
Abstract
Capsule Non-fish prey constitutes an important component of the diet of many fish-eating birds.
Aims In the present study, the role of non-fish prey in the diet of the Common Kingfisher Alcedo atthis was evaluated.
Methods The species and size spectrum of prey in the diet was studied at 15 nest sites on 6 trout streams, 1 river and 1 reservoir in the Czech Republic, using the analysis of the nest sediment.
Results 16 933 individual prey items were identified (99.93% fish and 0.07% non-fish prey). European Perch Perca fluviatilis, Roach Rutilus rutilus and Bleak Alburnus alburnus dominated the diets on the reservoir; Gudgeon Gobio gobio, European Chub Squalius cephalus and Roach those on the river; and Gudgeon, European Chub, Bullhead Cottus gobio, Roach, Bleak and Brown Trout Salmo trutta m. fario those in the trout streams. The sizes of their fish prey ranged from 16 to 134 mm in total length (LT) with an average size of 66 mm. The remains of non-fish prey were detected in only 5 of 30 nest sediments. The non-fish prey were mostly composed of large aquatic insect larvae: dragonflies Anax sp. and Aeshna sp., Common Club-tail Gomphus vulgatissimus and Great Diving Beetle Dytiscus marginalis. Kingfishers also took Spiny-cheek Crayfish Orconectes limosus, Newt Triturus sp. and a Lizard Lacerta sp. The estimated sizes of the non-fish prey ranged from 30 to 90 mm.
Conclusion The catch of non-fish prey appears to be accidental, and is more likely a result of target misinterpretation (fish-like body and fish-like movement) than a Kingfisher regularly switching to prey other than fish. The unique finding of a Lizard is the first record of an amniotic vertebrate in the diet of Common Kingfisher.
Fish-eating birds could be divided into several categories corresponding to the usual role of fish prey in their diet. The first category is represented by species such as Grey Heron Ardea cinerea or Night Heron Nycticorax nycticorax, in which the diet composition varies considerably according to different habitats and seasons and although fish can be a significant food source they may also disappear completely from the diet spectrum (Peris et al. Citation1995, Lekuona Citation2002, Rodríguez et al. Citation2007, Jakubas & Manikowska Citation2011 for Grey Heron; Kazantzidis & Goutner Citation2005, Hall & Kress Citation2008, Montesinos et al. Citation2008, Quiroga et al. Citation2013 for Night Heron). The second category is represented by species such as the Black Stork Ciconia nigra or Great Crested Grebe Podiceps cristatus, in which fish comprise the majority of the diet but the non-fish prey still occur as a constant component (usually <10% of the diet; Hampl et al. Citation2005 for Black Stork; Martinoli et al. Citation2003, Gagliardi et al. Citation2007 for Great Crested Grebe). The last category comprises species which are exclusive fish-eaters, such as the Great Cormorant Phalacrocorax carbo and the Common Kingfisher Alcedo atthis (Suter Citation1997, Keller Citation1998, Čech et al. Citation2008, Gwiazda & Amirowicz Citation2010, Emmrich & Düttmann Citation2011 for Great Cormorant; Raven Citation1986, Reynolds & Hinge Citation1996, Campos et al. Citation2000, Vilches et al. Citation2012, Čech & Čech Citation2013 for Common Kingfisher). However, especially in the case of the Common Kingfisher, most studies dealing with their diet have focused on only one or two streams/rivers (Campos et al. Citation2000, Čech & Čech Citation2013, Vilches et al. Citation2013), or the number of identified prey individuals was rather limited (Raven Citation1986, Reynolds & Hinge Citation1996). This makes it difficult to evaluate the real role of non-fish prey in the diet.
In this study, the diet of Common Kingfishers was analysed from 15 nest sites on 6 streams, 1 river and 1 reservoir. This represents the largest and most complex data set on the diet of Kingfishers ever published (Hallet Citation1977, Raven Citation1986, Campos et al. Citation2000, Vilches et al. Citation2012, Citation2013).
This study is part of a long-term ALCEDO project of the Czech Union for Nature Conservation (running from 1994) focusing on the biology and conservation of Kingfishers. The main aim is to quantify the dietary range of the Common Kingfisher in relation to the type of water body where they breed, and to quantify the prey species range, size distribution and the presence of non-fish prey.
MATERIALS AND METHODS
Study area
The study was carried out on Slapy Reservoir, Blanice River and on various trout streams (all within the Vltava River basin) in the southern part of Central Bohemia, Czech Republic (, ). All but one trout stream have been characterized as mesotrophic, Slapy Reservoir as meso- to eutrophic and Blanice River and Chotýšanka stream as eutrophic (Vlček et al. Citation1984, Čech & Čech Citation2006). The nest sites of the Common Kingfishers were situated at altitudes from 272 (Slapy) to 430 (Trnava) m.a.s.l.
Figure 1. A map of (a) Slapy Reservoir, (b) Blanice River and various trout streams in the south part of Central Bohemia and their locations in the Czech Republic. Grey dots indicate nest sites of Kingfishers Alcedo atthis where the mass of regurgitated pellets (nest sediment) was analysed. Numbers in parentheses indicate the year of sampling. Data published in Čech & Čech (Citation2013) are in italic (n = 2; Blanice River).
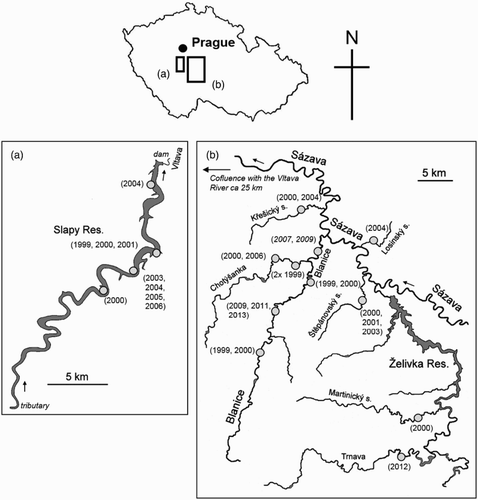
Table 1. The basic characteristics of Slapy Reservoir, Blanice River and various trout streams in the south part of Central Bohemia (according to Vlček et al. Citation1984) and the locations of the Kingfisher Alcedo atthis nest sites (n = 15) from which the mass of regurgitated pellets (nest sediment) was analysed. Nest site locations on the Blanice River and Chotýšanka stream are listed in order the mouth of the river/stream to the spring (source), and for Slapy Reservoir from the dam to the inflowing river (tributary), i.e. both against the flow (cf. also ).
Sampling
The species composition and sizes of prey in the diets of the Kingfishers were investigated from the mass of regurgitated pellets (nest sediment) collected from their nest tunnels and chambers during the years 1999–2013. Altogether, 9 nest sediments were analysed from Slapy Reservoir, 9 from Blanice River and 12 from the various trout streams (). No nest was occupied by the same pair of Common Kingfishers twice (P. Čech, pers. obs.).
All the regurgitated material from each individual nest was immersed for one week in a concentrated detergent solution, then washed through a sieve (mesh size 1 mm), dried at room temperature and analysed under a binocular microscope (magnification 8 times and 16 times).
To identify the species and sizes of the fish preyed upon, a reference collection of diagnostic bones was constructed for each of the potential prey species (see Čech et al. Citation2008, Čech & Vejřík Citation2011, Čech & Čech Citation2013). For dissection of the bones, fish (n = 731) were taken using gill nets, seine nets, dip nets and fishing rod catches from various streams, rivers, reservoirs and ponds belonging to the Vltava River basin during the years 2000–12. All fish were measured (total length, LT; to the nearest 1 mm), boiled, dissected and the diagnostic bones, selected according to Hallet (Citation1977, Citation1982), Reynolds & Hinge (Citation1996), Čech et al. (Citation2008) and Čech & Vejřík (Citation2011), were measured to the nearest 0.1 mm. Pharyngeal bones os pharyngeum were selected for Cyprinidae and Stone Loach Barbatula barbatula; lower jaws dentale for Percidae, Salmonidae and for Northern Pike Esox lucius; upper jaws maxillare for salmonid species; and preopercular bones praeoperculare for Bullhead Cottus gobio and percid species. Overall, 31 304 diagnostic bones were examined in this study. The measurements selected were the pharyngeal bone tip for cyprinid species, dental length for percid and salmonid species and for Northern Pike, maxilar length for salmonid species, preopercular length for Bullhead and the preopercular gape for percid species (for details see Čech et al. Citation2008, Čech & Vejřík Citation2011). For each species from the reference collection, a linear regression established the relationship between the measured dimension of the diagnostic bone and the fish total length (Čech et al. Citation2008, Čech & Vejřík Citation2011, Čech & Čech Citation2013). In the case of the Stone Loach, the linear regression relationship was taken from Reynolds & Hinge (Citation1996). For the Grayling Thymallus thymallus, the polynomial regression of standard length (LS) on dental length (dimension ‘dentary 1’) was taken from the work of Hájková et al. (Citation2003) and recalculated to total length using the equation: LT = LS × 1.14.
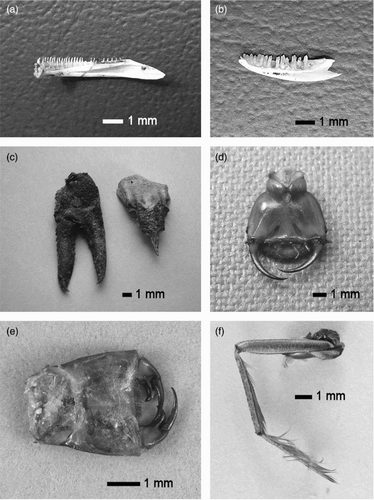
The species and size of non-fish prey in the diet of the Kingfishers were identified from the remains of head capsules, mandibles mandibulae, masks labia, palpal lobes palpi labiale and legs using the key of water insect larvae (Rozkošný Citation1980), from head, body, wing and leg remains (terrestrial insects), from claws chelae, remains of rostrum and abdomen (crayfish Astacoidea) and from lower jaws (non-fish vertebrates) using the authors’ personal collection of diagnostic material (). In all cases, the species and estimated size of prey were validated by specialists from the University of Ostrava, Faculty of Science (dragonflies Odonata), The Natural History Museum (beetles Coleoptera), The Nature Conservation Agency of the Czech Republic (dragonflies Odonata), The Biology Centre of the Czech Academy of Sciences, Institute of Entomology (other insects) and The University of South Bohemia, Faculty of Science (non-fish vertebrates).
Figure 2. Examples of non-fish prey remains found in the diet of Kingfishers Alcedo atthis. (a) Newt Triturus sp. – lower jaw. (b) Lizard Lacerta sp. – lower jaw. (c) Spiny-cheek Crayfish Orconectes limosus – claw and fragment of rostrum. (d) Great Diving Beetle Dytiscus marginalis larva – head with large mandibles from ventral view. (e) Common Club-tail Gomphus vulgatissimus larva – labium with massive palpal lobes from the top view. (f) Water Boatman Corixa sp. – paddling third leg (found in an individual pellet of Kingfisher on 14 March 2014, Blanice River). Photo: M. Čech (a–d) and F. Weyda (e, f).
RESULTS
In total, 16 933 individual prey were identified in the 30 analysed nest sediments of Common Kingfishers hunting on Slapy Reservoir, Blanice River and on various trout streams in the south part of Central Bohemia. Only 12 of these individual prey items (numerically 0.07%) belong to the category of non-fish prey, mostly to the category of large water insect larvae (Aeshnidae, Gomphidae and Dytiscidae; ). An absolute majority of the diet (99.93%) was composed of fish, from 27 species and 6 families (Cyprinidae, Balitoridae, Esocidae, Salmonidae, Cottidae and Percidae; ). The size of the fish prey ranged from 16 mm LT (European Chub Squalius cephalus) to 134 mm LT (Bleak Alburnus alburnus) with an average size of 66 mm LT (). Most of the fish prey (c 75%) taken was within a size range of 50–90 mm LT, and the optimum prey size represented fish between 60 and 80 mm LT ().
Figure 3. Frequency distributions of total length (LT) of all fish species found in the diets of Kingfishers Alcedo atthis hunting on Slapy Reservoir, Blanice River and on various trout streams in the Czech Republic, Central Bohemia, during the years 1999 and 2013 (cf. also ; 30 nest sediments, n = 16 921). Estimated sizes of individual non-fish prey (n = 12) are indicated by symbols: &, Spiny-cheek Crayfish Orconectes limosus; $, Hawkmoth Macroglossum stellatarum; #, Grasshopper Chorthippus parallelus; §, Common Club-tail Gomphus vulgatissimus larva; *, Dragonfly Anax sp., Aeshna sp. larva; ¶, Great Diving Beetle Dytiscus marginalis larva; †, Newt Triturus sp.; ‡, Lizard Lacerta sp. Black arrow indicates the mean LT of consumed fish (irrespective of species) and white arrow indicates the mean length of consumed non-fish prey (irrespective of species).
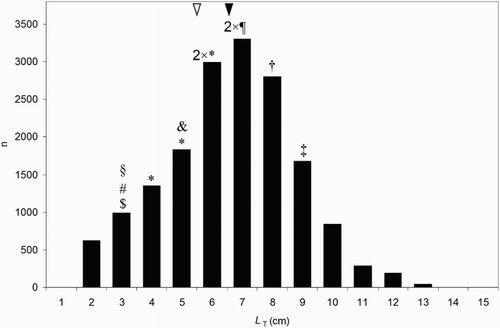
Table 2. Species composition of the diets of Common Kingfishers Alcedo atthis hunting on Slapy Reservoir, Blanice River and on various trout streams in the Czech Republic: A summary of data from 30 nest sediments collected in the years 1999–2013. n, number of individuals and %a, percentage of total abundance. The values of the three most numerous fish species are in bold.
In Slapy Reservoir, the diet of the Kingfishers was dominated by European Perch Perca fluviatilis (58.2% in numbers), followed by Roach Rutilus rutilus (16.9%) and Bleak (12.7%). In addition to the 18 fish species, non-fish prey were dragonfly Anax and Aeshna larvae, Great Diving Beetle Dytiscus marginalis larva, Grasshopper Chorthippus parallelus, Hummingbird Hawk-moth Macroglossum stellatarum and Spiny-cheek Crayfish Orconectes limosus (c for Spiny-cheek Crayfish, ). The estimated size of the non-fish prey was 30–70 mm (). The remains of non-fish prey were found in three out of nine nest sediments.
In Blanice River, the diet was dominated by Gudgeon Gobio gobio (35.1%), followed by European Chub (19.9%), Roach (15.8%) and Bleak (9.4%). Apart from 22 fish species, in 1 of 9 nest sediments non-fish prey were also found, namely Common Club-tail Gomphus vulgatissimus larva, Great Diving Beetle larva and Newt Triturus sp. (a, d, e, ). The estimated size of the non-fish prey was 30–80 mm ().
In the trout streams, the diet of the Kingfishers was dominated by Gudgeon (24.1%), followed by European Chub (18.6%), Bullhead (9.8%), Roach (9.3%), Bleak (9.1%) and Brown Trout Salmo trutta m. fario (8.1%). Apart from 25 fish species, only a Lizard Lacerta sp., of an estimated size of 90 mm, was detected in the diet of a Kingfisher (Losinský stream; b & 3, ). The non-fish prey was found only in 1 of 12 nest sediments.
DISCUSSION
Previous studies have demonstrated that the diet spectrum of the Common Kingfisher is relatively narrow, with only 5 (Reynolds & Hinge Citation1996, Campos et al. Citation2000) or between 9 and 11 (Raven Citation1986, Vilches et al. Citation2012, Citation2013) fish species identified in the diet. The most extensive study, performed on the Lesse River, in Belgium (Hallet Citation1977; 14 475 individual prey found), gave evidence for the Common Kingfisher hunting for 8 fish species, although Cyprinid fishes, which composed 56% of the diet in numbers, were not identified to species level.
In this study, between 18 and 25 fish species were found in the diets of the Common Kingfishers on individual types of water bodies (Slapy Reservoir, Blanice River and trout streams). However, only 1 (Gudgeon) out of 27 fish species was present in all analysed nest sediments, although Roach and Dace Leuciscus leuciscus were missing in only one (M. Čech, pers. data). The diet included fish species from all sections of the water body: sub-surface (European Chub, Bleak and Dace), mid-water (European Perch, Roach, Stone Moroko Pseudorasbora parva and Brown Trout) and benthic (Gudgeon, Bullhead and Stone Loach). In some regions of Europe, benthic species seem to be ignored by the Common Kingfisher even if they are abundant in the water body (e.g. Raven Citation1986 for Stone Loach and Bullhead; Vilches et al. Citation2012 for Pyrenean Stone Loach Barbatula quignardi and Ebro Barbel Luciobarbus graellsii).
The size ranges of fish prey (16–134 mm LT, optimum 60–80 mm LT; this study) slightly extended the range of fish taken by the Common Kingfisher stated in previous studies (Hallet Citation1977, Raven Citation1986, Reynolds & Hinge Citation1996, Campos et al. Citation2000, Vilches et al. Citation2012), especially in the case of maximum prey size. However, similarly to other fish-eating birds swallowing their prey whole, this upper size limit is clearly dependent on the body shape (i.e. species) of the fish (Čech et al. Citation2008, Čech & Čech Citation2011).
From the results presented above, it is evident that the Common Kingfisher is an almost exclusive fish-eater and that a catch of non-fish prey is rare. The remains of non-fish prey were detected in only 5 of 30 nest sediments analysed and, even in these samples, non-fish prey represented only 0.09–2.27% of the diet numerically (average 0.64%). This finding is in accordance with the results of other studies (Raven Citation1986, Reynolds & Hinge Citation1996, Campos et al. Citation2000, Vilches et al. Citation2012, Citation2013). A larger proportion of non-fish prey in the diet of the Kingfisher has been reported only by Iribarren & Nevado (Citation1982) from Spain where 96 bird stomachs, collected from 43 sites, contained (except for fish) 3.22% of Anura (by weight; those identified were frogs Rana sp.), 10.4% of White-clawed Crayfish Austropotamobius pallipes and 8.49% of insects (mostly Odonata and Coleoptera).
The non-fish remains, mostly tiny rove beetles Staphylinidae and small freshwater molluscs of the families Hydrobiidae, Planorbidae, Sphaeriidae, Lymnaeidae and Bithyniidae, were also detected in each of 12 Kingfisher nests on the Lesse River, Belgium (Hallet Citation1977). However, as the author correctly stated, rove beetles are more likely to be nest commensals, preying on various Diptera larvae in the nest sediment, than the real prey of Kingfisher and shells of molluscs came from the stomachs of benthivorous fish, such as Bullhead or Grayling, as a secondary prey. The only prey type which could be considered as a primary prey of Kingfisher seems to be large backswimmers Notonectidae and aquatic coleopterans (Hallet Citation1977). M. Čech and P. Čech (unpubl. data) found remains of Water Boatman Corixa sp. from the related family Corixidae in one individual pellet of a Kingfisher hunting on Blanice River (f). Beetles are mentioned as a prey of the Common Kingfisher in a summarizing work of Cramp (Citation1990), and larvae of the Great Diving Beetle have been found in the nest sediments of Kingfishers hunting on Blanice River and Slapy Reservoir (this study) as well as in two pellets of a Kingfisher hunting on Orlík Reservoir (Vltava River basin; M. Čech, P. Čech unpubl. data).
Large larvae of dragonflies were the most numerous non-fish prey in this study, and they are also listed as a potential prey by Cramp (Citation1990). In one individual pellet collected from Blanice River on 14 March 2014, M. Čech and P. Čech (unpubl. data) found diagnostic bones of one Gudgeon and three masks of Common Club-tail larvae and, similarly, in one pellet collected on 26 February 2010 on Slapy Reservoir, the same authors found bones of Bleak, Roach, two Perch and remains of a dragonfly larva.
Crayfish represent another type of prey occurring in the diet of Kingfisher (Iribarren & Nevado Citation1982, Cramp Citation1990). In the present study only one Spiny-cheek Crayfish was found, in a nest sediment from Slapy Reservoir. On this reservoir, the same species of invasive crayfish was found in the diet of another exclusive fish-eater, the Great Cormorant, representing 0.16% of the diet numerically (Čech Citation2012).
There is no doubt that the Kingfisher is a fish specialist. Also, we suggest that catching of dragonfly larvae, water beetle larvae, crayfish and newts (this study) is most likely a result of target misinterpretation (fish-like body and fish-like movement) than a real selection of non-fish prey. Common Kingfishers have been recorded diving for pieces of vegetation (Cramp Citation1990) or, more specifically, for willow leaves carried away by the stream (Čech & Čech Citation2011). Exceptionally, however, individual Common Kingfishers could temporarily switch to prey other than fish. For example, Vacher & Rufray (Citation2005) reported an observation of a Common Kingfisher taking larvae of the Common Spadefoot Toad Pelobates fuscus from a pond located near a forest edge in Alsace, France. Similarly, Čech & Čech (Citation2011) described the case of a young Common Kingfisher successfully hunting for Common Water Striders Gerris lacustris.
It is not known whether the occasional terrestrial insect (grasshopper, hawk-moth; this study) in the diet is a result of capture on land, or capture after the prey accidentally fell into the water (Čech & Čech Citation2013), but Kingfishers have occasionally been seen to hunt over grassland (Slapy Reservoir; P. Čech, pers. obs.).
Amphibians, snakes, lizards and even small birds and mammals are usual components of the diet of many other kingfisher species (Woodall Citation1991). However, the unique finding of a lower jaw from a 90 mm lizard in the nest sediment from Losinský stream is the first record of an amniotic vertebrate in the diet of Common Kingfisher (cf. Cramp Citation1990).
With the largest and most complex data set ever published (30 nest sediments analysed, 16 933 prey individuals found) the present study has shown that fish dominate the diet of Common Kingfishers (99.93% of the diet numerically). The taking of non-fish prey may be a result of target misinterpretation rather than a strategic switch to prey other than fish.
ACKNOWLEDGEMENTS
The authors thank M. Burgis for carefully reading and correcting the English and A. Dolný (University of Ostrava, Faculty of Science), L. Hanel (Nature Conservation Agency of the Czech Republic), T. Soldán (Biology Centre CAS, Institute of Entomology), P. Havelková, B. Hlivka (both University of South Bohemia, Faculty of Science) and V. Kubánˇ (Natural History Museum) for their help with the identification of non-fish prey. The authors thank F. Weyda for photos of non-fish prey remains.
Additional information
Funding
REFERENCES
- Campos, F., Fernández, A., Gutiérrez-Corchero, F., Martin-Santos, F. & Santos, P. 2000. Diet of the Eurasian kingfisher (Alcedo atthis) in northern Spain. Folia Zool. 49: 115–121.
- Čech, M. 2012. Diet of the great cormorant (Phalacrocorax carbo) in the Vltava River basin: a summary of results. Sylvia 48: 39–55 (in Czech with abstract and extended summary in English).
- Čech, M. & Čech, P. 2006. Diet of kingfisher at various types of waters. In Čech, P. (ed.) Kingfisher (Alcedo atthis), Its Conservation and Research, pp. 55–71. Proceedings of the 1st international seminar, 02/19 ZO ČSOP Alcedo, Vlašim, Czech Republic (in Czech with extended abstract in English).
- Čech, M. & Čech, P. 2011. Diet of the common kingfisher (Alcedo atthis) in relation to habitat type: a summary of results from the Czech Republic. Sylvia 47: 33–47 (in Czech with abstract and extended summary in English).
- Čech, M. & Čech, P. 2013. The role of floods in the lives of fish-eating birds: predator loss or benefit? Hydrobiologia 717: 203–211. doi: 10.1007/s10750-013-1625-3
- Čech, M. & Vejřík, L. 2011. Winter diet of great cormorant (Phalacrocorax carbo) on the River Vltava: estimate of size and species composition and potential for fish stock losses. Folia Zool. 60: 129–142.
- Čech, M., Čech, P., Kubečka, J., Prchalová, M. & Draštík, V. 2008. Size selectivity in summer and winter diets of great cormorant (Phalacrocorax carbo): does it reflect a season-dependent difference in foraging efficiency? Waterbirds 31: 438–447. doi: 10.1675/1524-4695-31.3.438
- Cramp, J.S. (ed). 1990. The Birds of the Western Palearctic. Vol. 4. Oxford University Press, Oxford.
- Emmrich, M. & Düttmann, H. 2011. Seasonal shifts in diet composition of great cormorants Phalacrocorax carbo sinensis foraging at a shallow eutrophic inland lake. Ardea 99: 207–216. doi: 10.5253/078.099.0211
- Gagliardi, A., Martinoli, A., Preatoni, D., Wauters, L.A. & Tosi, G. 2007. From mass of body elements to fish biomass: a direct method to quantify food intake of fish eating birds. Hydrobiologia 583: 213–222. doi: 10.1007/s10750-006-0528-y
- Gwiazda, R. & Amirowicz, A. 2010. Towards the optimal foraging strategy: is seasonal shift in the diet of cormorants Phalacrocorax carbo (L.) in a dam reservoir the effect of water temperature or size pattern in fish assemblages? Pol. J. Ecol. 58: 783–792.
- Hájková, P., Roche, K. & Kocian, Ľ. 2003. On the use of diagnostic bones of brown trout, Salmo trutta m. fario, grayling, Thymallus thymallus and Carpathian sculpin, Cottus poecilopus in Eurasian otter, Lutra lutra diet analysis. Folia Zool. 52: 389–398.
- Hall, C.S. & Kress, S. W. 2008. Diet of nestling black-crowned night-herons in a mixed species colony: implications for tern conservation. Wilson J. Ornithol. 120: 637–640. doi: 10.1676/07-035.1
- Hallet, C. 1977. Contribution a l'étude du régime alimentaire du Martin-Pêcheur (Alcedo atthis) dans la vallée de la Lesse. Aves 14: 128–144.
- Hallet, C. 1982. Etude du comportement de predation du Martin-Pêcheur Alcedo atthis (L.): taille preferentielle de capture du chabot Cottus gobio L. et de la truite Salmo trutta L. Revue d’ Ecologie (La Terre et la Vie) 36: 211–222.
- Hampl, R., Bureš, S., Baláž, P., Bobek, M. & Pojer, F. 2005. Food provisioning and nestling diet of the black stork in the Czech Republic. Waterbirds 28: 35–40. doi: 10.1675/1524-4695(2005)028[0035:FPANDO]2.0.CO;2
- Iribarren, I.B. & Nevado, L.D. 1982. Contribution a ľétude du régime alimentaire du Martin-Pêcheur (Alcedo atthis L. 1758). Alauda 50: 81–91.
- Jakubas, D. & Manikowska, B. 2011. The response of grey herons Ardea cinerea to changes in prey abundance. Bird Study 58: 487–494. doi: 10.1080/00063657.2011.608423
- Kazantzidis, S. & Goutner, V. 2005. The diet of nestlings of three Ardeidae species (Aves, Ciconiiformes) in the Axios Delta, Greece. Belg. J. Zool. 135: 165–170.
- Keller, T. 1998. The food of cormorants (Phalacrocorax carbo sinensis) in Bavaria. J. Ornithol. 139: 389–400 (in German with English summary). doi: 10.1007/BF01653465
- Lekuona, J.M. 2002. Food intake, feeding behaviour and stock losses of cormorants, Phalacrocorax carbo, and grey herons, Ardea cinerea, at a fish farm in Arcachon Bay (Southwest France) during breeding and non-breeding season. Folia Zool. 51: 23–34.
- Martinoli, A., Gagliardi, A., Preatoni, D.G., Di Martino, S., Wauters, L.A. & Tosi, G. 2003. The extent of great crested grebe predation on bleak in Lake Como, Italy. Waterbirds 26: 201–208. doi: 10.1675/1524-4695(2003)026[0201:TEOGCG]2.0.CO;2
- Montesinos, A., Santoul, F. & Green, A.J. 2008. The diet of the night heron and purple heron in the Guadalquivir Marshes. Ardeola 55: 161–167.
- Peris, S.J., Briz, F.J. & Campos, F. 1995. Shifts in the diet of the grey heron (Ardea cinerea) in the Duero basin, central-west Spain, following the introduction of exotic fish species. Folia Zool. 44: 97–102.
- Quiroga, M., Leon, E., Beltzer, A. & Olguin, P. 2013. Diet of black-crowned night-herons (Nycticorax nycticorax) in a wetland of the Parana River's alluvial valley. Ekoloji 22: 43–50. doi: 10.5053/ekoloji.2013.886
- Raven, P. 1986. The size of minnow prey in the diet of young kingfisher Alcedo atthis. Bird Study 33: 6–11. doi: 10.1080/00063658609476884
- Reynolds, S.J. & Hinge, M.D.C. 1996. Food brought to the nest by breeding kingfisher (Alcedo atthis) in the New Forest of southern England. Bird Study 43: 96–102. doi: 10.1080/00063659609460999
- Rodríguez, A., Rodríguez, B., Rumeu, B. & Nogales, M. 2007. Seasonal diet of the grey heron Ardea cinerea on an oceanic island (Tenerife, Canary Islands): indirect interaction with wild seed plants. Acta Ornithol. 42: 77–87. doi: 10.3161/068.042.0102
- Rozkošný, R. (ed.). 1980. The Key of the Water Insect Larvae. Academia, Praha.
- Suter, W. 1997. Roach rules: shoaling fish are a constant factor in the diet of cormorants Phalacrocorax carbo in Switzerland. Ardea 85: 9–27.
- Vacher, J.-P. & Rufray, X. 2005. Predation of Pelobates fuscus (Laurenti, 1768) by the kingfisher Alcedo atthis (Linnaeus, 1758). Herpetozoa 18: 189–190.
- Vilches, A., Miranda, R. & Arizaga, J. 2012. Fish prey selection of the common kingfisher Alcedo atthis in Northern Iberia. Acta Ornithol. 47: 169–177. doi: 10.3161/000164512X662278
- Vilches, A., Arizaga, J., Miranda, R. & Ibbotson, A. 2013. Impact of common kingfisher on a salmon population during the nestling period in southern England. Knowl. Manag. Aquat. Ecosyst. 410: 1–8.
- Vlček, V., Kestřánek, J., Kříž, H., Novotný, S., & Píše, J. 1984. Streams, Rivers and Reservoirs. Academia, Praha, Czech Republic.
- Woodall, P.F. 1991. Morphometry, diet and habitat in the kingfishers (Aves, Alcedinidae). J. Zool. 223: 79–90. doi: 10.1111/j.1469-7998.1991.tb04750.x