
Abstract
Capsule Maximum provisioning rates occurred during the seasonal peak in caterpillar availability, controlling for brood age and time of day.
Aims To determine if the seasonal availability of caterpillars influenced Wood Warbler Phylloscopus sibilatrix nest provisioning rates.
Methods Remote cameras were used to monitor nest visits at 14 nests in 8 British oak Quercus woodlands during 2013. The timing of the caterpillar peak was estimated by monitoring the production of caterpillar faecal pellets under oak trees.
Results Maximum provisioning rates occurred when the date of provisioning coincided with the date of peak caterpillar availability, but the percentage change in provisioning rates during asynchrony was dependent on brood age. Younger broods experienced a greater decline in provisioning rates than older broods during asynchrony. Provisioning rates peaked in the morning and declined during the day, but when caterpillar availability was low there was little within-day variation.
Conclusion Provisioning rates show seasonal variation that is correlated with caterpillar availability. Previous studies suggest that Wood Warbler reproductive fitness is independent of synchrony between the breeding cycle and the caterpillar peak, however, the implications for adult fitness are unknown and should be investigated further.
The widespread decline of Afro-Palearctic migrant bird populations during the past 30 years is widely recognized (Sanderson et al. Citation2006, Hayhow et al. Citation2014, Vickery et al. Citation2014), with woodland specialists and long-distance migrants showing the most severe declines (Gregory et al. Citation2007, Hewson & Noble Citation2009, Ockendon et al. Citation2012). Drivers of declines are generally unknown, with many potential causes hypothesized. One potential factor that may particularly affect long-distance migrants, due to constraints on the timing of key lifecycle stages, is a trophic mismatch between migrants' timing of breeding, and prey availability when nestling food requirements are highest. Woodland phenology is regulated by temperature, and warming of spring climate over recent decades has resulted in earlier phenology of tree budburst, invertebrates and to a lesser extent birds (Phillimore et al. Citation2013, Visser & Both Citation2005). The phenology mismatch hypothesis predicts that a temporal mismatch between the timing of peak resource availability and peak demand from consumers can have negative consequences for individuals and populations (Both et al. Citation2006, Both et al. Citation2009, Jones & Creswell Citation2010). However, studies investigating the relationship between trophic mismatch and Afro-Palearctic migrant declines have been limited to a small number of species (mainly flycatchers Ficedula), and a general link has yet to be established (Knudsen et al. Citation2011).
Many woodland birds feed caterpillars to their young during the breeding season (Naef-Daenzer & Keller Citation1999, Thomas et al. Citation2001, Both et al. Citation2006, Maziarz & Wesołowski Citation2010), but caterpillar availability shows a very short, pronounced seasonal peak (Veen et al. Citation2010, Smith et al. Citation2011). In parts of Northern Europe, the timing of the caterpillar peak has advanced due to localized spring warming over the last few decades (Both et al. Citation2006). In the Netherlands, Pied Flycatchers Ficedula hypoleuca failed to sufficiently advance the timing of the breeding cycle to track the advancing date of peak caterpillar availability, with negative consequences for reproductive fitness (Both et al. Citation2006). However, advancing spring temperatures can also have a positive impact on caterpillar-dependent species. For example, in Wytham Wood, UK, the proportion of breeding Great Tits Parus major that achieved synchrony with the caterpillar peak has increased in response to warming spring temperatures, resulting in greater reproductive output (Cresswell & McCleery Citation2003).
Wood Warblers Phylloscopus sibilatrix are long-distance migrants breeding in Europe and Central Asia, and wintering in Equatorial Africa. The European population declined by approximately 33% between 1980 and 2009, but the magnitude of declines varied regionally (Vickery et al. Citation2014), with the British population declining by 66% between 1995 and 2012 (Hayhow et al. Citation2014). Wood Warblers breed in natural and semi-natural broadleaved and coniferous woodland, with densities highest where Oak Quercus is dominant (Wesołowski Citation1985, Wesołowski & Maziarz Citation2009, Mallord et al. Citation2012a). Oak woodland supports higher volumes of caterpillar (Lepidopteran larvae) biomass than other woodland types (Burger et al. Citation2012, Veen et al. Citation2010), and longitudinal studies of Wood Warbler breeding ecology in Poland show that caterpillars form the largest proportion of nestling diet, followed by spiders and winged insects (Maziarz & Wesołowski Citation2010). Caterpillars are highly nutritious (Banjo et al. Citation2006), and typically have a large body mass relative to other invertebrates. This makes them nutritionally valuable for nestlings and can minimize the cost per unit effort of adult foraging rates.
Although Wood Warblers selectively exploit caterpillars when they are abundant (Maziarz & Wesołowski Citation2010), evidence from Poland suggests that populations do not synchronize the breeding cycle with the caterpillar peak, and that nestling condition and survival rates are independent of seasonal caterpillar availability (Maziarz & Wesołowski Citation2010). Nonetheless, it is likely that when caterpillars are scarce, adults compensate by adjusting their foraging and provisioning rates to meet the energetic requirements of nestlings. For example, Blue Tit Parus caeruleus foraging effort increases by up to 100% during periods of low caterpillar availability (Thomas et al. Citation2001). This can have fitness consequences for adults and nestlings, and is correlated with reduced reproductive fitness and survival rates in several woodland bird species (Naef-Daenzer & Keller Citation1999, Thomas et al. Citation2001, Tremblay et al. Citation2005, Both et al. Citation2006). In light of Wood Warbler population declines, it is therefore important to understand any potential effects of trophic mismatch on foraging and provisioning rates.
Here, we aimed to determine if Wood Warbler nest provisioning rates were correlated with the seasonal availability of caterpillars in British oak woodlands. We hypothesized that adults would make fewer nest visits per hour when caterpillars were abundant. This was based on the assumption that adults might increase the number of caterpillars per load during the peak, and fewer visits would therefore be required to satisfy the energetic requirements of nestlings.
METHODS
Study area
The study was conducted in the Loch Lomond & The Trossachs National Park in Central Scotland, and Dartmoor National Park, Devon, Southwest England, during the Wood Warbler breeding season between April and July 2013. Eight woodlands (three in Scotland and five in England) were surveyed for breeding Wood Warblers (, ). These were dominated by oak, with frequent Ash Fraxinus excelsior, Rowan Sorbus aucuparia, Birch Betula, Holly Ilex aquifolium and Alder Alnus.
Figure 1. Map of the study sites in Scotland and England. Rectangles on the inset maps show the location of the study areas in Great Britain. Shaded areas on the inset maps indicate the approximate range of Wood Warblers in Great Britain based on BirdLife International & Natureserve (Citation2011).
Table 1. Details of the study sites and total number of nests that made it to the nestling stage (total nests) in each woodland, as well as the number of nests used in the analysis (study nests). Estimated dates of peak caterpillar availability (Julian day) are also given. Site designations are also shown, with Site of Special Scientific Interest abbreviated as SSSI.
Nest monitoring
From mid-April, visits were made to woodlands at intervals of between one and five days to identify all breeding territories. The locations of singing or calling adults and evidence of breeding activity (e.g. nest building) were recorded using a GPS. Once located, nests were monitored for the presence of eggs at intervals of between three and four days to minimize disturbance. Once incubation was initiated, future visits were timed so that the hatch date could be accurately determined. We did not attempt to record first egg laying date.
Automated cameras (for details see Bolton et al. Citation2007, Stevens et al. Citation2008, Mallord et al. Citation2012b) were deployed at nests to record provisioning rates (n adult nest visits/hour: Temrin et al. Citation1997). Provisioning events were defined as an adult landing at the nest entrance and feeding young. Camera sensitivity was adjusted to capture images at 0.2-second intervals when movement at the nest was detected, ensuring all provisioning events were captured. To verify this, we manually recorded provisioning rates at a sample of nests with active cameras. A single observer carried out watches from a distance of approximately 15–20 m to minimize disturbance (in all instances normal behaviour, e.g. incubation or provisioning, resumed within five minutes of the observer's arrival). Only camera data recorded during daylight were analysed, and we expected camera sensitivity to be independent of time of day. Manual counts of provisioning rates were therefore made opportunistically during daylight, normally before changing camera batteries and memory cards in the morning. There was no significant difference between estimates of hourly provisioning rates obtained by direct observation or from camera data (Wilcoxon signed-rank test: V = 15, P = 0.40, n = 9 hours from three nests).
It was not possible to differentiate between adult males and females. No attempt was made to quantify beak load sizes or identity prey items brought to nests, since they were often too small to discern or obscured from view.
Fieldwork in Scotland was licensed by Scottish Natural Heritage (Bird Licence no. 19441). No license was required in England.
Estimating peak caterpillar availability
To estimate the date of peak caterpillar availability, we monitored frass fall in the study areas throughout the season using frass fall traps (Smith et al. Citation2011). A total of 26 traps (mean 4.33 ± 0.71 se, range 3–7 traps per woodland) were deployed beneath oak trees <1 km from nests. Traps were emptied at approximately weekly intervals (mean 6.71 ± 0.39 se, range 3–15 days per sampling period). For each woodland, we calculated the average daily frass weight per trap for the deployment period. Frass fall was highly peaked in time, and the peak was defined as the middle date of the deployment period with the highest average daily weight (Smith et al. Citation2011).
Data analysis
Several hundred hours of camera data were recorded, and it was not possible to analyse the entire data set within the duration of the project. We therefore obtained unbiased measures of provisioning rates using a re-sampling approach. A preliminary investigation showed that provisioning began after 06:00 hours and stopped before 21:00 hours. We expected within-day variation in provisioning rates, and divided a day into three five-hour blocks for resampling; from 06:00–11:00 hours, 11:00–16:00 hours and 16:00–21:00 hours. We then randomly selected one hour from each time block per day of camera data for transcription.
We used a generalized linear mixed model (GLMM) with a Poisson error structure to evaluate the influence of temporal and brood specific variables on provisioning rates. Relative caterpillar availability, our key fixed effect of interest, was calculated as the number of days' difference between the date of provisioning and the date of the local frass peak (synchrony), where a value of 0 days corresponded to the date of the peak. Seasonal caterpillar availability in oak woodlands shows a strong, curvilinear pattern (Smith et al. Citation2011), and we expected provisioning rates to reflect this. We therefore included the effect of synchrony as a fixed, continuous covariate as well as the quadratic term synchrony2. Wood Warbler nestling food requirements increase with age, and peak at approximately ten days old (Maziarz & Wesołowski Citation2010, Temrin et al. Citation1997). To account for this, two-way interactions were included between synchrony + synchrony2 and a fixed covariate of brood age (n days old on the date of provisioning). The first hatch date for the clutch was defined as 0 days old.
We expected provisioning rates to peak in the morning and decline during the day, and for each sub-sample of hourly provisioning rates, time of day was included as a fixed, continuous covariate. We hypothesized that when caterpillars were abundant, nestling energetic requirements would be met earlier in the day, and expected within-day variation in provisioning rates to vary as a function of caterpillar availability. This was explored by including a two-way interaction between time of day and synchrony + synchrony2.
A random intercept of nest, hierarchically nested within site, accounted for repeat observations of hourly provisioning rates at the same nest, non-independence of nest location and potential differences in average caterpillar abundance between sites. Brood size (n nestlings) was included as an offset to account for the expectation that provisioning rates would vary proportionally with the number of nestlings.
The measure of synchrony was highly correlated with Julian day (i.e. season: rs = 0.76, P < 0.001) and because of this the general effect of season was likely to be indistinguishable from the effect of synchrony. Since our a priori assumption was that any seasonal effect on provisioning rates would operate through food availability (i.e. caterpillar availability), the effect of season (i.e. separate to caterpillar availability) was not explored further. However, for comparison, we present the ΔAICc values for the global model described above and the alternative where Julian day + Julian day2 replace synchrony + synchrony2.
We used an information theoretic approach (Burnham & Anderson Citation2002) to select the best combination of fixed effects from the global model. We first generated a full set of candidate models with all possible fixed effect combinations (excluding additional interactions to prevent over-parameterization) and corresponding Akaike information criterion values corrected for small sample size (AICc). Interactions and quadratic terms were not considered without their constituent main effects. Models in the set with ΔAICc < 4 were considered equally parsimonious, and a final model was constructed by averaging the coefficients of all models below this threshold (Burnham & Anderson Citation2002, Bolker et al. Citation2009, Burnham et al. Citation2011). This approach was considered appropriate since the global model had relatively few parameters, and fully satisfied our a priori hypotheses (Anderson & Burnham Citation2002, Stephens et al. Citation2005). Averaged coefficients were calculated using the conditional method, where coefficients are averaged across only those models where the parameter of interest is present (Burnham & Anderson Citation2002).
Model predictions and 95% confidence intervals (CI) for the parameters were made using averaged coefficients, with non-focal predictors set at their median values. Predicted values are given for the median brood size only. R statistical software was used for analysis (R Core Team Citation2014), the GLMM was implemented using the lme4 package (Bates et al. Citation2014) and the MuMIn package (Barton Citation2013) was used for model selection and averaging.
RESULTS
Camera data were available for 29 broods (Scotland n = 7; England n = 22), but surrounding vegetation constrained camera position and image quality varied between nests. We therefore inspected images prior to analysis and data were excluded if provisioning events were unidentifiable. This was done with no prior knowledge of brood synchrony or other nest variables, preventing bias. After inspection, data from 14 nests (Scotland n = 6, England n = 8) were selected for analysis (). The mean number of camera days per nest was 4 (± 0.77 se, range 1 − 10 days per nest) and, after re-sampling, 174 hours of camera data were analysed (mean = 12 ± 2 se, range 3–30 hours per nest) from 3 June to 13 July 2013. This covered 41 days of the season, totalling 58 nest days.
Brood size ranged between four and seven (mean = 6, 0.20 se, n = 14 nests) and the mean nestling age per day of camera data was seven days old (± 0.47 se, range 1–13 days old, n = 58 nest days). There was no correlation between nest hatch dates and the timing of the caterpillar peak (r = −0.34, P = 0.2, n = 14 nests).
The estimated dates of peak frass fall for individual study sites are shown in . The mean difference between the timing of the frass peak and the day of provisioning (synchrony) was eight days (2 se, range −18–36 days, n = 58 nest days). Although we did not systematically identify prey items, caterpillars, spiders (Aranae) and winged insects (e.g., Tipulidae) were identified opportunistically.
Provisioning rates
Hourly provisioning rates ranged from 0–38 visits per hour (mean = 14 ± 1 se, n = 174 hours). Twenty-one models in the full model set (n = 38 models; Table S1) had ΔAICc values of <4, and are shown in . The top ranking model contained only the effects of brood age and time of day. Based on AICc weights, this model was 2.8 times more strongly supported than the global model (evidence-based ratio = wi /wj = 0.14/0.05 = 2.8). The null model had no support (wi < 0.0001). The global model was 3.96 ΔAICc points higher than the alternative where Julian day replaced synchrony.
Table 2. Model selection summary: 21 GLMMs had ΔAICc values of <4 (full set n = 38 models, Table S1 Supplementary Online Material). Parameters in the model were brood age (Age), synchrony with the caterpillar peak (Sync and Sync2) and time of day (Time). Models are ranked by AICc weight (wi); where higher weighted models have more support.
Model-averaged parameter estimates () showed that maximum provisioning rates occurred between −3 and 0 days synchrony (a), but the percentage change in provisioning rates during asynchrony was dependent on brood age. To illustrate this interaction, a shows the model-averaged predictions of the relationship between provisioning rates and synchrony at ages 1 and 10 days old, respectively (solid line ± CI). In one-day-old broods, mean provisioning rates declined by 40.5% from 8.4 visits per hour (5.7–11 CI) during the frass peak (i.e. 0 days synchrony) to 5 visits per hour (1.6–8.5 CI) 30 days after the peak. In ten-day-old broods, mean provisioning rates declined by 22.4% from 17.4 visits per hour (13–21.8 CI) during the frass peak to 13.5 visits per hour (8.1–18.9 CI) 30 days after the peak.
Figure 2. Observed values (points) and predicted values (lines ± 95% CI) from the GLMM analysis of hourly provisioning rates. Panel (a) illustrates the effect of the interaction between synchrony and brood age on provisioning rates, where synchrony of 0 days corresponds to the date of the frass peak. Conditional averaged model predictions were made at the focal ages of 1 day (solid line) and 10 days (dashed line), with non-focal predictors set at their median values (time: 12:00 hours; offset (brood size): n = 6). Open circles show the observed values for age 1 day, filled squares the observed values for age 10 days, and filled circles all other observed values. Panel (b) shows the effect of the interaction between synchrony and time of day on provisioning rates. Predictions were made at 0 days synchrony (dashed line) and +30 days synchrony (solid line), and non-focal predictors set at median values (brood age: 7 days; offset (brood size): n = 6). Filled circles show observed values ≤ 15 days synchrony, and open circles > 15 days synchrony to illustrate the pattern in the observed data.
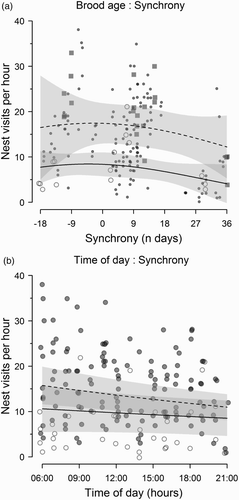
Table 3. Conditional model-averaged parameter estimates for the GLMM analysis ± 95% CI. Coefficients are back transformed. Relative variable importance (RVI) is also given.
Model-averaged predictions showed that, during the frass peak, mean provisioning rates peaked in the morning and declined linearly during the day (b). This became less apparent as the date of provisioning and the frass peak became asynchronous, and 30 days after the peak there was little within-day variation (b).
DISCUSSION
We expected provisioning rates to be lower when broods were synchronized with the seasonal caterpillar peak. However, in contrast to expectations, provisioning rates were lower when broods were asynchronous with the peak. The decline in provisioning rates during asynchrony can probably be explained by adults spending longer periods of time foraging for sufficient quantities of prey, although we did not quantify this during the study.
The most important predictors of provisioning rates in our model were brood age and time of day, and average hourly provisioning rates given by Temrin et al. (Citation1997) for any given age agree closely with our results. In contrast, Temrin et al. (Citation1997) found no effect of time of day. This contradictory result may be related to local conditions, such as differences in actual caterpillar abundance or the species composition of the prey community.
In our model, the interaction between brood age and synchrony showed that the percentage decline in provisioning rates for asynchronous broods was greatest during the early stages of nestling development. A possible explanation is that nestling diet diversifies with age (Temrin et al. Citation1997, Maziarz & Wesołowski Citation2010), and younger broods are therefore less able to accept alternative prey when caterpillar availability is low. As a result, adults may be forced to spend longer periods searching for caterpillars in order to feed younger broods in nests timed before or after the caterpillar peak. This explanation is supported by evidence from Poland, which demonstrated that the proportion of caterpillars in the diet can remain high when frass fall is below detectable levels (Maziarz & Wesołowski Citation2010), suggesting that adults actively seek caterpillars even when availability is low.
During the caterpillar peak, provisioning rates were strongly peaked in the morning, but the interaction between synchrony and time of day showed that there was little within-day variation when broods were asynchronous. This could be due to nestlings becoming gorged in the morning when caterpillars are readily available, and energetic requirements are therefore met earlier in the day, but this is speculative and requires further study.
If provisioning rates are correlated with caterpillar availability, do Wood Warblers also experience a fitness cost when mismatch occurs? Previous studies suggest that Wood Warbler reproductive fitness is unaffected by trophic mismatch. For example, in Poland, populations did not synchronize the breeding cycle with the caterpillar peak during a 30-year period, despite the importance of caterpillars in the diet of nestlings (Wesołowski & Maziarz Citation2009, Maziarz & Wesołowski Citation2010). Likewise, we found no relationship between hatching date and the timing of the caterpillar peak. Furthermore, nestling body condition and survival rates are also independent of caterpillar availability (Maziarz & Wesołowski Citation2010). However, the implications for adults are unknown. In other woodland passerines, changes in provisioning or foraging rates during trophic mismatch are correlated with reduced adult and nestling fitness (Naef-Daenzer & Keller Citation1999, Thomas et al. Citation2001, Tremblay et al. Citation2005). For example, in Blue Tits, adult survival in the population declines significantly following trophic mismatch (Thomas et al. Citation2001), with potentially negative consequences for population viability. If the decline in Wood Warbler provisioning rates during asynchrony is associated with adults flying longer distances from the nest to search for prey, this may have similarly negative consequences for adult survival rates, and requires further investigation.
Two caveats have to be highlighted when interpreting our findings. Firstly, since we did not measure beak load size during the study or measure nestling growth rates, it is impossible to determine if the decline in provisioning rates during asynchrony resulted in broods receiving less food. Secondly, our study was restricted to a single season, and further studies in the same area would be necessary to allow proper generalization of our findings to other years. With these caveats in mind, we conclude that provisioning rates are correlated with seasonal caterpillar availability, but that this relationship depends on brood age. However, on the basis of our results alone we cannot make inferences about possible fitness consequences, and in light of Wood Warbler population declines, we suggest that future studies investigate the effects of trophic mismatch on adult fitness. This will be logistically challenging, as Wood Warblers exhibit low inter-annual site fidelity (Temrin et al. Citation1997, Wesołowski & Maziarz Citation2009), making it difficult to monitor adult survival rates between years.
Supplemental data
Table S1 showing all models considered during model selection can be accessed at 10.1080/00063657.2015.1034086.
Supplementary material
Download MS Word (95 KB)ACKNOWLEDGEMENTS
The authors would like to thank Ken and Linda Smith, Andrew Cristinacce, Chris Orsman and Kiri Stone for assistance with data collection.
ORCID
Malcolm D. Burgess http://orcid.org/0000-0003-1288-1231
REFERENCES
- Anderson, D.R. & Burnham, K.P. 2002. Avoiding pitfalls when using information-theoretic methods. J. Wildlife Manage. 66: 912–918.
- Banjo, A.D., Lawal, O.A. & Songonuga, E.A. 2006. The nutritional value of fourteen species of edible insects in southwestern Nigeria. Afr. J. Biotechnol. 5: 298–301.
- Barton, K. 2013. MuMIn: Multi-model inference. R package version 1.9.5. [online] http://CRAN.R-project.org/package=MuMIn [accessed 21 April 2014].
- Bates, D., Maechler, M., Bolker, B. & Walker, S. 2014. lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1-5. [online] http://CRAN.R project.org/package=lme4 [accessed 21 April 2014].
- BirdLife International & Natureserve. 2011. Phylloscopus sibilatrix. Bird species distribution maps of the world. [online] http://www.iucnredlist.org. [accessed 18th March 2014].
- Bolker, B.M., Brooks, M.E., Clark, C.J., Geange, S.W., Poulsen, J.R., Stevens, M.H.H. & White, J.S.S. 2009. Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol. 24: 127–135.
- Bolton, M., Butcher, N., Sharpe, F., Stevens, D. & Fisher, G. 2007. Remote monitoring of nests using digital camera technology. J. Field Ornithol 78: 213–220.
- Both, C., Bouwhuis, S., Lessells, C.M. & Visser, M.E. 2006. Climate change and population declines in a long-distance migratory bird. Nature 441: 81–83.
- Both, C., Van Turnhout, C.A., Bijlsma, R.G., Siepel, H., Van Strien, A.J. & Foppen, R.P. 2009. Avian population consequences of climate change are most severe for long-distance migrants in seasonal habitats. Proc. R. Soc B Biol. Sci. doi:10.1098/rspb.2009.1525
- Burger, C., Belskii, E., Eeva, T., Laaksonen, T., Mägi, M., Mänd, R., Qvarnström, A., Slagsvold, T., Veen, T., Visser, M.E., Wiebe, K.L., Wiley, C., Wright, J. & Both, C. 2012. Climate change, breeding date and nestling diet: how temperature differentially affects seasonal changes in pied flycatcher diet depending on habitat variation. J. Anim. Ecol. 81: 926–936.
- Burnham, K.P. & Anderson, D.R. 2002. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. Springer, London.
- Burnham, K.P., Anderson, D.R. & Huyvaert, K.P. 2011. AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav. Ecol. Sociobiol. 65: 23–35.
- Cresswell, W. & McCleery, R. 2003. How great tits maintain synchronization of their hatch date with food supply in response to long-term variability in temperature. J. Anim. Ecol. 72: 356–366.
- Gregory, R.D., Vorisek, P., Van Strien, A., Gmelig Meyling, A.W., Jiguet, F., Fornasari, L., Reif, J., Chylarecki, P. & Burfield, I. 2007. Population trends of widespread woodland birds in Europe. Ibis 149: 78–97.
- Hayhow, D.B., Conway, G., Eaton, M.A., Grice, P.V., Hall, C., Holt, C.A., Kuepfers, A., Noble, D.G., Oppel, S., Risely, K., Stringer, C., Stroud, D.A., Wilkinson, N. & Wotton, S. 2014. The State of the UK's Birds 2014. RSPB, BTO, WWT, JNCC, NE, NIEA, NRW and SNH, Sandy.
- Hewson, C.M. & Noble, D.G. 2009. Population trends of breeding birds in British woodlands over a 32-year period: relationships with food, habitat use and migratory behaviour. Ibis 151: 464–486.
- Jones, T. & Cresswell, W. 2010. The phenology mismatch hypothesis: are declines of migrant birds linked to uneven global climate change? J. Anim. Ecol. 79: 98–108.
- Knudsen, E., Lindén, A., Both, C., Jonzén, N., Pulido, F., Saino, N., Sutherland, W.J., Bach, L.A., Coppack, T., Ergon, T., Gienap, P., Gill, J.A., Gordo, O., Hedenström, A., Lehikoinen, E., Marra, P.P., Møller, A.P., Nilsson, A.L.K., Péron, G., Ranta, E., Rubolini, D., Sparks, T.H., Spina, F., Studds, C.E., Sæther, S.A., Tryjanowski, P. & Stenseth, N.C. 2011. Challenging claims in the study of migratory birds and climate change. Biol. Rev. 86: 928–946.
- Mallord, J.W., Charman, E.C., Cristinacce, A. & Orsman, C.J. 2012a. Habitat associations of Wood Warblers Phylloscopus sibilatrix breeding in Welsh oakwoods. Bird Study 59: 403–415.
- Mallord, J.W., Orsman, C.J., Cristinacce, A., Butcher, N., Stowe, T.J. & Charman, E.C. 2012b. Mortality of Wood Warbler Phylloscopus sibilatrix nests in Welsh Oakwoods: predation rates and the identification of nest predators using miniature nest cameras. Bird Study 59: 286–295.
- Maziarz, M. & Wesołowski, T. 2010. Timing of breeding and nestling diet of Wood Warbler Phylloscopus sibilatrix in relation to changing food supply. Bird Study 57: 540–552.
- Naef-Daenzer, B. & Keller, L.F. 1999. The foraging performance of great and blue tits (Parus major and P. caeruleus) in relation to caterpillar development, and its consequences for nestling growth and fledging weight. J. Anim. Ecol. 68: 708–718.
- Ockendon, N., Hewson, C.M., Johnston, A. & Atkinson, P.W. 2012. Declines in British-breeding populations of Afro-Palaearctic migrant birds are linked to bioclimatic wintering zone in Africa, possibly via constraints on arrival time advancement. Bird Study 59: 111–125.
- Phillimore, A.B., Proios, K., O'Mahony, N., Bernard, R., Lord, A.M., Atkinson, S. & Smithers, R.J. 2013. Inferring local processes from macro-scale phenological pattern: a comparison of two methods. J. Ecol. 101: 774–783.
- R Core Team. 2014. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [online] http://www.R-project.org/ [accessed 21 April 2014].
- Sanderson, F.J., Donald, P.F., Pain, D.J., Burfield, I.J. & Van Bommel, F.P. 2006. Long-term population declines in Afro-Palearctic migrant birds. Biol. Conserv. 131: 93–105.
- Smith, K.W., Smith, L., Charman, E., Briggs, K., Burgess, M., Dennis, C., Harding, M., Isheroowd, C., Isherwood, I. & Mallord, J. 2011. Large-scale variation in the temporal patterns of the frass fall of defoliating caterpillars in oak woodlands in Britain: implications for nesting woodland birds. Bird Study 58: 506–511.
- Stephens, P.A., Buskirk, S.W., Hayward, G.D. & Martinez del Rio, C. 2005. Information theory and hypothesis testing: a call for pluralism. J. Appl. Ecol. 42: 4–12.
- Stevens, D.K., Anderson, G.Q.A., Grice, P.V., Norris, K. & Butcher, N. 2008. Predators of Spotted Flycatcher Muscicapa striata nests in southern England as determined by digital nest-cameras. Bird Study 55: 179–187.
- Temrin, H., Brodin, A., Åkerström, O. & Stenius, S. 1997. Parental investment in monogamous pairs of Wood Warblers (Phylloscopus sibilatrix). J. Ornithol. 138: 93–101.
- Thomas, D.W., Blondel, J., Perret, P., Lambrechts, M.M. & Speakman, J.R. 2001. Energetic and fitness costs of mismatching resource supply and demand in seasonally breeding birds. Science 291: 2598–2600.
- Tremblay, I., Thomas, D., Blondel, J., Perret, P. & Lambrechts, M.M. 2005. The effect of habitat quality on foraging patterns, provisioning rate and nestling growth in Corsican blue tits Parus caeruleus. Ibis 147: 17–24.
- Veen, T., Sheldon, B.C., Weissing, F.J., Visser, M.E., Qvarnström, A. & Sætre, G.P. 2010. Temporal differences in food abundance promote coexistence between two congeneric passerines. Behav. Ecol. 162: 873–884.
- Vickery, J.A., Ewing, S.R., Smith, K.W., Pain, D.J., Bairlein, F., Škorpilova, J. & Gregory, R.D. 2014. The decline of Afro-Palearctic migrants and an assessment of potential causes. Ibis 156: 1–22.
- Visser, M.E. & Both, C. 2005. Shifts in phenology due to global climate change: the need for a yardstick. Proc. R. Soc. B Biol. Sci. 272: 2561–2569.
- Wesołowski, T. 1985. The breeding ecology of the Wood Warbler Phylloscopus sibilatrix in primaeval forest. Ornis Scand. 16: 49–60.
- Wesołowski, T. & Maziarz, M. 2009. Changes in breeding phenology and performance of Wood Warblers Phylloscopus sibilatrix in a primeval forest: a thirty-year perspective. Acta Ornithol. 44: 69–80.