
Abstract
Capsule Ortolan Bunting occurrence is associated with bare ground, Lucerne, shrub cover and hedgerows/tree rows.
Aims To assess the habitat features selected by Ortolan Buntings at the territory level in semi-open landscapes, in the northern Apennines of Italy.
Methods We mapped territories in ten different plots and built a habitat selection model comparing 52 occupied cells with 52 unoccupied ones (cell size: 1 ha). We built multivariate adaptive regression splines models based on ground-measured variables.
Results The model revealed an association with intermediate Lucerne cover (50% of the cell), high shrub cover, bare ground (≥5%) and hedgerows/tree rows (≥25 m/ha). The most important driver of occurrence was bare ground (optimum at 5–20%).
Conclusion The maintenance of the mosaic and low-intensity farmed landscape, the promotion of Lucerne and the conservation/restoration of hedgerows/tree rows, may be promoted through the Rural Development Programme. The conservation of bare soil, grassland and shrubs at optimum amount at fine-scale could be the object of an agri-environment scheme targeted specifically at the Ortolan Bunting.
The Ortolan Bunting Emberiza hortulana is sharply declining. In the period 1980–2009, it underwent a dramatic decrease (–89% at the pan-European level), displaying the largest decline out of 38 widespread Afro-Palearctic migrant species (Vickery et al. Citation2014). This granivorous species is concentrated in Europe (which includes 50–74% of its range) and has an unfavourable conservation status in most European countries (BirdLife International Citation2004). It largely inhabits open or semi-open habitats, in rather warm and dry areas, and occupies a variety of breeding habitats, with apparently different preferences in different parts of its distribution (Cramp & Perrins Citation1994). In central and northern Europe, Ortolan Buntings mostly occur in heterogeneous and semi-open farmlands (Cramp & Perrins Citation1994, Dale & Olsen Citation2002, Goławski & Dombrowski Citation2002, Berg Citation2008); in southern Europe the species may occupy also (and often predominantly) open and semi-open shrubland or steppe-like habitats (Cramp & Perrins Citation1994, Guerrieri et al. Citation2006, Brotons et al. Citation2008, Menz et al. Citation2009), although farmed habitats may still be important in some temperate areas (Morelli Citation2012).
In general, Ortolan Buntings in the Mediterranean region are associated with areas of sparse vegetation and scattered trees (Cramp & Perrins Citation1994), whereas they are excluded by later stages of vegetation succession (Bogliani et al. Citation2003, Sirami et al. Citation2007). Both in southern and northern Europe, the species is often associated with burnt areas (Dale & Manceau Citation2003, Brotons et al. Citation2008, Menz et al. Citation2009). Therefore, the Ortolan Bunting may be considered a colonizer of the early vegetation stages (Menz et al. Citation2009). The species predominantly forages on sparsely vegetated or bare patches of ground. It nests mainly on the ground (Menz & Arlettaz Citation2012), but may rarely also nest in low bushes (Cramp & Perrins Citation1994). Trees (often in rows), rocks or bushes are required as song-posts (Cramp & Perrins Citation1994).
Several factors can affect the status of this declining species. Conditions experienced during migration and wintering periods may be important for this Afro-Palaearctic migrant, and in particular climate or anthropogenic changes in wintering grounds and illegal trapping during migration may exert notable effects (Menz & Arlettaz Citation2012 and references therein). However, the huge decline of the species had been largely ascribed to changes that have occurred in its breeding habitats. Detrimental changes include several types of modifications in agricultural practices and intensification (Goławski & Dombrowski Citation2002, Revaz et al. Citation2005, Vepsäläinen et al. Citation2005) including the switch from growing oats and rye to growing maize (Menz & Arlettaz Citation2012). The loss of patches of bare ground is particularly detrimental to the species, given its foraging ecology (Menz & Arlettaz Citation2012), and can be caused by vegetation closure through natural succession on abandoned areas (Sirami et al. Citation2007, Sondell et al. Citation2011), as well as by agricultural intensification and in particular by the heavy use of fertilizers (Menz & Arlettaz Citation2012). The role of changing breeding habitat in the decline of the Ortolan Bunting is highlighted by the fact that in areas where suitable breeding habitat is increasing (e.g. Spain, due to wildfires, Pons Citation2004, Brotons et al. Citation2008, Menz et al. Citation2009) or stable (e.g. central-eastern Italy, Morelli Citation2012), the species’ population trend is positive (Brotons et al. Citation2008, Menz & Arlettaz Citation2012, Morelli Citation2012). Therefore, a good understanding of the species’ ecological needs during the breeding period is of basic importance for its conservation, and given the different habitat associations reported from different parts of the species’ range, it is essential to develop region-specific approaches.
With this study, we aim to assess, at the territory level, which habitat features are selected by breeding Ortolan Buntings. On the basis of the common traits of habitat preferences and habitat use in Ortolan Buntings in different areas, we hypothesize that the species could be associated with bare ground, shrubs and selected crop types at the territory level (Golawski & Dombrowski Citation2002, Brotons et al. Citation2008, Menz et al. Citation2009, Morelli Citation2012).
METHODS
Study area
The study took place in Oltrepò pavese, southern Lombardy, province of Pavia, in northern Italy. The whole area extends over approximately 1100 km2 and is characterized, from north to south, by a gradient of increasing elevation, from 50 m above sea level (a.s.l.) at the Po River, to 1700 m a.s.l. for the highest mountain in the area. The lowland is dominated by cereal cultivation with small woodlots and other habitats, the foothills by vineyards, the low-elevation mountainsides by non-intensive cultivation and woodland, and the middle and upper elevations by woodland with scattered pastures, partly subjected to abandonment (Brambilla et al. Citation2012, Brambilla Citation2015). In Lombardy, as well as in Italy in general (Gustin et al. Citation2009), the Ortolan Bunting has an unfavourable status (‘bad’ conservation status; Brambilla et al. Citation2013).
Fieldwork
Territory mapping of Ortolan Buntings was carried out in April–June 2011, in ten plots (average size 116 ha ± 37 ha; see Supplemental data for further information), which were visited at least four times. Those plots were all dominated by open or semi-open landscapes, although the proportion of fields, grassland, vineyards and other habitats varied among plots. Territories were defined at the end of the fieldwork on the basis of simultaneous contacts and repeated observations as is usual for mapping studies (see Birrer et al. Citation2007, Brambilla et al. Citation2009 and references therein for similar methods).
Habitat selection model
Habitat variables were measured in the field at all territories, and at an equal number of unoccupied sites, to quantify the fine-scale land-use cover and habitat structure (). Variables were measured within a grid consisting of 100 m × 100 m square cells (1 ha), which was superimposed on the study area. The size of the grid cells was chosen to match the approximate size of territories at high density and the size of the ‘core area’ mostly used by individuals during the breeding season. Ortolan Bunting pairs may sometimes show a weak territorial behaviour (Cramp & Perrins Citation1994), and they can occur a few tens of meters apart (Cramp & Perrins Citation1994; pers. obs.); densities up to 0.8 pairs per ha and 1 male per ha have been reported from central and northern Italy (Gustin et al. Citation2009). When a single territory was spread across two neighbouring cells, its associated habitat variables were defined as the average features of the two cells. We found 52 territories and considered all them for analysis, together with an equal number of randomly chosen control cells. The only constraint was that the number of control and territory cells within each sub-area should be the same.
Table 1. Habitat variables used to model fine-scaled habitat selection in Ortolan Buntings in northern Italy.
The habitat selection model was built using multivariate adaptive regression splines (MARS), which related Ortolan Bunting occurrence to the habitat variables reported in . MARS is a flexible machine-learning technique (Friedman Citation1991, Hastie et al. Citation2009) often used in ecological studies (Leathwick et al. Citation2005, Elith & Leathwick Citation2007, MacNally et al. Citation2008, Heinanen & von Numers Citation2009). Its ability to cope with nonlinear effects makes it particularly suitable for investigating the habitat selection of a species like the Ortolan Bunting, which inhabits complex mosaic habitats and so may be associated with different variables according to different patterns, with specific thresholds or preferred intervals for each relevant habitat type. Prior to MARS analyses, variable correlations were checked; no pair of variables was highly intercorrelated (|r| < 0.7 for all pairs). The analysis was carried out using the earth package version 3.2–1 (http://cran.r-project.org/web/packages/earth/index.html; Milborrow Citation2011a) in R 3.0.1 (R Development Core Team Citation2013). The following settings were used for model selection: threshold = 0.001, penalty = 3, degree of interactions = 1 (no interaction allowed among variables). We used a penalty value of 3 instead of the commonly adopted 2 for models without interactions, because the results were identical for the two values (same variables and same species–habitat relationships), apart for a drop in the occurrence probability at intermediate-low level of shrub cover with penalty 2, which was biologically meaningless and likely due to overfitting. Therefore, we used a penalty equal to 3, which provided the same results without such a drop. The model was subjected to a five-fold cross-validation to estimate the model performance over different subsets of the original data.
Model discriminatory ability was evaluated by means of the area under the curve (AUC) calculated on the cross-validation, whereas variable importance was estimated by means of the evimp command (Milborrow Citation2011a). The latter is performed with the earth package and uses three different criteria to estimate variable importance in MARS models (see Milborrow Citation2011a and Jedlikowski et al. Citation2014 for details). The plotmo package version 1.3–1 (http://cran.r-project.org/web/packages/plotmo/index.html) was used to plot the fitted functions (Milborrow Citation2011b).
RESULTS
The MARS model for habitat selection identified the most important four habitat variables as: Lucerne Medicago sativa cover, shrub cover, cover of bare soil, total length of hedgerows and tree rows. Ortolan Buntings were associated with intermediate Lucerne cover (around 50% of the cell), high shrub cover, bare ground (at least 5% of the cell), length of hedgerows or tree rows (at least 25 m/ha) ( and ).
Figure 1. Graphical summary of the selected MARS model for habitat selection in Ortolan Bunting at the territory level. The species–habitat relationships represent the probability of species occurrence (on Y axis) in relation to habitat variables (on X axis; unit: percentage cover for the three cover variables, linear meters for length of hedgerows and tree rows) within the 1-ha cell.
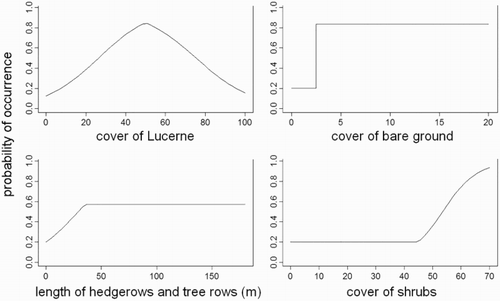
Table 2. Summary of the MARS model for fine-scaled habitat selection. For a correct interpretation of variable effect on the occurrence probability of Ortolan Buntings, refer to . RSS: decrease in the residual sum of squares; GCV: generalized cross-validation of the model.
The MARS model explained a fairly good portion of the initial deviance, with R2 = 0.38. The five-fold cross-validated model had R2 = 0.22 and an AUC = 0.80 ± 0.08 sd. The evimp function for the MARS model suggested the following ranking of variable importance (factors listed from the most to the less important): bare ground, Lucerne cover, shrub cover, tree rows and hedgerows.
DISCUSSION
The decline of the Ortolan Bunting has been linked to changes in its breeding habitat, so an understanding of the ecological requirements of the species in the breeding period is crucial for its conservation. Our results confirmed the primary importance of bare ground. Bare ground has been repeatedly reported as fundamental for Ortolan Buntings because it is a preferred foraging habitat (Menz & Arlettaz Citation2012 and references therein). At the small scale of this investigation, the model predicted high occurrence probabilities are associated with a 5–20% cover of bare ground. Moreover, the occurrence probability peaked with an intermediate cover of Lucerne, a 60–70% cover of shrubs and at least 25 m/ha of tree rows and hedgerows. Both Lucerne availability and hedgerow abundance are known to promote the occurrence of some species of conservation concern in the same area (Brambilla & Rubolini Citation2009, Brambilla et al. Citation2009). Lucerne is likely to be preferred by birds foraging in bare ground or sparse vegetation, such as Ortolan Buntings (Menz & Arlettaz Citation2012, Morelli Citation2012) or Woodlarks Lullula arborea (Brambilla & Rubolini Citation2009), because it is often seeded at a relatively low density, and usually Lucerne fields in these hilly areas offer easily accessible ground for several weeks during the breeding period (Brambilla & Rubolini Citation2009). Shrubland may offer plenty of song-posts, shelter and also alternative nesting sites (lower bushes; Cramp & Perrins Citation1994). Most of the shrubland included in territories was characterized by generally low growing species (e.g. Juniperus communis, Genista pilosa, Spartium junceum, Cytisus sessilifolius), with some scattered taller bushes (e.g. Prunus sp.), which were often used as song-posts. The association with shrub cover may be further promoted by the rather loose structure of shrublands in sloping areas, where soil erosion may increase the availability of very small patches of bare ground, sometimes not fully discernible at first sight but occurring among shrubs and potentially exploited by foraging Ortolan Buntings.
At the wider landscape level, the presence of grassland was well represented, so the lack of association between it and Ortolan Buntings was potentially due to the lack of variation in cover of grassland for cells with (mean 21%) and without (26%) buntings. No positive or negative associations were found with other types of cultivation. Although relatively common in the area, cereal fields, vineyards, and the less widespread mixed fodder and orchards, had no tangible effect on the occurrence probability of Ortolan Buntings.
Given the likely overwhelming importance of conserving suitable habitats in the breeding grounds of Ortolan Bunting, the conclusions of our study could be used to inform management recommendations for the species’ conservation, at least in this part of its range. The first implication is the maintenance of the typical low-intensity farming mosaic of the Apennines. The heterogeneous landscape of hilly and low-mountain sites in this area is characterized by a mix of relatively small fields separated by hedgerows, vineyards, grassland, shrublands, woodlots and calanques (sandy or rocky mountainsides strongly subjected to erosion). This mixture offers complementary resources such as song-posts, foraging and nesting habitats, to Ortolan Buntings (Menz & Arlettaz Citation2012) and other species of conservation concern (Bogliani et al. Citation2003, Brambilla et al. Citation2012 and references therein). The main threats to this mosaic landscape are represented by abandonment (Brambilla et al. Citation2010) and secondarily by agricultural intensification, especially with vineyard expansion at the expense of semi-natural grassland and shrubland (Bogliani et al. Citation2003), but also by interventions targeted at stabilizing slopes. In the past decades, the latter interventions have consisted of planting trees (mostly non-native species) over grassland with scattered shrubs to prevent soil erosion and calanque formation, which has the effect of reducing the fine-scaled habitat mosaic that is so suitable for the species.
The maintenance of the mosaic landscape associated with low-intensity farming (with particular emphasis on grassland, strongly declining in the area; Brambilla et al. Citation2010 and references therein), as well as the promotion of Lucerne and the conservation or restoration of hedgerows and tree rows, may be promoted by a correct definition of the measures included in the Rural Development Programme, which in Italy is defined at the regional level. The creation of patches of bare soil and the relative amount of this and other specific habitat features in compact habitat mosaics, including grassland, shrubs and bare ground, could be the object of a dedicated agri-environment scheme targeted at the Ortolan Bunting.
Supplemental data
Supplemental data for this article can be accessed at 10.1080/00063657.2015.1126219.
Supplementary material
Download PDF (1.6 MB)ACKNOWLEDGEMENTS
We are very grateful to F. Reginato for help with variable measurements and to E. Vigo, R. Falco, V. Bergero and two anonymous reviewers for suggestions and cooperation.
Additional information
Funding
References
- Berg, Å. 2008. Habitat selection and reproductive success of Ortolan Buntings Emberiza hortulana on farmland in central Sweden – the importance of habitat heterogeneity. Ibis 150: 565–573.
- BirdLife International. 2004. Birds in Europe: Population Estimates, Trends and Conservation Status. BirdLife International, Cambridge.
- Birrer, S., Spiess, M., Herzog, F., Jenny, M., Kohli, L. & Lugrin, B. 2007. The Swiss agrienvironment scheme promotes farmland birds: but only moderately. J. Ornithol. 148 (suppl. 2): 295–303.
- Bogliani, G., Cova, C. & Polani, F. 2003. La natura tra Nure e Scrivia. Il territorio del Giardino di Pietra Corva. Provincia di Pavia, Pavia.
- Brambilla, M. 2015. Landscape traits can contribute to range limit equilibrium: habitat constraints refine potential range of an edge population of Black-headed Bunting Emberiza melanocephala. Bird Study 62: 132–136.
- Brambilla, M. & Rubolini, D. 2009. Intra-seasonal changes in distribution and habitat associations of a multi-brooded bird species: implications for conservation planning. Anim. Conserv. 12: 71–77.
- Brambilla, M., Guidali, F. & Negri, I. 2009. Breeding-season habitat associations of the declining Corn Bunting Emberiza calandra – a potential indicator of the overall bunting richness. Ornis Fenn. 86: 41–50.
- Brambilla, M., Casale, F., Bergero, V., Bogliani, G., Crovetto, G.M., Falco, R., Roati, M. & Negri, I. 2010. Glorious past, uncertain present, bad future? Assessing effects of land-use changes on habitat suitability for a threatened farmland bird species. Biol. Conserv. 143: 2770–2778.
- Brambilla, M., Falco, R. & Negri, I. 2012. A spatially explicit assessment of within-season changes in environmental suitability for farmland birds along an altitudinal gradient. Anim. Conserv. 15: 638–647.
- Brambilla, M., Gustin, M. & Celada, C. 2013. Species appeal predicts conservation status. Biol. Conserv. 160: 209–213.
- Brotons, L., Herrando, S. & Pons, P. 2008. Wildfires and the expansion of threatened farmland birds: the Ortolan Bunting Emberiza hortulana in Mediterranean landscapes. J. Appl. Ecol. 45: 1059–1066.
- Cramp, S. & Perrins, C.M. (eds) 1994. The Birds of the Western Palearctic, Vol. 9. Oxford University Press, Oxford.
- Dale, S. & Manceau, N. 2003. Habitat selection of two locally sympatric species of Emberiza buntings (E. citrinella and E. hortulana). J. Ornithol. 144: 58–68.
- Dale, S. & Olsen, B.F.G. 2002. Use of farmland by Ortolan Buntings (Emberiza hortulana) nesting on a burned forest area. J. Ornithol. 143: 133–144.
- Elith, J. & Leathwick, J. 2007. Predicting species distributions from museum and herbarium records using multiresponse models fitted with multivariate adaptive regression splines. Divers. Distrib. 13: 265–275.
- Friedman, J.H. 1991. Multivariate adaptive regression splines. Ann. Stat. 19: 1–67.
- Goławski, A. & Dombrowski, A. 2002. Habitat use of yellowhammers Emberiza citrinella, Ortolan Buntings E. hortulana, and Corn Buntings Miliaria calandra in farmland of east-central Poland. Ornis Fenn. 79: 164–172.
- Guerrieri, G., Miglio, M. & Santucci, B. 2006. Habitat e riproduzione dell'Ortolano, Emberiza hortulana, in ambienti agricoli marginali dell'Italia centrale. Riv. Ital. Orn. 76: 47–68.
- Gustin, M., Brambilla, M. & Celada, C. 2009. Valutazione dello stato di conservazione dell'avifauna italiana. Ministero dell'Ambiente e della Tutela del Territorio e del Mare & LIPU/BirdLife Italia, Roma.
- Hastie, T., Tibshirani, R. & Friedman, J. 2009. The Elements of Statistical Learning: Data Mining, Inference, and Prediction. Springer-Verlag, New York.
- Heinanen, S. & von Numers, M. 2009. Modelling species distribution in complex environments: an evaluation of predictive ability and reliability in five shorebird species. Divers. Distrib. 15: 266–279.
- Jedlikowski, J., Brambilla, M. & Suska-Malawska, M. 2014. Fine-scale selection of nesting habitat in Little Crake Porzana parva and Water Rail Rallus aquaticus in small ponds. Bird Study 61: 171–181.
- Leathwick, J.R., Rowe, D., Richardson, J., Elith, J. & Hastie, T. 2005. Using multivariate adaptive regression splines to predict the distributions of New Zealand's freshwater diadromous fish. Freshw. Biol. 50: 2034–2052.
- Mac Nally, R., Fleishman, E., Thomson, J.R. & Dobkin, D.S. 2008. Use of guilds for modelling avian responses to vegetation in the Intermountain West (USA). Glob. Ecol. Biogeogr. 17: 758–769.
- Menz, M.H.M. & Arlettaz, R. 2012. The precipitous decline of the Ortolan Bunting Emberiza hortulana: time to build on scientific evidence to inform conservation management. Oryx 46: 122–129.
- Menz, M.H.M., Brotons, L. & Arlettaz, R. 2009. Habitat selection by Ortolan Buntings Emberiza hortulana in post-fire succession in Catalonia: implications for the conservation of farmland populations. Ibis 151: 752–761.
- Milborrow, S. 2011a. Package ‘earth’ 3.2–1. Multivariate adaptive regression spline models. Available from: http://cran.r-project.org/web/packages/earth.
- Milborrow, S. 2011b. Plotmo: plot a model's response while varying the values of the predictors. Available from: http://cran.r-project.org/web/packages/plotmo.
- Morelli, F. 2012. Correlations between landscape features and crop type and the occurrence of the Ortolan Bunting Emberiza hortulana in farmlands of Central Italy. Ornis Fenn. 89: 264–272.
- Pons, P. 2004. Hortolà Emberiza hortulana. In Estrada, J., Pedrocchi, V., Brotons, L. & Herrando, S. (eds) Atles dels ocells nidificants de Catalunya 1999–2002 [Catalan Breeding Bird Atlas 1999–2002]: 544–545. Institut Català d'Ornitologia and Lynx Edicions, Barcelona.
- R Development Core Team. 2013. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
- Revaz, E., Posse, B., Gerber, A., Sierro, A. & Arlettaz, R. 2005. Quel avenir pour le Bruant Ortolan Emberiza hortulana en Suisse? Nos Oiseaux 52: 67–82.
- Sirami, C., Brotons, L. & Martin, J.L. 2007. Vegetation and songbird response to land abandonment: from landscape to census plot. Divers. Distrib. 13: 42–52.
- Sondell, J., Brookes, C. & Persson, M. 2011. Ortolan Bunting Emberiza hortulana at Kvismaren, central Sweden – breeding studies and suggested management. Ornis Svecica. 21: 167–178.
- Vepsäläinen, V., Pakkala, T., Piha, M. & Tiainen, J. 2005. Population crash of the Ortolan Bunting Emberiza hortulana in agricultural landscapes of southern Finland. Annal. Zool. Fenn. 42: 91–107.
- Vickery, J.A., Ewing, S.R., Smith, K.W., Pain, D.J., Bairlein, F., Škorpilová, J. & Gregory, R.D. 2014. The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156: 1–22.