
Abstract
Capsule Rock Ptarmigan place their nests adjacent to boulders, in their lee and on their sunnier aspects.
Aims To investigate the proximity and orientation of Rock Ptarmigan nests in relation to boulders and vegetation height.
Methods The distances from boulders of 61 Rock Ptarmigan nests and 60 random points were compared. Orientation variables of nests from boulders were analysed with circular statistical formulae.
Results Rock Ptarmigan nests were closer to boulders than random points. The mean orientation of nests relative to adjacent boulders was south-east and the prevailing wind was westerly. Vegetation height at nest sites was taller than at randomly selected sites.
Conclusion Rock Ptarmigan which nest next to, in the lee of, and on the south-eastern aspects of boulders, probably gain advantages of protection from the prevailing wind and passive thermoregulation from solar heat.
Thermoregulatory demands account for approximately half of a bird's energy expenditure and energy transfer by heat loss or gain is determined by the microhabitat the animal selects (Walsberg Citation1985). The microclimate of a bird's nest site can affect the incubating bird's thermoregulation (With & Webb Citation1993, Nelson & Martin Citation1999) and its efficiency of hatching its eggs (Burton Citation2006, Butler et al. Citation2009). The orientation of nests, as in tree cavities or nest boxes, can affect the microclimate of nests and the nesting birds' reproductive success, (Wiebe Citation2001, Goodenough et al. Citation2008), and Tree Swallows Tachycineta bicolor select south- and east-facing nest boxes which have higher internal morning temperatures than north- and west-facing boxes (Ardia et al. Citation2006).
Choice of nest site cover by ground-nesting birds is a simple way for a bird to increase protection from unfavourable weather or concealment from predators, and birds can enhance the microclimate around their nest by selecting a favourable orientation relative to the surrounding cover, prevailing wind direction and solar radiation. For example, the Australasian Pipit Anthus novaeseelandiae in the Snowy Mountains of Australia, a ground-nesting bird, orientates its nest entrance away from prevailing cold fronts (Norment & Green Citation2004), and other alpine species orientate their nests to face the morning sun, such as the Water Pipit Anthus spinoletta (Rauter et al. Citation2002) and the North American Gray-headed Junco Junco caniceps, Lincoln's Sparrow Melospiza lincolnii and the White-crowned Sparrow Zonotrichia leucophrys (Hadley Citation1969). Nest site selection may provide benefits both with respect to shelter from wind and exposure to solar radiation, and it is probably often not possible to determine which, if either is of primary importance. Microclimates vary and the importance of either criterion likely varies too, e.g. the orientation of nests among ground-nesting birds is more directional towards the sun in higher latitudes (Burton Citation2007).
The Rock Ptarmigan Lagopus muta is a ground-nesting grouse which lives in Arctic-alpine tundra where vegetation cover is mostly short and variable in degree of protection from weather and predators. Rock Ptarmigan have a circumpolar distribution, and in Japan, where the vegetation in the species' range has a mean height of approximately 50 cm, the birds selectively nest among shorter shrubs of approximate mean height 38 cm (Sawa et al. Citation2011) while in the European Alps, where the vegetation in their habitat is generally shorter, they selectively nest in locally taller shrub cover (Scherini et al. Citation2003). In Svalbard, where there is no shrub cover, they nest in rocky situations (Steen & Unander Citation1985), and in Scotland where the local race Lagopus muta millaisi live in boulder-strewn, Arctic-alpine ground they often nest in the open, close to surface boulders (Watson Citation1972, Watson & Moss Citation2008). Birds in Scotland which nest close to boulders have been anecdotally recorded as nesting in such sites because they gain shelter from wind (MacDonald Citation1970, Watson Citation1972) and a cryptic background for concealment from predators (Watson & Moss Citation2008), however, none of this has been tested. The vegetation in the alpine ground where Rock Ptarmigan in Scotland live is very short or prostrate, less than 15 cm (Watson & Rae Citation1993), and not as tall as that selected in Japan or the Alps, so it is possible that Scottish Rock Ptarmigan use boulders for cover in lieu of taller vegetation. If the birds use boulders for shelter from the prevailing wind, they would be expected to place their nests on the leeward side of such, perhaps more so by ptarmigan nesting on windward hill slopes.
The aim of this study was to determine the distance and orientation of Ptarmigan nests from boulders and the vegetation height around the nests relative to random points, to test whether Rock Ptarmigan selectively place their nests near boulders or in tall vegetation thus providing the birds with concealment from predators, in positions that provide greater shelter from the prevailing wind, and greater exposure to sunshine.
METHODS
The study area was in the eastern Scottish Highlands on the adjacent hills, Cairnwell and Meall Odhar (56°52′N, 3°25′W) during 1989–1992 and 2012. The climate is subarctic-oceanic with snowfall in any month (Watson et al. Citation1998) and the habitat used by the birds was short or prostrate Arctic-alpine heath dominated by Heather Calluna vulgaris, Bilberry Vaccinium myrtillus and Crowberry Empetrum nigrum, with some swards of alpine grassland, mostly Mat Grass Nardus stricta. Boulders were scattered throughout the study area as it lay in an area of glacial deposits, with shallow peat and topsoils. Potential predators of incubating birds or their eggs were Golden Eagle Aquila chrysaetos, Northern Raven Corvus corax, Red Fox Vulpes vulpes and Stoat Mustela erminea.
Most nests were found by watching birds back to their nests and the rest by visually searching within the birds' territories until all these were found. This eliminated any bias in sampling towards any possibility of finding nests in easier to see locations. Most breeding hens were one year old and most in 1989–1992 were marked with individual combinations of colour-rings. No marked birds were recorded in more than two years and the oldest hen Rock Ptarmigan recorded elsewhere has been seven years old (Holder & Montgomerie Citation1993), therefore all unmarked birds in 2012 were considered to be new subjects. There were 73 nest records in total which, to avoid pseudoreplication, was reduced to 61, removing any second nest by a bird and any records from nests of unmarked birds other than in 2012. The nests were all set in well-vegetated ground, in a shallow scrape lined with plant material most likely collected from nearby. Once the chicks had hatched and left the nest, the number of unhatched eggs was noted, and the distance was measured between each nest and the nearest boulder of a size which might give shelter to a Ptarmigan. As this size could not identified a priori, it was considered that such a boulder be at least as wide as a Ptarmigan (approximately 15 cm), for this would likely provide cover from wind, and as only birds nesting within a certain distance of a boulder would likely be sheltered, a distance of 25 cm from a boulder, the approximate length of a Ptarmigan, was used as a criterion for a distance at which any microclimate related to a boulder might affect a nesting bird. A line was taken between the centre of each nest and the closest part of the nearest boulder; the distance was measured and the orientation of the nest relative to the boulder was measured to the nearest degree with a compass then corrected to true north. Vegetation height around each nest was measured by recording the sward height of the vegetation within 10 cm of the edge of the nest at four ordinate points and the mean taken. The distances and orientations from boulders and heights of vegetation were also assessed at 60 computer generated random locations within a boundary delineated by a minimal convex polygon based on the locations of ptarmigan. These locations were mapped during daily observations of all ptarmigan behaviour during the nesting period. As the study site covered windward and leeward hill slopes in relation to the prevailing wind, nest site characteristics on each of these were compared as well as all the data as a whole. All mean values are presented with ± standard errors.
Differences in the distance to the nearest boulder and vegetation height between nests and random points were analysed with t-tests using log-transformed data to normalize the residuals. Tests as to whether the orientation of nests relative to the nearest boulder differed from a uniform orientation were undertaken using Rao's spacing Test (U) using Oriana (Kovach Citation2010). Rao's spacing test was used for statistical analyses of circular data because this has advantages over other commonly used goodness-of-fit tests, especially if any sample is possibly polymodal (Bergin Citation1991). Because daily records of wind directions were grouped to the nearest 45°, any analyses involving these were done with Rayleigh's (Z) and χ2 tests. Wind direction was recorded on site at midday for each day the observer was in the field during the study.
RESULTS
Rock Ptarmigan nests were significantly closer to boulders than randomly selected points, (t-test of log n transformed data, t = 8.3, df = 119, P < 0.001, two-tailed) (). Nests were within 7–108 cm of a boulder (mean = 19 ± 2.63 cm, median = 11 cm, n = 61) and random points were within 0–755 cm (mean = 189 ± 26.3 cm, median = 131 cm, n = 60). Nineteen nest centres were within 7 cm, which would entail the incubating bird abutting the boulder as the mean nest radius was 7 cm. Vegetation height at nest sites ranged from 3 to 28 cm (mean = 9.61 ± 0.53 cm, n = 61); that at random sites ranged from 0 to 22 cm (mean = 6.53 ± 0.53 cm). The difference in vegetation height between nest sites and random points was dependent on whether the nest was within 25 cm of a boulder (Two-way ANOVA, F1,119 = 18.2, P < 0.001). The height of the vegetation at nest sites (9.73 ± 0.63 cm) within 25 cm of a boulder was significantly taller (P < 0.01, n = 48, 14) than at random sites (3.36 ± 0.64 cm) but there was no difference (P = 0.12, n = 13, 46) between vegetation heights at nests (9.38 ± 0.97) and random sites (7.28 ± 0.59) greater than 25 cm from a boulder. The mean wind direction calculated from 254 daily observations was 260° (Rayleigh's Z-test = 69.2, P < 0.001), which was similar to the west/south-west prevailing wind direction as recorded at Braemar, the nearest meteorological station 15 km away (Meteorological Office). The orientations of the nesting sites in relation to a boulder were assessed as a whole and in two subsets; one of nests on slopes facing the prevailing wind and the other on leeward slopes. The nests on the windward south-west facing slopes were on aspects within 90° of the mean direction of the wind, i.e. 260 ± 90°, 170–350°, and nests on the leeward north-east facing slopes were within the remaining 180° (). There were 48 nests within 25 cm of a boulder, 32 were on a north-eastern slope and 16 on slopes with a south-western aspect.
Figure 1. Distances of Rock Ptarmigan nests (n = 61) and random points (n = 60) from the nearest boulder.
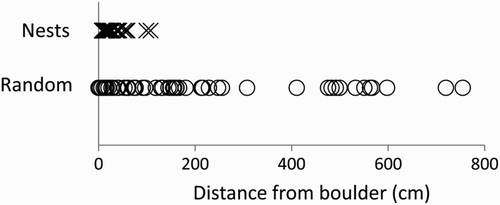
Figure 2. Orientation of Rock Ptarmigan nests from the nearest boulder on: (a) north-east (n = 32) and (b) south-west slopes (n = 16). Radial lines indicate the respective mean orientations 124° and 108° with 95% confidence intervals.
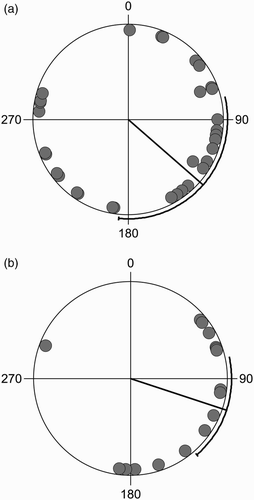
The distribution of nest orientations was significantly non-uniform for all nests (Rao's spacing test: U = 166, P < 0.01, n = 48), for nests on north-east aspects (U = 161, P < 0.05, n = 32) and for nests on south-west aspects (U = 180, P < 0.05, n = 16). The mean nest orientations were 115 ± 17.6° for all nests, 124 ± 36.0° for nests on a north-east slope and 108 ± 15.8° for nests on a south-west slope. Although birds nesting on a north-eastern slope placed their nests in a wider range of orientations than those nesting on a south-westerly slope, there was no significant difference between the mean orientations or variances of the orientations of the two subsets (Mardia–Watson–Wheeler Test W = 2.9, P = 0.24, n = 16, 32). Ptarmigan tended to nest on the south-eastern aspect of boulders next to their nest, significantly different from the direction of the recorded wind (χ2 = 89.5, P < 0.001, df = 7, class width 45°), and no nests were in the north-west quadrant between 291° and 0° from the nearest boulder. Birds that placed their nests on south-eastern aspects of boulders nested closer to boulders (circular–linear correlation, r = 0.33, P = 0.006, n = 48, Oriana), but there was no circular–linear correlation between sward height at nests and the orientation of the nest to nearest boulders (P = 0.42, Oriana) and no difference between the sward height at nests on north-eastern (9.19 ± 0.78 cm) and south-western slopes (9.75 ± 1.04 cm) (t = 0.19, df = 46, P = 0.85, two-tailed).
There were too few nests affected by predation to allow statistical analysis of any relationship between nest site characteristics and predation. Three clutches were eaten by predators in 1991 (likely by Northern Raven or Stoat because these were seen hunting in the study area at the time) and no incubating hens were killed while on a nest.
DISCUSSION
Rock Ptarmigan nesting in the Arctic-alpine ground in Scotland likely seek shelter from the prevailing west/south-westerly wind by selecting nesting sites close to and in the lee of boulders and in taller vegetation. When in the lee of such boulders they also tend to be on their warmer southern aspects.
In a study of White-tailed Ptarmigan Lagopus leucura, Rock and Willow Ptarmigan Lagopus lagopus, Wilson & Martin (Citation2008) found that a tendency for females to nest on warmer west- or south-facing slopes would also offer thermoregulatory benefits. In the present study, the Rock Ptarmigan hens were restricted to nest within their cocks' territories which were on various hillside aspects so they could not all choose to nest on a warmer slope. However, by selecting to nest on the southern side of boulders the birds could gain heating from solar radiation and possibly also heat which had been absorbed by and later radiated from the rock. Also, birds avoided the north-west side of boulders which might be critical nesting behaviour because this aspect would likely be exposed to the prevailing wind and receive less sunshine. Although they are covered with dense feathering, birds absorb radiant heat energy (Lustik et al. Citation1970), and shelter from wind can provide a bird with more thermal benefit than improvement of radiation balance (Walsberg Citation1986). Birds can also adjust their metabolic rate by shifting between shaded or windy sites and those protected from wind or exposed to solar radiation (Wolf & Walsberg Citation1996). Rock Ptarmigan often sunbathe in positions sheltered from wind (Watson Citation1972), and during this study birds seen doing so were often on the sunny lee side of boulders, similar to their nesting behaviour.
Survival of a bird embryo is limited by incubation schedule, heat transfer to the eggs and choice of a thermally appropriate nest site (Webb Citation1987) and nests can be placed in relation to heat gain from solar radiation (Walsberg Citation1985). For example, Tree Pipits Anthus trivialis in southern Britain where the prevailing wind is also WSW, orientate their nests predominantly facing east, a direction that provides shelter from wind and allows warming by the morning sun and more young hatched from such nests (Burton Citation2006). Similarly in the southern hemisphere, Tawny Frogmouth Podargus strigoides nests are placed on branches orientated to the north-east providing shelter from the prevailing wind and exposure to the morning sun (Rae & Rae Citation2013). In the present study, the mean aspect was to the south-east, which would expose the birds to the morning sun, but reduce their exposure to the sun in the warmer afternoon hours. In White-tailed Ptarmigan, eastern-facing nests sheltered by rocks heated up most quickly in the morning and were warmer than other nests at 09:00 hours, but thereafter declined in temperature and were coolest during the majority of the day (Wiebe & Martin Citation1997).
As only three clutches of eggs and no incubating hens were taken by predators, any advantage of concealment from aerial predators by nesting close to boulders could not be tested as proposed in this study. To hide from an overhead raptor, Rock Ptarmigan sidle up to a boulder similar in colour to them and remain motionless (Watson & Moss Citation2008). Similar behaviour has been documented in cryptically coloured moths, which choose to land on backgrounds which resemble their colouring (Sargent Citation1966), they further adjust their crypsis by then orientating themselves for their colour and shape to match that of the substrate colour and shape (Kang et al. Citation2012). Similarly, cryptically coloured Tawny Frogmouths nest on and orientate themselves along like-coloured branches to avoid detection (Rae & Rae Citation2013). As Rock Ptarmigan in Scotland nest in short Arctic-alpine vegetation, concealment from predators by disruption of an incubating bird's outline would be advantageous, and the paucity of such predation itself in this study might be evidence of the effectiveness of this strategy.
The primary criterion for choice of location by nesting Ptarmigan appears to be shelter from wind. Thermoregulation is a critical factor in nest site selection for birds in Arctic-alpine zones, and nesting Rock Ptarmigan worldwide select the best local microhabitats available in different regional habitats. In Scotland, Rock Ptarmigan select to nest in the lee of and in sunny aspects of boulders which aid their thermoregulation.
ACKNOWLEDGEMENTS
I am grateful to Ian Patterson, Des Thompson and Adam Watson for early discussion on this project. Raymond and Judy Duncan gave valuable assistance in the field, especially in catching and marking birds. Duncan Rae and John Rawsthorne provided comments which helped improve an earlier draft. Comments from the Editor W. Cresswell and two anonymous reviewers helped improve the final draft.
REFERENCES
- Ardia, D.R., Pérez, J.H. & Clotfelter, E.D. 2006. Nest box orientation affects internal temperature and nest site selection by Tree Swallows. J. Field Ornithol. 77: 339–344. doi: 10.1111/j.1557-9263.2006.00064.x
- Bergin, T.M. 1991. A comparison of goodness-of-fit tests for analysis of nest orientation in Western Kingbirds (Tyrannus verticalis). The Condor 93: 164–171. doi: 10.2307/1368619
- Burton, N.H.K. 2006. Nest orientation and hatching success in the tree pipit Anthus trivialis. J. Avian Biol. 37: 312–317. doi: 10.1111/j.2006.0908-8857.03822.x
- Burton, N.H.K. 2007. Intraspecific latitudinal variation in nest orientation among ground-nesting passerines: a study using published data. The Condor 109: 441–446. doi: 10.1650/0010-5422(2007)109[441:ILVINO]2.0.CO;2
- Butler, M.W., Whitman, B.A. & Dufty Jr., A.M. 2009. Nest box temperature and hatching success of American Kestrels varies with nest box orientation. Wilson J. Ornithol. 121: 778–782. doi: 10.1676/08-124.1
- Goodenough, A.E., Hart, A.G. & Elliot, S.L. 2008. Variation in offspring quality with cavity orientation in the great tit. Ethol. Ecol. Evol. 20: 375–389. doi: 10.1080/08927014.2008.9522518
- Hadley, N.F. 1969. Microenvironmental factors influencing the nesting sites of some subalpine fringillid birds in Colorado. Arct. Alp. Res. 1: 121–126. doi: 10.2307/1550018
- Holder, K. & Montgomerie, R. 1993. Rock ptarmigan (Lagopus mutus). In Poole, A. & Gill, F. (eds) The Birds of North America 51, 1–24. Academy of Natural Sciences, Philadelphia and American Ornithologists Union, Washington, DC.
- Kang, C.-K., Moon, J.-Y., Lee, S.-I. & Jablonski, P.G. 2012. Camouflage through an active choice of a resting spot and body orientation in moths. J. Evolution. Biol. 25: 1695–1702. doi: 10.1111/j.1420-9101.2012.02557.x
- Kovach, W.L. 2010. Oriana – Circular Statistics for Windows, Ver. 3. Kovach Computing Services, Pentraeth. http://www.kovcomp.com/oriana
- Lustik, S., Talbot, S. & Fox, E.L. 1970. Absorption of radiant energy in Red-winged Blackbirds Agelaius phoeniceus. The Condor 72: 471–473. doi: 10.2307/1366396
- Macdonald, S.D. 1970. The breeding behaviour of the Rock Ptarmigan. Living Bird 9: 195–238.
- Nelson, K.J. & Martin, K. 1999. Thermal aspects of nest-site location for vesper sparrows and horned larks in British Columbia. Studies in Avian Biol. 19: 137–143.
- Norment, C.J. & Green, K. 2004. Breeding ecology of Richard's Pipit Anthus novaeseelandiae in the Snowy Mountains. Emu 104: 327–336. doi: 10.1071/MU04006
- Rae, S. & Rae, D. 2013. Orientation of tawny frogmouth Podargus strigoides nests and their position on branches optimises thermoregulation and cryptic concealment. Aust. J. Zool. 61: 469–474. doi: 10.1071/ZO13090
- Rauter, C., Reyer, H.-U. & Bollmann, K. 2002. Selection through predation, snowfall, and microclimate on nest-site preferences in the water pipit Anthus spinoletta. Ibis 144: 433–444. doi: 10.1046/j.1474-919X.2002.00013.x
- Sargent, T.D. 1966. Background selections of Geometrid and Noctuid moths. Science 154: 1674–1675. doi: 10.1126/science.154.3757.1674
- Sawa, Y., Takeuchi, Y. & Nakamura, H. 2011. Nest site selection and nesting biology of Rock Ptarmigan Lagopus muta japonicus in Japan. Bird Study 58: 200–207. doi: 10.1080/00063657.2011.559532
- Scherini, G.C., Tosi, G. & Wauters, L.A. 2003. Social behaviour, reproductive biology and breeding success of Alpine Rock Ptarmigan Lagopus mutus helveticus in northern Italy. Ardea 91: 11–23.
- Steen, J.B. & Unander, S. 1985. Breeding biology of the Svalbard Rock Ptarmigan Lagopus mutus hyperboreus. Ornis. Scand. 16: 198–204. doi: 10.2307/3676631
- Walsberg, G.E. 1985. Physiological consequences of microhabitat selection. In Cody, M.L. (ed.) Habitat Selection in Birds, 389–413. Academic Press, New York.
- Walsberg, G.E. 1986. Thermal consequences of roost-site selection: the relative importance of three modes of heat conservation. The Auk 103: 1–7. doi: 10.2307/4087224
- Watson, A. 1972. The behaviour of Ptarmigan. Br. Birds. 65: 6–26, 93–117.
- Watson, A. & Moss, R. 2008. Grouse. HarperCollins, London.
- Watson, A., Moss, R. & Rae, S. 1998. Population dynamics of Scottish Rock Ptarmigan cycles. Ecology 79: 1174–1192. doi: 10.1890/0012-9658(1998)079[1174:PDOSRP]2.0.CO;2
- Watson, A. & Rae, S. 1993. Ptarmigan. In Gibbons, D.W., Reid, J.B. & Chapman, R.A. (eds) The New Atlas of Breeding Birds, 128–129. T. & A.D. Poyser, London.
- Webb, D.R. 1987. Thermal tolerance of avian embryos: a review. The Condor 89: 874–898. doi: 10.2307/1368537
- Wiebe, K.L. 2001. Microclimate of tree cavity nests: is it important for reproductive success in Northern Flickers? The Auk 118: 412–421. doi: 10.1642/0004-8038(2001)118[0412:MOTCNI]2.0.CO;2
- Wiebe, K.L. & Martin, K. 1997. Effects of predation, body condition and temperature on incubation rhythms of white-tailed ptarmigan. Wildlife Biology 3: 143–151.
- Wilson, S. & Martin, K. 2008. Breeding habitat selection of sympatric White-tailed, Rock and Willow Ptarmigan in the southern Yukon Territory. J. Ornithol. 149: 629–637. doi: 10.1007/s10336-008-0308-8
- With, K.A. & Webb, D.R. 1993. Microclimate of ground nests: the relative importance of radiative cover and wind breaks for three grassland species. The Condor 95: 401–413. doi: 10.2307/1369363
- Wolf, B.O. & Walsberg, G.E. 1996. Thermal effects of radiation and wind on a small bird and implications for microsite selection. Ecology 77: 2228–2236. doi: 10.2307/2265716