
ABSTRACT
Wintering Northern Wheatears Oenanthe oenanthe in the Sahel region of Northern Nigeria held small (approximately 70 m diameter) distinct territories during the study period, and territory size did not differ between adult and first winter birds. Evidence suggests that Wheatears may maintain small territories for a significant duration of the winter, similar to many other migrants.
The decisions migrant birds make during the non-breeding period can carry over to influence many aspects of their population dynamics (Baillie & Peach Citation1992, Sherry & Holmes Citation1996, Norris et al. Citation2004, Both et al. Citation2006, Newton Citation2006, Pulido Citation2007, Studds et al. Citation2008, Reudink et al. Citation2009, Newton Citation2010). Many migrant species are site faithful (territorial) during the non-breeding season (Mcneil Citation1982, Holmes & Sherry Citation1992, Warkentin & Hernandez Citation1996, Sauvage et al. Citation1998, Marra Citation2000, Salewski et al. Citation2000, King & Hutchinson Citation2001, Cresswell Citation2014, Blackburn & Cresswell Citation2016); a decision which can have substantial consequences for survival (Monroy-Ojeda et al. Citation2013), the degree of migratory connectivity (Cresswell Citation2014), how susceptible a species is to the loss or alteration of one or more wintering sites (Sutherland Citation1998, Newton Citation2010), gene flow, and the conservation of distinct populations (Iverson & Esler Citation2006). Knowing whether species are site faithful during winter thus has numerous consequences for the conservation of migrants, many of which are declining (Sherry & Holmes Citation1996, Warkentin & Hernandez Citation1996, Jones & Cresswell Citation2010, Vickery et al. Citation2014).
Here we establish the extent to which the Northern Wheatear Oenanthe oenanthe, a long-distance migrant, is site faithful on its wintering grounds in West Africa, and give estimates of winter territory size. Although winter territoriality has been documented for this and other Oenanthe species (Sharland Citation1967, Cornwallis Citation1975, Panov Citation1999, Khoury et al. Citation2012), the scale of this within-winter site fidelity has not been explored in detail. Here we use colour ringing to closely track individually identifiable birds. We also examine age-dependent effects because aspects of migrant wintering ecology may differ with age (Marra Citation2000). Although population trends vary throughout the Northern Wheatear’s range, the European breeding population (25–49% of the global range) has experienced a significant moderate decline since 1980 (Birdlife International Citation2004).
The study took place in the winter of 2011 in Katsina state in the Sahel region of Northern Nigeria, bordering Niger (13°00′56.9″N, 7°07′22.6″E). The study site was a dry and rocky open area with patches of scrub, Acacia trees and bushes, and boulders, intersected with a dry riverbed (). We captured and colour-ringed 41 Northern Wheatears over an area of approximately 1.3 km2, between 15 November and 8 December, with the exception of two individuals that were captured earlier in mid-October during a preliminary visit to the site. Birds were captured using baited spring traps with conspecific playback and each uniquely colour-ringed, sexed, aged as adult or first winter (Jenni & Winkler Citation2004), and biometrics recorded (maximum wing chord, tarsus, mass, fat, and pectoral muscle scores). We resighted colour-ringed birds on 17 visits to the site between 3 and 14 December of the same winter. We resighted between dawn (approximately 06:00 hours) and 11:00 hours, and/or 16:00 hours until dusk (approximately 18:30 hours) using binoculars to locate birds (Swarovski SV 8 × 32) and a spotting scope to read combinations (Zeiss Diascope 65 mm with 25× eyepiece). Approximately 55 hours in total were dedicated to resighting ringed birds. The average number of resighting visits made to the site after a bird had been ringed was 15.7 visits (±0.3, range = 11–17; although note that not every territory was necessarily visited during a resighting visit). We attempted to locate all birds before they were disturbed and scored each sighting regarding whether the bird had been disturbed before being sighted. In the majority of cases, only one resighting location was recorded for each bird per site visit and only if at least 3 hours after a previous resighting. Site visits were separated by at least 5 hours, and Wheatears could cross their territories in seconds; therefore we are confident that resighting locations are reasonably independent. Locations of ringed individuals were recorded with a GPS device (Garmin GPSMAP 64). Coordinates were converted to UTM units (Zone 32N) for analyses. Resightings were viewed and edited where necessary in ArcMAP 10.1 (Ersi Citation2012) and Garmin BaseCamp software (version 4.2.4; Garmin International, Inc., Olathe, KS, USA).
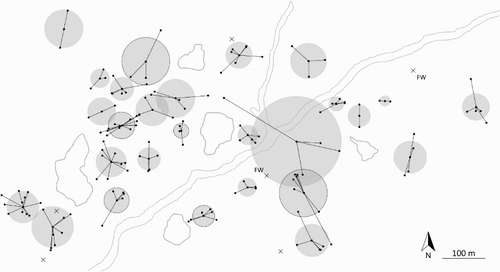
Figure 1. Individual territories of 33 Northern Wheatears wintering in northern Nigeria. Each point represents an undisturbed resighting, connected to the mean location of all resightings for that individual. The grey shaded circles illustrate the territory size for each individual resighted at least twice (n = 27 birds). Black crosses show the location of birds resighting only once following capture (n = 6 birds). Territories outlined in black and crosses with ‘FW’ show the territories of first winter birds. Shapes with grey outlines show the location of rocky outcrops and a dry riverbed. This figure shows that Wheatears held small, distinct territories throughout the duration of the resighting period.
We used an index to illustrate an estimate of territory size instead of the conventional method of minimum convex polygons, which requires many resightings for accurate descriptions of space use and is subject to several biases (Hansteen et al. Citation1997, Börger et al. Citation2006). For each individual, we averaged the location of all resightings scored as undisturbed to find the centre of the bird’s territory. We then calculated the distance of each resighting to the territory centre, and averaged this distance to give the mean distance of each resighting from the territory centre. This value was then doubled this to give a measure of territory size. Full methods are described in Blackburn & Cresswell (Citation2015). Because territory size estimates calculated from very few resightings were likely to be less accurate, we then repeated the above for birds only with at least five resightings in an attempt to reduce the influence of increasing resighting sample size on estimates of territory size (Börger et al. Citation2006), whilst still maintaining adequate sample sizes. Fifteen birds had at least five independent resightings. Although coordinates recorded with GPS devices have an error of approximately ±5 m (our device gave an accuracy of approximately ±2–3 m in Nigeria), we assume here that this error was similar across all recorded points and did not adjust for GPS inaccuracy when calculating our index of territory size. We used analysis of variance (anova) to explore the influence of age (adult versus first winter) on territory size, a general linear model (GLM) to test whether the estimate of territory size was influenced by the number of resightings for birds resighted at least five times, and logistic regression (GLM) to test whether the probability of resighting a bird was related to the number of resightings visits made after it was ringed. Analyses were carried out in R version 3.0.1 (R Development Core Team, Citation2013). Mean values are presented as means ± one standard error in all cases. A statistical significance level of P < 0.05 was chosen to reject null hypotheses.
Of the 41 Northern Wheatears ringed, 33 were resighted at least once after capture (mean number of resightings per individual = 4.6 ± 0.5, range = 1–13). The number of resighting visits made after ringing a bird was the same for birds that we never resighted compared to those that were resighted (GLM of resighted (y/n) ∼ number of resighting visits after ringing: β = 0.10 ± 0.18, z = 0.5, P = 0.60, n = 41); therefore the eight birds that were not resighted after capture may have been transients. Resighted birds were last seen between 4 and 53 days after capture (mean = 14.0 ± 2.2 days). All resighted individuals were observed to hold small, distinct territories during the entirety of the study (). The two individuals ringed in late October during the first visit were resighted 50 and 53 days later at their exact capture locations. Mean territory diameter was 73.5 m across all birds resighted at least twice (the minimum number of locations to calculate a territory size; ±7.2 m, range 25.2–220.0, n = 27; ). For birds resighted at least five times, mean territory size was 69.4 m (± 5.4 m, range 35.0–114.1, n = 15). The number of resightings had no influence on our estimates of territory size for birds resighted at least five times (lm of territory size∼number of resightings: P = 0.93, F1,13 = 0.25, n = 15). There was no difference in territory size between adult and first winter birds (lm of territory size∼age: P = 0.97, F2,23 = 0.03, n = 25, of which 20 = adult, 5 = first winter).
By following individually identifiable birds during part of the wintering period, we show that the Northern Wheatear, a long-distance migrant, is territorial for at least part of the wintering period. Our study period was relatively short, yet both Wheatears ringed during an earlier visit were resident in the same territories at least until the end of the study period (50 and 53 days). Furthermore, it was evident that territories were already established on our arrival in early- to mid-winter. This suggests that Wheatears are territorial for a large part, if not all, of the wintering period. Individuals responded strongly to playback, and territorial behaviour (singing, chasing neighbouring birds) was observed throughout the study. Although eight birds were not resighted after ringing and so may have been transient birds that did not overwinter in the study area, resightings were not extensive in effort or time, and we cannot be certain that this was the case. Our findings therefore agree with other studies documenting strong winter territoriality and site fidelity in several Oenanthe species, including Northern Wheatears (Cornwallis Citation1975, Panov Citation1999, Khoury et al. Citation2012, but see Smith Citation1971); for example, for Northern Wheatears wintering in Egypt and at stopovers during migration (Arizaga et al. Citation2011), and for Black-eared Wheatears Oenanthe hispanica wintering in Nigeria (Sharland Citation1967), but it should be noted than many of these accounts are anecdotal and did not closely follow individually identifiable birds.
High site fidelity within winters appears to be common across migrants both in the Palearctic and Neotropics (Moreau Citation1969, Kelsey Citation1989, Bates Citation1992, Brown et al. Citation2000, Salewski et al. Citation2000, Wunderle & Latta Citation2000, King & Hutchinson Citation2001, Latta & Faaborg Citation2001, Rappole et al. Citation2003, Barg et al. Citation2006, Monroy-Ojeda et al. Citation2013, Blackburn & Cresswell Citation2016). Likely, winter territoriality increases survival for these species. Moving during winter or not holding a territory is often associated with reduced survival (Rappole et al. Citation1989, Winker Citation1998), whereas being territorial can increase survival probability through familiarity and predictability of food resources and surroundings (Brown & Long Citation2007, Förschler et al. Citation2010).
Measures of territory size are scarce for Wheatears, but our findings are similar to winter territory sizes documented for Finsch’s Wheatears Oenanthe finschii (Khoury et al. Citation2012). Territory size did not differ between first winter and adult birds and the location of first winter territories was not distinct from that of adults (), and there appears to be no spatial segregation according to age, unlike some Neotropical examples (Stutchbury Citation1994, Marra & Holmes Citation2001). However, the number of first winter birds in our sample is small, our measure of territory size is relatively crude and based on small sample sizes for each individual and we do not measure territory quality. Nonetheless, territory size, location, and quality does not differ according to age and sex for Whinchats Saxicola rubetra, a similar Palearctic migrant (Blackburn & Cresswell Citation2015, Citation2016). This suggests that – like Whinchats – Northern Wheatears probably do not exhibit strong dominance hierarchies according to age on the wintering grounds, in contrast to several Neotropical species which do (Stutchbury Citation1994, Marra Citation2000). Further work is required to confirm whether the absence of age or dominance effects on territoriality is the general case for Palearctic wintering passerines.
In summary, we accurately document high within-winter site fidelity in a long-distance Palearctic migrant and suggest that Northern Wheatears maintain territories throughout the wintering period. More detailed knowledge of winter residency and whether individuals are capable of relocating under changing conditions is essential for the effective conservation of migrants given the increasing anthropogenic-driven decline across wintering habitats (Vickery et al. Citation2014).
Acknowledgements
We thank Arin Izang, Sulaiman Inuwa Muhammad, Chris Odey, Azi Isha Bazane, and staff of the A.P. Leventis Ornithological Research Institute for their support and assistance with fieldwork. This is paper number (107) from the AP Leventis Ornithological Research Institute. This study complies with the current ethic regulations of Nigeria and the A.P. Leventis Ornithological Research Institute.
Additional information
Funding
References
- Arizaga, J., Schmaljohann, H. & Bairlein, F. 2011. Stopover behaviour and dominance: a case study of the Northern Wheatear Oenanthe oenanthe. Ardea 99: 157–165. doi: 10.5253/078.099.0205
- Baillie, S.R. & Peach, W.J. 1992. Population limitation in Palearctic-African migrant passerines. Ibis 134: 120–132. doi: 10.1111/j.1474-919X.1992.tb04742.x
- Barg, J.J., Aiama, D.M., Jones, J., Robertson, R.J. & Yasukawa, K. 2006. Within-territory habitat use and microhabitat selection by male Cerulean Warblers (Dendroica cerulea). Auk 123: 795–806. doi: 10.1642/0004-8038(2006)123[795:WHUAMS]2.0.CO;2
- Bates, J.M. 1992. Winter territorial behavior of Gray Vireos. Wilson Bull. 104: 425–433.
- Birdlife International. 2004. Birds in Europe: population estimates, trends and conservation status. BirdLife Conservation Series No. 12.
- Blackburn, E. & Cresswell, W. 2015. Fine-scale habitat use during the non-breeding season suggests that winter habitat does not limit breeding populations of a declining long-distance Palearctic migrant. J. Avian. Biol. 46: 622–633.
- Blackburn, E. & Cresswell, W. 2016. High winter site fidelity in a long-distance migrant: implications for wintering ecology and survival estimates. J. Ornithol. 157: 93–108. doi: 10.1007/s10336-015-1252-z
- Börger, L., Franconi, N., De Michele, G., Gantz, A., Meschi, F., Manica, A., Lovari, S. & Coulson, T. 2006. Effects of sampling regime on the mean and variance of home range size estimates. J. Anim. Ecol. 75: 1393–1405. doi: 10.1111/j.1365-2656.2006.01164.x
- Both, C., Sanz, J.J., Artemyev, A.V., Blaauw, B., Cowie, R.J., Dekhuizen, A.J., Enemar, A., Javinen, A., Nyholm, N.E.I., Potti, J., Ravussin, P.A., Silverin, B., Slater, F.M., Sokolov, L.V., Visser, M.E., Winkel, W., Wright, J. & Zang, H. 2006. Pied Flycatchers Ficedula hypoleuca travelling from Africa to breed in Europe: differential effects of winter and migration conditions on breeding date. Ardea 94: 511–525.
- Brown, D.R. & Long, J.A. 2007. What is a winter floater? Causes, consequences, and implications for habitat selection. Condor 109: 548–565. doi: 10.1650/8351.1
- Brown, D.R., Stouffer, P.C. & Strong, C.M. 2000. Movement and territoriality of wintering hermit thrushes in Southeastern Louisiana. Wilson Bull. 112: 347–353. doi: 10.1676/0043-5643(2000)112[0347:MATOWH]2.0.CO;2
- Cornwallis, L. 1975. The comparative ecology of eleven species of wheatears (genus Oenanthe) in SW Iran. PhD Thesis, Oxford University.
- Cresswell, W. 2014. Migratory connectivity of Palaearctic-African migratory birds and their responses to environmental change: the serial residency hypothesis. Ibis 156: 493–510. doi: 10.1111/ibi.12168
- ERSI. 2012. ArcGIS Desktop: Release 10. Environmental Systems Research Institute, Redlands, California.
- Förschler, M.I., Del Val, E. & Bairlein, F. 2010. Extraordinary high natal philopatry in a migratory passerine. J. Ornithol. 151: 745–748. doi: 10.1007/s10336-010-0495-y
- Hansteen, T.L., Andreassen, H.P. & Ims, R.A. 1997. Effects of spatiotemporal scale on autocorrelation and home range estimators. J. Wildlife Manage. 61: 280–290. doi: 10.2307/3802583
- Holmes, R.T. & Sherry, T.W. 1992. Site fidelity of migratory warblers in temperate breeding and Neotropical wintering areas: implications for population dynamics, habitat selection, and conservation. In Hagan III, J.M. & Johnston, D.W. (eds) Ecology and Conservation of Neotropical Migrant Landbirds. Smithsonian Institution Press, Washington, DC, 563–575.
- Iverson, S.A. & Esler, D. 2006. Site fidelity and the demographic implications of winter movements by a migratory bird, the harlequin duck Histrionicus histrionicus. J. Avian Biol. 37: 219–228. doi: 10.1111/j.2006.0908-8857.03616.x
- Jenni, L. & Winkler, R. 2004. Moult and Ageing of European Passerines. Academic Press, London.
- Jones, T. & Cresswell, W. 2010. The phenology mismatch hypothesis: are declines of migrant birds linked to uneven global climate change? J. Anim. Ecol. 79: 98–108. doi: 10.1111/j.1365-2656.2009.01610.x
- Kelsey, M.G. 1989. A comparison of the song and territorial behaviour of a long-distance migrant, the Marsh Warbler Acrocephalus palustris, in summer and winter. Ibis 131: 403–414. doi: 10.1111/j.1474-919X.1989.tb02788.x
- Khoury, F., Boulad, N. & Janaydeh, M. 2012. Territory size variations in wintering Finsch’s Wheatears, Oenanthe finschii: (Aves: Passeriformes). Zool. Middle East 57: 35–43. doi: 10.1080/09397140.2012.10648961
- King, J.M.B. & Hutchinson, J.M.C. 2001. Site fidelity and recurrence of some migrant bird species in The Gambia. Ringing Migr. 20: 292–302. doi: 10.1080/03078698.2001.9674255
- Latta, S.C. & Faaborg, J. 2001. Winter site fidelity of Prairie Warblers in the Dominican Republic. Condor 103: 455–468. doi: 10.1650/0010-5422(2001)103[0455:WSFOPW]2.0.CO;2
- Marra, P.P. 2000. The role of behavioral dominance in structuring patterns of habitat occupancy in a migrant bird during the nonbreeding season. Behav. Ecol. 11: 299–308. doi: 10.1093/beheco/11.3.299
- Marra, P.P. & Holmes, R.T. 2001. Consequences of dominance-mediated habitat segregation in American Redstarts during the nonbreeding season. Auk 118: 92–104. doi: 10.1642/0004-8038(2001)118[0092:CODMHS]2.0.CO;2
- McNeil, R. 1982. Winter resident repeats and returns of austral and boreal migrant birds banded in Venezuela. J. Field Ornithol. 53: 125–32.
- Monroy-Ojeda, A., Grosselet, M., Ruiz, G. & Valle, E.D. 2013. Winter site fidelity and winter residency of six migratory neotropical species in Mexico. Wilson J. Ornithol. 125: 192–196. doi: 10.1676/12-072.1
- Moreau, R.E. 1969. The recurrence in winter quarters (Ortstreue) of trans-Saharan migrants. Bird Study 16: 108–110. doi: 10.1080/00063656909476227
- Newton, I. 2006. Can conditions experienced during migration limit the population levels of birds? J. Ornithol. 147: 146–166. doi: 10.1007/s10336-006-0058-4
- Newton, I. 2010. The Migration Ecology of Birds. Academic Press, London.
- Norris, D.R., Marra, P.P., Kyser, T.K., Sherry, T.W. & Ratcliffe, L.M. 2004. Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. P R. Soc. Lond., Ser. B: Biol. Sci. 271: 59–64. doi: 10.1098/rspb.2003.2569
- Panov, E. 1999. Wheatears of Palearctic: Ecology, Behaviour and Evolution. KMK Scientific Press, Moscow.
- Pulido, F. 2007. Phenotypic changes in spring arrival: evolution, phenotypic plasticity, effects of weather and condition. Clim. Res. 35: 5–23. doi: 10.3354/cr00711
- Rappole, J.H., King, D.I. & Diez, J. 2003. Winter vs. breeding-habitat limitation for an endangered avian migrant. Ecol. Appl. 13: 735–742. doi: 10.1890/1051-0761(2003)013[0735:WVBLFA]2.0.CO;2
- Rappole, J.H., Ramos, M.A. & Winker, K. 1989. Wintering wood thrush movements and mortality in southern Veracruz. Auk 106: 402–410.
- R Development Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/.
- Reudink, M.W., Marra, P.P., Kyser, T.K., Boag, P.T., Langin, K.M. & Ratcliffe, L.M. 2009. Non-breeding season events influence sexual selection in a long-distance migratory bird. P. R. Soc. Lond., Ser. B: Biol. Sci. 276: 1619–1626. doi: 10.1098/rspb.2008.1452
- Salewski, V., Bairlein, F. & Leisler, B. 2000. Recurrence of some Palaearctic migrant passerine species in West Africa. Ringing Migr. 20: 29–30. doi: 10.1080/03078698.2000.9674224
- Sauvage, A., Rumsey, S. & Rodwell, S. 1998. Recurrence of Palaearctic birds in the lower Senegal river valley. Malimbus 20: 33–53.
- Sharland, R.E. 1967. Wheatears in winter quarters. Niger. Om. Soc. Bull. 4: 12–16.
- Sherry, T.W. & Holmes, R.T. 1996. Winter habitat quality, population limitation, and conservation of Neotropical-Nearctic migrant birds. Ecology 77: 36–48. doi: 10.2307/2265652
- Smith, K.D. 1971. Notes on Oenanthe species in winter in Africa. Bird Study 18: 71–80. doi: 10.1080/00063657109476298
- Studds, C.E., Kyser, T.K. & Marra, P.P. 2008. Natal dispersal driven by environmental conditions interacting across the annual cycle of a migratory songbird. P. Natl. Acad. Sci., USA 105: 2929–2933. doi: 10.1073/pnas.0710732105
- Stutchbury, B.J. 1994. Competition for winter territories in a Neotropical migrant: the role of age, sex and color. Auk 111: 63–69. doi: 10.2307/4088505
- Sutherland, W.J. 1998. The effect of local change in habitat quality on populations of migratory species. J. Appl. Ecol. 35: 418–421. doi: 10.1046/j.1365-2664.1998.00320.x
- Vickery, J.A., Ewing, S.R., Smith, K.W., Pain, D.J., Bairlein, F., Škorpilová, J. & Gregory, R.D. 2014. The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156: 1–22. doi: 10.1111/ibi.12118
- Warkentin, I.G. & Hernandez, D. 1996. The conservation implications of site fidelity: a case study involving Nearctic-Neotropical migrant songbirds wintering in a Costa Rican mangrove. Biol. Conserv. 77: 143–150. doi: 10.1016/0006-3207(95)00146-8
- Winker, K. 1998. The concept of floater. Ornitol. Neotrop. 9: 111–119.
- Wunderle Jr, J.M. & Latta, S.C. 2000. Winter site fidelity of Nearctic migrants in shade coffee plantations of different sizes in the Dominican Republic. Auk 117: 596–614. doi: 10.1642/0004-8038(2000)117[0596:WSFONM]2.0.CO;2