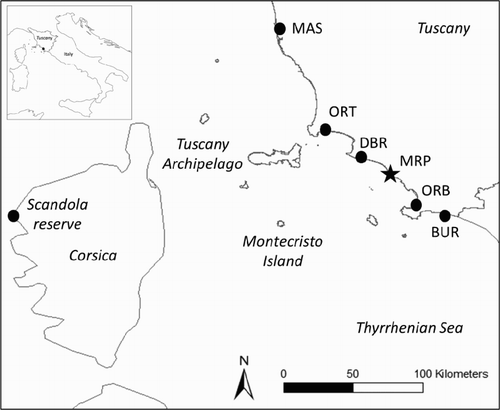
The choice of the reintroduction site took into account the strategic geographical position of the area as highly suitable for the future Osprey population expansion (Sforzi Citation2004, Dominici et al. Citation2007). The release site (42°39’ N, 11°05’ E) was located in the MRP, at the centre of an extensive coastal wetland system in Southern Tuscany. The system (hereafter called ‘study area’) is composed of several protected areas (): Burano Lake (BUR – WWF protected area – 40 km from the release site); Orbetello Lagoon (ORB – WWF natural reserve – 30 km from the release site); Diaccia Botrona Natural Reserve (DBR – 15 km from the release site); Orti-Bottagone Marsh (ORT – WWF natural reserve – 50 km from the release site); Massaciuccoli Lake (MAS – LIPU natural reserve – 140 km from the release site). Lagoons and saltwater marshes provide suitable fishing grounds for Ospreys. The release area is located in one of the Integral Reserves of the MRP at the mouth of the Ombrone River (for further details on this area see Monti et al. Citation2012).
Figure 1. Geographical location and abbreviations of the main sites along the coastal wetlands system in Tuscany: black dots from North to South of Tuscany, Massaciuccoli Lake (MAS), Orti-Bottagone Marsh (ORT), Diaccia Botrona Nature Reserve (DBR), Orbetello Lagoon (ORB) and Burano Lake (BUR). Black star: Maremma Regional Park (MRP = release site). The islands of the Tuscany Archipelago and the Scandola Reserve (black dot) in Corsica are also shown.
The Corsican Osprey population was used as the donor population. Although strong direct persecution reduced this population at only three breeding pairs in 1974, local management actions since then allowed a rapid demographic recovery. The population was considered stable with around 30 breeding pairs (Thibault et al. Citation2001, Bretagnolle et al. Citation2008). A total of 6–8 chicks was collected per year from the donor population without compromising its survival, for a minimum duration of five years of releases (Dominici et al. Citation2007). Juvenile Ospreys were taken from nests located along the west coast of Corsica (including the Scandola Natural Marine Reserve; 42°25’ N, 8°36’ E) at an age of 35–42 days from clutches of three eggs. The biggest and oldest chick was taken for translocation, while the younger chicks were left in the nest, where their chances of survival were increased in the absence of one sibling. Chicks were transported by helicopters to the hacking tower in the MRP, where they were kept for approximately three weeks until release. Each individual was marked with both a metal ring and a coloured darvic ring with an alpha-numeric code (for at distance identification) and equipped with a 10 g tail-mounted VHF radiotransmitter (Biotrack Ltd, UK).
Behaviour was monitored on a daily basis during the pre-release phase from dawn to dusk. Home ranges and space-use strategies were investigated throughout the post-fledging dependence period (PFDP) by direct observations and intensive VHF radiotracking (for details of the monitoring protocol see Monti et al. Citation2012). During the PFDP (July–August), juveniles mainly moved within a radius of about 1 km in the surroundings of the release pens. Nonetheless, they performed also greater movements, exploring larger areas (<15 km).
Since the radio battery life only lasted a maximum of three months (from June to September), individuals were detected through direct observations by reading ring codes after this period of telemetry. For each site of the study area, the presence of Ospreys was checked once a week, on average. Resightings of ringed birds were used to calculate the minimum distance covered and the main direction of juveniles' first movements.
Breeding attempts were strictly monitored. Early in the season (February–March) each site of the study area was visited two times per week to check for individuals performing courtship displays, constructing nests or trying to mate. Later on, occupied breeding sites were kept under daily surveillance (from April to August). Wild-born chicks were handled at 5–6 weeks after hatching to take body measures and to be marked with a metal ring and a coloured darvic ring with an alpha-numeric code.
Estimation of apparent survival
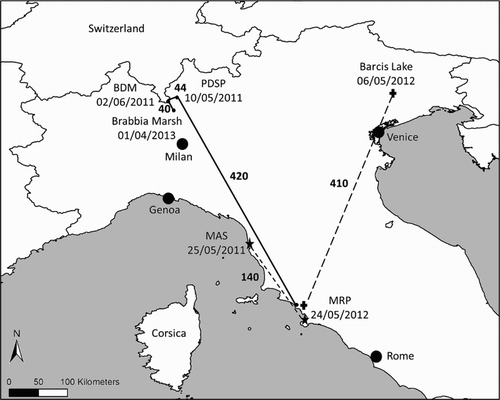
Using records of resightings and recoveries of marked non-territorial and territorial birds, a capture-mark-recapture (CMR) analysis was carried out on reintroduced birds. Overall, 185 resightings were collected during the period 2006–2013. Only four sightings have been reported from areas well beyond the Southern Tuscany wetland study area (); these data, obtained without protocol or constant field effort, were not included in the survival analyses. Thus, we considered 181 records.
Figure 2. Map showing long-distance dispersal movements of three reintroduced females in Italy: A4, released in 2009 (black dots and solid lines); L5, released in 2010 (black starts and sketched lines); S5, released in 2010 (black cross and dotted lines). For each location the name of the place or its abbreviation, the date of sighting and the minimum line of sight distance from the previous location (in bold and expressed in km) are shown.
The study area was checked every 1–2 days during the release phases (from June to August) and every week during the rest of the year. We assumed that the loss of coloured rings was rare and unlikely for juveniles and so this was ignored for analyses (Tavecchia et al. Citation2012, Mihoub et al. Citation2014). As we were mainly interested in estimating survival during the first phases of the reintroduction, we set four specific age classes according to the species' biology: (1) juveniles, during summer, before dispersal (from first ringing in July to end of August; 3–4 months of age; 2 months after fledging in the wild); (2) juveniles after dispersal (from September to February; 5–10 months of age); (3) immature birds; 10–20 months old; (4) adults older than 20 months. Two main periods of six months were considered for the analyses: summer (from March to August, corresponding to the breeding season) and winter (from September to February, corresponding to the wintering season). We also considered two sex categories (coded in two groups in our analyses). In addition, we estimated the annual apparent survival for immature and adult birds.
Recoveries of dead individuals only occurred in summer periods, in the study area, and only for juveniles at their first summer (no recoveries for immature or adults, or during winter periods), whereas resightings occurred all year round. Each observation (ringing, resighting or recovery) was coded according to three events, describing the status of the individual (dead or alive). We used the multistate framework to code individual encounter histories with one state ‘alive’ and one state ‘dead’, according to Lebreton et al. (Citation1999). Model selection was performed using the program E-SURGE (Choquet et al. Citation2009a) with an Akaike Information Criterion corrected for sample size (QAICc) (Burnham & Anderson Citation2002). In our initial model, the survival Φ and resighting p rates are age, time and sex-dependent while recovery r rate was kept constant. Considering that during the PFDP marked birds were closely monitored by intensive radiotracking, we set resighting and recovery rates equal to 1 for this period. Recovery rate was assumed to be zero after the first summer because of the battery failure of VHF transmitters and of no recovery data reported. Goodness-of-fit (GOF) tests of the initial model were performed using U-CARE 2.5 (Choquet et al. Citation2009b). Estimates are given with 95% confidence intervals following within brackets.
RESULTS
Movements and dispersal and first breeding events
Between 2006 and 2010, 32 juvenile Ospreys (20 females and 12 males) were translocated from Corsica to MRP (). From September onwards, 71.4% of birds moved and were not detected anymore, whereas 28.6% were observed again. In particular between 2006 and 2013, eight individuals (four males and four females) were recorded outside the release area. Three out of four males were resident during the subsequent years, visiting different wetlands, mainly located along the Tuscany coast (, ). The males settled in this area soon after leaving the study site and after it was intensively explored during four to five years, it finally became the place of first breeding attempts and reproduction.
Figure 3. The number of Ospreys observed in the study area per year: juveniles banded and released (black bars), number of immature individuals (grey bars), number of territorial adults that did not reproduce (white bars with skew lines), number of breeders (grey bars with skew lines) and number of wild-born chicks (white bars).
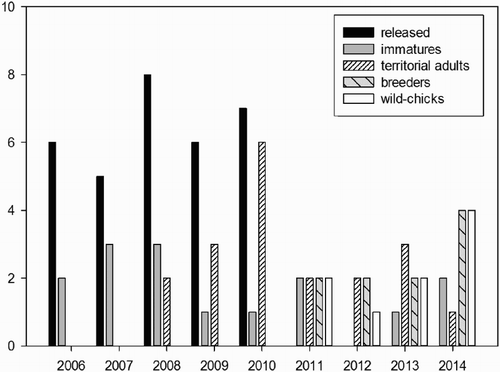
Table 1. Furthest movements of the released Ospreys, ID, sex, date of the end of the PFDP, date and time elapsed from the PFDP (days) until the first resighting, and location (with line of sight distance in km from the release site) are reported.
Occasional sightings reported by birdwatchers gave the opportunity to assess the minimum distance covered and the direction of juvenile Ospreys’ movements in their first year of life (). Four sightings were reported also from areas well beyond the Southern Tuscany wetland study area (). On average the distance covered after leaving the release area was greater for females (246.2 ± 201.5 km) than for males (38.7 ± 16.5 km). All females but one did not come back to the release area ().
First breeding attempts were performed in 2010 by three pairs in the study area (). All of them were made up of one reintroduced adult males (mean age = 3.6 ± 0.57 years) and a wild-born female of unknown origin (unmarked). One of these pairs, composed of a male released in 2006 (age 5) and an unmarked female, settled in a salty swamp in the MRP and successfully raised two chicks in 2011, one chick in 2012, two chicks in 2013 and one in 2014 (total of six chicks). Between 2011 and 2013, a second territorial pair settled in the MRP but failed to breed. Finally in 2014, a third pair composed of a reintroduced female (age 4) and a male wearing only a metal ring bred in the Diaccia Botrona Natural Reserve, producing three fledglings ().
Mortality and apparent survival
Four cases of mortality were recorded among the translocated birds (four females and one male). For three fledglings females, the cause of death was predation by carnivores (feathers with chewed up rachis base found). Electrocution occurred in one case: the dead body with burned feather tips was found on the ground under an electric pole located outside the MRP.
According to the GOF test, our initial model reasonably fits the data (χ2 = 9.1, df = 9, P = 0.43). The best model assumes a sex effect for resighting rate (larger for males (0.97 [0.81–0.99]) than for females (0.69 [0.40–0.88])) and an age effect for survival (). Survival rate for juveniles was very high (0.87 [0.71–0.95]) for the first two months (PFDP in the summer) and decreased to 0.26 [0.13–0.46] after dispersal and during their first winter (period September–February). Annual survival was estimated at 0.69 [0.29–0.92] for second-year birds (immature), and at 0.93 [0.65–0.99] for adults. To obtain an estimate comparable with those of other published studies, we pooled our data (originally split in three periods) to build up an annual cluster. We then multiplied survival rate of the first two months (0.87) × survival rate for six months after leaving the area (0.26) × a four-month survival rate extracted from the annual survival rate of immature (the cubic root of 0.69). In this way, we obtained a first-year apparent annual survival of 0.20.
Table 2. Model selection for survival and resighting rates with recovery rate as kept constant. Age consisted of four age classes: juveniles during summer, juveniles during winter, immatures and adults. Season was summer and winter periods, t means a time effect (i.e. variation between years), np is the number of identifiable parameters and QAICc is the Akaike Information Criterion corrected for sample size. Models are ranked by decreasing QAIC values.
DISCUSSION
In our translocated birds, the PFDP of three months and the onset of longer range dispersal in late August were within the range reported for the species (Cramp & Simmons Citation1980). Wild-born male Ospreys are known to be more philopatric than wild-born females and males generally cover shorter distances during the natal dispersal phase (Poole Citation1989, Johnson & Melquist Citation1991, Martell et al. Citation2002). Our data on translocated birds confirmed this behaviour, explaining the higher probability (97%) of resighting a male rather than a female (69%) in the study area.
Any reintroduction programme can be considered successful when the new population becomes self-sustaining (Schaub et al. Citation2009). Ospreys achieve sexual maturity (and start searching for a suitable territory for breeding) at the age of 2–4 years (Poole Citation1989, Englund & Green Citation2008). First breeding attempts of inexperienced pairs usually lead to a high percentage of failures, but this failure rate decreases in the following years as their experience grows (Poole Citation1989, Muriel et al. Citation2006). The settlement and breeding of the first pair hence constitutes an important milestone for the formation of a new viable population and for the attraction of other individuals in the area.
However in a reintroduction context, modelling population dynamics is an important tool to understand the demographic causes of variation in population sizes which may suggest how to re-orientate conservation actions to enhance the survival probability of young birds, hence enabling the success of a reintroduction project (Sarrazin & Barbault Citation1996). That is true especially for long-lived species in which low juvenile and high adult survival rates are expected (Clobert & Lebreton Citation1993). A few data are available from individuals released in reintroduction programmes in America (Hammer & Hatcher Citation1983, Rymon Citation1989, Martell et al. Citation2002), but no CMR analyses were carried out for a proper estimation of apparent survival. No data have been published so far for the Mediterranean area (whether wild and reintroduced populations). Although our data set of reintroduced individuals was rather small, our results showed a high survival (0.87) in the first stages of the PFDP, between the release and their departure. Such a high survival rate initially seems to exclude the possibility that there were negative effects related to difficulties encountered by fledglings because of the hacking method (e.g. lack of parental care and artificial feeding initially after the release). Analogous causes of mortality (e.g. predation, electrocution) were reported for other Osprey reintroduction projects (Mackrill Citation2005, Casado & Ferrer Citation2008, CIBIO Citation2011).
During the PFDP, young Ospreys returned to the hacking facilities to feed on artificial platforms (where fresh fish was supplied until the definitive departure of birds), while first fishing attempts were generally performed only after the departure (Monti et al. Citation2012). This is a crucial moment for young birds, which must rapidly learn how to capture live fish to survive to their first winter. In their first winter we found a very low survival rate (0.26), suggesting that juvenile birds likely encountered difficulties in catching prey, finding suitable habitat or dealing with unfavourable environmental conditions. By pooling these values we obtained a first-year annual survival of 0.20, i.e. 2.5 times below the survival probability of 0.50 obtained from wild-born juveniles from a population in continental France (Wahl & Barbraud Citation2014). However, the annual survival rate increased to 0.69 for second-year birds (immatures) but it was still lower than the survival rate of 0.87 estimated for the immatures and adults in Continental France (Wahl & Barbraud Citation2014). This suggests that reintroduced juveniles may be affected by the lack of parental care in the dispersal phase of their life (for example, fishing lessons from parents during the PFDP may result in higher survival of young once they are dispersed). Thus, it would be more interesting to compare our values with other reintroduced population in Europe (e.g. England) or in the Mediterranean area (e.g. Andalucía and Portugal) as soon as such analyses are available.
On the other hand, annual survival rate was relatively high (0.93) for adults compared to other wild populations: in Sweden 0.81 (Ryttmann Citation1994), the USA 0.85–0.90 (Spitzer et al. Citation1983) and continental France 0.87 (Wahl & Barbraud Citation2014). This result could be related to the current low population density in the area, and suggests favourable conditions for adults and low levels of competition with conspecifics for food resources and for territories, as already postulated in other reintroductions (Martell et al. Citation2002). A future increase in population size could possibly account for greater local competition with effects on demographic parameters, as recorded for the wild Corsican population (Bretagnolle et al. Citation2008). Another factor which could play in favour of a high adult survival in our case might be related to the fact that most of the breeders were resident and did not migrate, but rather spent the winter close to the breeding areas in Italy, thus avoiding the risk and energy costs that accompany a long-distance migration. For northern Ospreys, Klaassen et al. (Citation2014) estimated mortality rate six times higher during migration than during stationary periods, suggesting that events during the migration have an important impact on the population dynamics of long-distance migrants. However, the Mediterranean basin is known to be an area where direct persecution by human (e.g. hunting and poaching) is widespread (millions of birds, including also protected species such as raptors, are killed annually; www.birdlife.org). This aspect might increase mortality for those individuals spending all their lives in the Mediterranean basin, and especially for inexperienced juvenile birds.
Finally, these high adult survival rates were calculated on a very small sample size and this young population only contains young adults with high survival expectancy and no senescent individuals as yet. Further studies should include larger Osprey populations in the Mediterranean, monitored for a longer period (e.g. wild populations from Corsica and Balearic islands; reintroduced populations in Andalucía).
Reintroduction has proved to be an appropriate method to locally re-establish Osprey populations. Although, results from European experiences (i.e. reintroductions in Rutland Water – England and Andalucía – Spain) show that to achieve the goal of a self-sustaining population which does not require extra human intervention (e.g. supplementary translocations), both a long time span and/or a high number of individuals to be released are needed. In particular, 75 Scottish birds were released between 1996 and 2001 in England before obtaining 7 territorial pairs in 2013 (Tim Mackrill pers. comm.); 191 northern Ospreys (from Germany, Scotland and Finland) were used between 2003 and 2010 for the reintroduction in Andalucía before obtaining 12 territorial pairs in 2013 (Eva Casado pers. comm.). In Italy, despite constraints related to the limited number of chicks available for translocation each year (e.g. the donor population in Corsica being relatively small), two pairs are currently breeding in the area eight years after the start of the project and several territorial individuals are regularly seen in the Southern Tuscany coastal wetland study area. This is quite comparable to the projects in England and Andalucía that are classified as successful.
Here, we have estimated the apparent survival rate of the re-establishing population as a first step to monitor the demographic parameters of the new population. Once more data and a longer time period of data are available, a proper population viability analyses will allow us to estimate the growth rate and probability of extinction of the population taking into account number of breeding and territorial pairs, fecundity and recruitment of other eventual wild-born individuals. A gradual increase in the number of breeding pairs is now expected in the wetland system of coastal Tuscany. Furthermore, the seven islands included in the Tuscany Archipelago National Park and located midway between Corsica and Tuscany could function as ‘stepping-stones’ and might play a relevant role for the future expansion of the species. The recent building of artificial nests in Corsica, Tuscany (coastal territories and Montecristo Island) and Sardinia (Porto Conte Regional Park, Alghero province) could stimulate the colonization of new sites by mature Ospreys, so favouring the process of natural expansion of the populations. These structures aim to attract floaters and encourage the first phases of settlement and reproduction. This would ensure both local conservation to the species and genetic exchanges between the two breeding populations (Corsica and Italy). Overall this may ensure the re-establishment of at least a part of the ancient range of Osprey distribution in the Central Mediterranean basin.
ACKNOWLEDGEMENTS
We would like to thank the Corsica Regional Park for the constant support and for providing the Osprey nestlings. Thanks also to Roberto Muriel and Pertti Saurola for their important advice during the initial phases of the project. For the precious support in fieldwork during the release days we are grateful to Tim Mackrill and John Wright (from Rutland Water, UK). For their contribution in the resightings of ringed birds we are in debt with Beppe Anselmi, Gianfranco Martini, Francesco Pezzo, Fabio Cianchi, Pietro Giovacchini, Paolo Fastelli, Luca Becherucci, Paolo Maria Politi, Nicola Maggi, Sandro Canci and Alessandro Troisi. Vincenzo Rizzo Pinna, Alessia Martelli and Elena Germondari provide field support and data recording for the monitoring of the breeding pairs. We want to thank also Dr Ettore Randi, head of the genetic laboratory of the ISPRA who performed the genetic determination of sex from feather samples. We are indebted to the park wardens and the administrative personnel of the Maremma Regional Park for the logistic support provided.
Funding
Funding was provided by the Maremma Regional Park Agency and the Tuscany Region Administration. FM benefits from a grant from the Maremma Regional Park, from the Foundation Prince Albert II de Monaco and the Associazone Italiana della Fondation Prince Albert II de Monaco ONLUS and from a mobility grant from the Università Italo Francese/Université Franco Italienne.
Related Research Data
REFERENCES
- Alerstam, T. 1990. Bird Migration, 420. Cambridge University Press, Cambridge.
- Armstrong, D.P. & Seddon, P.J. 2008. Directions in reintroduction biology. Trends Ecol. Evol. 23: 20–25. doi: 10.1016/j.tree.2007.10.003
- Arrigoni degli Oddi, E. 1929. Ornitologia italiana, 440–441. Ulrico Hoepli, Milano.
- Bretagnolle, V., Mougeot, F. & Thibault, J.C. 2008. Density dependence in a recovering Osprey population: demographic and behavioural processes. J. Anim. Ecol. 77: 998–1007. doi: 10.1111/j.1365-2656.2008.01418.x
- Brichetti, P. & Fracasso, G. 2003. Ornitologia italiana. Vol. 1 – Gaviidae-Falconidae, 290–293. Alberto Perdisa Editore, Bologna.
- Bulgarini, F., Calvario, E., Fraticelli, F., Petretti, F. & Sarrocco, S. 1998. Libro Rosso degli Animali d'Italia – Vertebrati, 210. WWF Italia, Roma.
- Burnham, K.P. & Anderson, D.R. 2002. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd edn. Springer, New-York.
- Casado, E. & Ferrer, M. 2008. Osprey Pandion haliaetus reintroduction project in Andalucia. Report 2008: 40.
- Choquet, R., Rouan, L. & Pradel, R. 2009a. Program E-SURGE: a software application for fitting Multievent models. In Thomson, D.L., Cooch, E.G. & Conroy, M.J. (eds.) Springer Series: Environmental and Ecological Statistics, Vol. 3: 845–865. Springer, New York.
- Choquet, R., Lebreton, J.D., Gimenez, O., Reboulet, A.M. & Pradel, R. 2009b. U-CARE: utilities for performing goodness of fit tests and manipulating CApture–REcapture data. Ecography 32: 1071–1074. doi: 10.1111/j.1600-0587.2009.05968.x
- CIBIO (Centro de Investigação em Biodiversidade e Recursos Genéticos). 2011. Reintroduction of the Osprey (Pandion haliaetus) in Portugal. Annual report 2011: 51.
- Clobert, J. & Lebreton, J.D. 1993. Estimation of demographic parameters in bird populations. In Perrins, C.M., Lebreton, J.D. & Hirons, G.J.M. (eds.) Bird Population Studies, 75–102. Oxford University Press, Oxford.
- Cramp, S. & Simmons, K.E.L. 1980. Handbook of the Birds of Europe, The Middle East and North Africa. The Birds of the Western Palearctic, Vol. 2: 265–277. Oxford University Press, Oxford.
- Dennis, R. & Dixon, H. 2001. The experimental reintroduction of Ospreys Pandion haliaetus from Scotland to England. Vogelwelt 122: 147–154.
- Dominici, J.M., Susini, P.A. & Thibault, J.C. 2007. The Osprey in Corsica: from the near-extinction to the hacking project with the Maremma Park (Tuscany). In International meeting in Western-Palearctic Osprey populations (1–2 December 2007), Alberese.
- Englund, J.V. & Greene, V.L. 2008. Two-year-old nesting behavior and extra-pair copulation in a reintroduced Osprey population. J. Raptor Res. 42: 119–124. doi: 10.3356/JRR-05-71.1
- Frugis, S. & Frugis, D. 1963. Le Paludi pugliesi a sud del Gargano. Riv. ital. Ornitol. 33: 79–123.
- Griffith, B., Scott, J.M., Carpenter, J.W. & Reed, C. 1989. Translocation as a species conservation tool: status and strategy. Science 245: 477–480. doi: 10.1126/science.245.4917.477
- Hammer, D.A. & Hatcher, R.M. 1983. Restoring Osprey populations by hacking preflighted young. In Bird, D.M., Seymour, N.R. & Gerrard, J.M. (eds.) Biology and Management of Bald Eagles and Ospreys, 293–297. Harpell Press, Ste. Anne de Bellevue.
- Johnson, D.R. & Melquist, W.E. 1991. Wintering distribution and dispersal of northern Idaho and eastern Washington Ospreys. J. Field Ornithol. 62: 517–520.
- Klaassen, R.H.G., Hake, M., Strandberg, R., Koks, B.J., Trierweiler, C., Exo, K.M., Bairlein, F. & Alerstam, T. 2014. When and where does mortality occur in migratory birds? Direct evidence from long-term satellite tracking of two raptors. J. Anim. Ecol. 83: 176–184. doi: 10.1111/1365-2656.12135
- Lebreton, J.D., Almeras, T. & Pradel, R. 1999. Competing events, mixture of information and multistratum recapture models. Bird Study 46S: S32–S38.
- Lok, T., Overdijk, O. & Piersma, T. 2013. Migration sometimes takes a toll: spoonbills suffer higher mortality during longer northward migrations. Spoonbills as a model system, a demographic cost-benefit analysis of differential migration (T. Lok), chapter 8. PhD Thesis, University of Groningen.
- Mackrill, T. 2005. Rutland Water Osprey Project. Annual Report 2005: 47.
- Martell, M.S., Englund, J.V. & Tordoff, H.B. 2002. An urban Osprey population established by translocation. J. Raptor Res. 36: 91–96.
- Mihoub, J.B., Princé, K., Duriez, O., Lécuyer, P., Eliotout, B. & Sarrazin, F. 2014. Comparing the effects of release methods on survival of the Eurasian black vulture Aegypius monachus reintroduced in France. Oryx 48: 106–115. doi: 10.1017/S0030605312000981
- Monti, F. 2012. The Osprey, Pandion haliaetus: state of knowledge and conservation of the breeding population of the Mediterranean basin. Initiative PIM: 26.
- Monti, F., Sforzi, A. & Dominici, J.M. 2012. Post-fledging dependence period of Ospreys Pandion haliaetus released in central Italy: home ranges, space use and aggregation. Ardeola 59: 17–30. doi: 10.13157/arla.59.1.2012.17
- Monti, F., Nibani, H., Dominici, J.M., Idrissi, H.R., Thevenet, M., Beaubrun, P.C. & Duriez, O. 2013. The vulnerable Osprey breeding population of the Al Hoceima National Park, Morocco: present status and threats. Ostrich 84: 199–204. doi: 10.2989/00306525.2013.865280
- Muriel, R., Ferrer, M., Casado, E. & Schmidt, D. 2006. First breeding success of Osprey (Pandion haliaetus) in mainland Spain since 1981 using cross-fostering. J. Raptor Res. 40: 303–304. doi: 10.3356/0892-1016(2006)40[303:FBSOOP]2.0.CO;2
- Muriel, R., Ferrer, M., Casado, E. & Calabuig, C.P. 2010. First successful breeding of reintroduced Ospreys Pandion haliaetus in mainland Spain. Ardeola 57: 175–180.
- Poole, A.F. 1989. Ospreys: A Natural and Unnatural History, 246. Cambridge University Press, Cambridge.
- Rymon, L.M. 1989. The restoration of Ospreys, Pandion haliaetus, to breeding in Pennsylvania by hacking (1980–89). In Meyburg, B.U. & Chancellor, R.D. (eds.) Raptors in the Modern World, 359–362. WWGBP, Berlin.
- Ryttman, H. 1994. Estimates of survival and population development of the Osprey Pandion haliaetus, Common Buzzard Buteo buteo, and Sparrowhawk Accipiter nisus in Sweden. Ornis Svec. 4: 159–172.
- Sarrazin, F. & Barbault, R. 1996. Reintroduction: challenges and lessons for basic ecology. Trends Ecol. Evol. 11: 474–478. doi: 10.1016/0169-5347(96)20092-8
- Saurola, P. 2005. Monitoring and conservation of Finnish Ospreys Pandion haliaetus in 1971–2005. Status of Raptor Populations in Eastern Fennoscandia. Proceedings of the workshop, Kostomuksha, Kaerelia, Russia, November 8–10, 2005: 125–132.
- Schaub, M., Zink, R., Beissmann, H., Sarrazin, F. & Arlettaz, R. 2009. When to end releases in reintroduction programmes: demographic rates and population viability analysis of bearded vultures in the Alps. J. Appl. Ecol. 46: 92–100. doi: 10.1111/j.1365-2664.2008.01585.x
- Seddon, P.J., Armstrong, D.P. & Maloney, R.F. 2007. Developing the science of reintroduction biology. Conserv. Biol. 21: 303–312. doi: 10.1111/j.1523-1739.2006.00627.x
- Sforzi, A. 2004. Report: ricostituzione di una popolazione nidificante di Falco pescatore (Pandion haliaetus) nel Parco Regionale della Maremma. Parco Regionale della Maremma, Grosseto.
- Sforzi, A., Monti, F. & Sammuri, G. 2007. Re-establishing an Osprey breeding population in the coastal Tuscany: the Maremma translocation project. In International meeting in Western-Palearctic Osprey populations (1–2 December 2007), Alberese. http://balbuzard.lpo.fr/actualites/Abstrats_colloque-international_Italie07.pdf
- Sillett, T.S. & Holmes, R.T. 2002. Variation in survivorship of a migratory songbird throughout its annual cycle. J. Anim. Ecol. 71: 296–308. doi: 10.1046/j.1365-2656.2002.00599.x
- Soutullo, A., Urios, V., Ferrer, M. & Peñarrubia, S.G. 2006. Post-fledging behaviour in Golden Eagles Aquila chrysaetos: onset of juvenile dispersal and progressive distancing from the nest. Ibis 148: 307–312. doi: 10.1111/j.1474-919X.2006.00530.x
- Spitzer, P.R., Poole, A.F. & Schebel, M. 1983. Initial recovery of breeding Ospreys in the region between New York City and Boston. In Bird, D.M. (ed.) Biology and Management of Bald Eagles and Ospreys, 231–241. Harpell Press, Ste Anne de Bellevue.
- Tavecchia, G., Adrover, J., Muñoz Navarro, A. & Pradel, R. 2012. Modelling mortality causes in longitudinal data in the presence of tag loss: application to raptor poisoning and electrocution. J. Appl. Ecol. 49: 297–305. doi: 10.1111/j.1365-2664.2011.02074.x
- Thibault, J.C. & Bretagnolle, V. 2001. Monitoring, research and conservation of Osprey Pandion haliaetus on Corsica, Mediterranean, France. Vogelwelt 122: 173–178.
- Thibault, J.C. & Patrimonio, O. 1992. Falco pescatore. Fauna d'Italia, 621–632. Uccelli I. Edizioni Calderini, Bologna.
- Thibault, J.C., Triay, R., Beaubrun, P., Boukhalfa, D., Dominici, J.M. & Torre, A. 1996. Osprey (Pandion haliaetus) in the Mediterranean: characteristic of a resident population with a patchy distribution. In Muntaner, J. & Mayol, J. (eds.) Biologia y Conservaciòn de las Rapaces Mediterràneas, 135–144. Monografias, no 4. SEO, Madrid.
- Thibault, J.C., Bretagnolle, V. & Dominici, J.M. 2001. Le Balbuzard pêcheur en Corse: du martyre au symbole de la protection de la nature, 184. Alain Piazzola, Ajaccio.
- Triay, R. & Siverio, M. 2008. El aguila pescadora en España. Poblacion en 2008 y metodo de censo. SEO/Birdlife. Madrid: 82.
- Wahl, R. & Barbraud, C. 2014. The demography of a newly established Osprey Pandion haliaetus population in France. Ibis 156: 84–96. doi: 10.1111/ibi.12114