
ABSTRACT
Capsule: Restoration of grasslands on ski-pistes caused a recovery in the bird community, but not to the extent that it was equivalent to a natural Alpine grassland community.
Aim: To test whether revegetation of ski-pistes in open habitat areas results in bird community recovery.
Methods: The bird communities in two ski resorts in the Italian Maritime Alps were surveyed using a standardized area count method in three different plot types: non-restored ski-pistes (newly constructed), restored ski-pistes and control plots in grassland far from ski-pistes.
Results: In 49 independent plots, 32 species were recorded. Species richness and abundance of birds were significantly higher on restored than on non-restored ski-pistes, independently of the species group considered and the analyses carried out. Bird community parameters of restored ski-pistes were still lower than those of natural grassland, as shown by results of typical grassland species.
Conclusion: Our results suggest that an apparently successful restoration of ski-pistes may be not enough to promote a complete recovery of bird communities. The complete recovery of local bird communities may be promoted only if an integral recovery of the original vegetal communities is achieved. We suggest the best conservation option is to adopt techniques to maintain as far as possible original grassland if construction of new ski-pistes is unavoidable.
Over the last century, the European Alps have been subject to an increasing anthropogenic impact due to the building of ski-pistes and the development of winter tourism facilities (Simons Citation1988, Mackenzie Citation1989, Pechlaner & Tschurtschenthaler Citation2003). The skiing industry is of major economic importance in the alpine region, and it has recently experienced a period of great expansion (Koenig & Abegg Citation1997, Elsasser & Messerli Citation2001, Wipf et al. Citation2005). Several thousands of kilometres of ski-pistes are used for downhill skiing (Rolando et al. Citation2007), and in the Swiss Alps alone about 220 km2 are directly affected by ski-pistes (Amacher-Hoppler & Schoch Citation2008).
The main impact of ski-pistes is on vegetation and soils, since the natural vegetation and most of the upper soil horizons are removed during the construction process (machine-grading, used to smooth out underlying rock and soil), to provide suitable slopes for skiers and to enhance the use of artificial snow (Mosimann Citation1985, Wipf et al. Citation2005, Isselin-Nondedeu & Bédécarrats Citation2007, Delarze et al. Citation1998). Moreover, the use of snow-grooming vehicles in the preparation of pistes causes changes in the underlying soil structure and vegetation (Cernusca et al. Citation1990, Rixen et al. Citation2004). Finally, summer management at regular intervals, involving cutting of shrubs and machine-grading, produces further damage to vegetation (Bayfield Citation1996, Titus & Tsuyuzaki Citation1999). The recovery of vegetation may be successful below the treeline, whereas it is extremely difficult above the treeline, because of the scarcity of soil and the peculiar traits of high altitude plant species (very low growth rates, low seed production and insufficient agents of seed dispersal; Urbanska & Fattorini Citation2000).
In addition to the apparent negative effects on soils and vegetation, the impacts of winter recreation are most often negative for fauna. Results from meta-analyses have shown that richness, abundance and diversity of fauna were lower in areas affected by winter recreation when compared to undisturbed areas (Sato et al. Citation2013). Several studies have shown negative effects of ski-pistes on animals, such as birds (Laiolo & Rolando Citation2005, Rolando et al. Citation2007, Patthey et al. Citation2008, Caprio et al. Citation2011), small mammals (Hadley & Wilson Citation2004, Sanecki et al. Citation2006, Negro et al. Citation2009, Rolando et al. Citation2013), reptiles (Sato et al. Citation2014a, Citation2014b) and invertebrates (Negro et al. Citation2009, Citation2010, Citation2013, Rolando et al. Citation2013, Keßler et al. Citation2012, Kašák et al. Citation2013).
Below the treeline, ski-pistes through forest may induce habitat fragmentation that limits small mammal movements (Negro et al. Citation2012), whereas above the treeline they likely do not significantly cause habitat fragmentation, but landscape changes may nevertheless affect bird species richness and distribution (Caprio et al. Citation2011). The area above the treeline is of particular concern, because climate changes will probably induce operators and stakeholders to shift skiing activities and ski-pistes to higher altitudes (Elsasser & Messerli Citation2001, Fukushima et al. Citation2002, Bicknell & McManus Citation2006, Scott et al. Citation2007) where ecosystems are particularly sensitive (Körner Citation2003).
The scarcity of vegetation on ski-pistes of high altitude grasslands is likely the most relevant determinant of bird diversity. Sparsely grass-covered ski-pistes are landscape features which lower grassland species richness and probability of occurrence of certain passerine species (Caprio et al. Citation2011). Moreover, the amount of grass vegetation controls abundance and diversity of ground-dwelling arthropods (Negro et al. Citation2010) that, in turn, may directly affect birds feeding on invertebrates (Rolando et al. Citation2007). In general, a further problem regards detecting habitat effects at community levels. A completely holistic approach may be unsatisfactory because different species often react differently to the same environmental factors, therefore communities are often split into guilds, which group animals according to their eco-ethological characteristics (Verner Citation1984). Therefore, the response of specialist species (i.e. grassland species) may be different to those species that are more associated with shrubs and forests (Laiolo & Rolando Citation2005, Rolando et al. Citation2007).
In this paper, we tested the hypothesis that revegetation of grassland ski-pistes promotes bird community recovery. We considered ski-pistes located in pastures ranging from about 1500 to 2200 m above sea level (a.s.l.) at two ski resorts in the Maritime Alps (the southernmost part of the Alps). Here, due to the low altitude of several ski-pistes and the influence of the maritime climate, old ski-pistes have been recolonized by vegetation and are nowadays grass-covered. Other, more recent, ski-pistes have not recovered and appear as strips of bare ground with scarce vegetation. We therefore compared whole bird community and guild diversity in plots located in (i) ski-pistes of recent construction with depleted or no vegetation (hereafter non-restored ski-pistes), (ii) old ski-pistes that were entirely grass-covered, showing a rather natural appearance (hereafter restored ski-pistes) and (iii) natural grasslands. We also investigated bird–habitat relationships to identify structural components of the habitat (e.g. grass cover, shrub cover, height of vegetation etc.) that control composition and abundance of grassland bird communities.
Methods
Study area
The study was carried out within the Limone and Limonetto skiing districts in the Vermenagna Valley (southwestern Italian Alps; 44°11′N 7°33′E), partly encompassed within a protected area (Site of Community Interest IT1160056 ‘Alpi Marittime’). Beech Fagus sylvatica forests extend all over the area, but large pastures also occur at the same altitude as forests, from 1500 m a.s.l. upwards. Pastures are characterized by Nardus stricta prairies interspersed with shrub patches represented by Common Juniper Juniperus communis, Alpenrose Rhododendron ferrugineum and, to a lesser extent, Bilberry Vaccinium myrtillus. They are the outcome of intensive historical pastoral activities that at first removed Beech forests and then maintained open habitats. Cattle and sheep, even though pastoralism is declining, are still present in the area.
We considered all the ski-pistes of the study area (a total of 53 km), but sampling was carried out in open habitat only, that is where tracks crossed open pastures, from about 1500 to 2200 m a.s.l. Restored ski-pistes are 3–10 years old and are still in use but show in most cases a natural appearance, with a rather high grass cover. This depends on land use and climatic peculiarities of the study area. Because of the long history of pastoralism, open habitats also occur below the tree line that, due to the influence of the sea (the distance between Limone Piemonte and the Ligurian sea is less than 50 km), is set at a rather high altitude (about 1800 m for the Beech and 2500 m for the European Larch Larix decidua). Most of the open habitat ski-pistes are therefore below the natural tree line and because of this, their restoration (partly due to natural dynamics and partly due to artificial seeding) is quicker and more successful than that of other alpine localities where open habitat ski-pistes are located above the tree line. Moreover, during construction, original grass and topsoil were not removed everywhere; smooth pastures, in particular, were left untouched. Restored ski-pistes were not machine-graded during summer. Conversely, non-restored ski-pistes are of recent construction (1–2 years) or have experienced recent summer machine-grading and appear as strips with bare ground either without grass, or with a very few dispersed grass tufts. In most of the ski-pistes, snow-making is used to supplement natural snow during the winter season.
Bird and habitat surveys
Field work was carried out in the morning, during the first 4 hours after sunrise, in June and July 2011 by Enrico Caprio (EC), who sampled birds using a standardized area count method, surveying birds in circular plots of 50 m radius. Counts lasted 15 min, during the first 10 minutes of which the observer stood still and quiet at the centre of the plot, as in standard point counts. In the last 5 minutes of the count, the observer moved around, to flush secretive and non-singing individuals, and stopped at suitable vantage points to look and listen, recording all birds seen or heard within the plot (Rolando et al. Citation2007). Each census plot was visited twice (in June and July); the total number of species from the two censuses was used as a measure of species richness, and the higher number of individuals over the two visits was used as a measure of bird species abundance per plot. Three types of plots were defined: plots centred on non-restored ski-pistes (n = 14), on restored ski-pistes (n = 17) and in natural grassland habitats (n = 18). Points were selected on the basis of accessibility, also avoiding sources of possible disturbance (i.e. close to roads or livestock), or locations where detectability may have been reduced. At the same time, we kept other landscape features as constant as possible (i.e. distance to forest, slope, exposition and altitude).
When possible, sampling was organized in sets of three, the plots of the three types being located in the same skiing district with the same landscape and topography (see above). Ski-piste strips were 65–185 m wide (mean 114.2 ± 38.5 sd); when the strip was narrower than 100 m (restored ski-pistes n = 2, non-restored ski-pistes n = 3), a variable portion of open habitat at the side of the ski-piste was included in the plot. Plots were set at a minimum distance of 300 m. Avian communities were described in terms of two diversity parameters, species richness (S) and total abundance. Following Laiolo & Rolando (Citation2005), species were classified as ecotone/grassland or shrub/woodland according to their ecological preferences.
Altitude and seven habitat cover and structure variables were collected for each plot: percentage of shrub; stone-rock; soil-rubble (i.e. open-ground) and grass (all coverage estimated by eye); Shannon diversity of the vegetation (Shannon Index, ), where pi is the relative frequency of species i calculated according the relative frequency of vegetation types (area of grasses, Juniper, Alpenrose and other bushes estimated by eye); mean height of the vertical component of the habitat (mean of 20 measurements of shrub, stone-rock, soil-rubble and grass per plot, shared according to the relative cover percentages and recorded with a wooden dowel subdivided into 1-cm units) and heterogeneity of the vertical component (coefficient of variation (CV) = sd/mean × 100).
Data analysis
Differences between plot types
Differences in habitat cover (i.e. shrub, stone-rock, soil-rubble and grass cover) and habitat structure (i.e. H′ of vegetation cover, mean height and CV of vertical structure and heterogeneity) between the three plot types (i.e. non-restored ski-pistes, restored ski-pistes and natural grasslands) were tested by means of one-way anova. To attain the normal distribution, the values of Shannon index were log transformed [y = log(x + 1)]. Factor analysis (FA; Gaunch Citation1984) was chosen to reveal patterns in the data for habitat cover and structure (i.e. the seven variables listed above).
Differences in mean bird species richness and total abundance were tested by means of one-way anova. To attain the normal distribution, the values of species richness and total abundance were transformed to square roots [y = √(x + 0.5)].
Relationships between bird community and habitat
We tested for the effects of habitat cover (percentage of shrub, stone-rock, soil-rubble and grass cover), habitat structure (H′ of vegetation cover, mean height and CV of the vertical component of the habitat), plot type (categorical variable defining plots in natural grasslands, in non-restored and restored ski-pistes) and altitude on bird species richness and abundance of individuals using Generalized Linear Models (GLMs). To reduce correlation amongst variables, we first examined all pairwise correlations to identify strongly correlated pairs (r > |0.7|). The result of this preliminary analysis showed that none of the above habitat cover and structure variables were highly collinear. To model the distribution of species richness and abundance of individuals, count (i.e. number of species or individuals) was modelled specifying a Poisson error distribution and a log link function. Full models were subject to a model reduction procedure whereby non-significant terms were sequentially dropped from a model until only significant terms remained. Since initial models of abundance of individuals showed over-dispersion, we used a quasi-Poisson error distribution model.
In addition to the full model which tested habitat cover, habitat structure and plot type as covariates, we ran separate models with only habitat cover and structure variables (i.e. percentage of shrub, stone-rock, soil-rubble and grass cover, H′ of vegetation cover, mean height and CV of the vertical component of the habitat and relative heterogeneity) or only plot type (i.e. natural grassland, non-restored and restored ski-pistes), to verify through Akaike’s Information Criterion (AIC) which set of variables carried more information. GLMs were calculated with R 15.0.1 (R Core Team Citation2014).
Results
We conducted surveys at 49 independent plots ranging in altitude from 1478 to 2250 m a.s.l. (mean: 1812 ± 161.5 m). Thirty-two species were detected, of which 24 were grassland or ecotone species and eight were shrub or woodland species (online supplementary material Appendix 1). Whinchat Saxicola rubetra, Water Pipit Anthus spinoletta, Black Redstart Phoenicurus ochruros and Linnet Linaria cannabina were the most common species, with frequencies of occurrence (the number of times species were observed within 98 samples) higher than 10%.
Differences between plot types
Habitat
Non-restored ski-pistes showed a significantly lower percentage of grass cover and average vertical structure and a higher soil-rubble cover than natural grassland and restored ski-pistes. No significant habitat difference between restored ski-pistes and natural grassland was found (). Shrubs (especially rhododendron) were found in natural grasslands only (a mean ± sd cover of 4.58 ± 13.67%), because they were removed from ski-pistes during construction.
Table 1. Mean ± sd habitat cover and structure variables, bird species richness and abundance of individuals in natural grasslands, non-restored and restored ski-pistes. Interplot differences were tested with a one-way anova. Least significant difference post-hoc tests were used for pairwise comparisons of means.
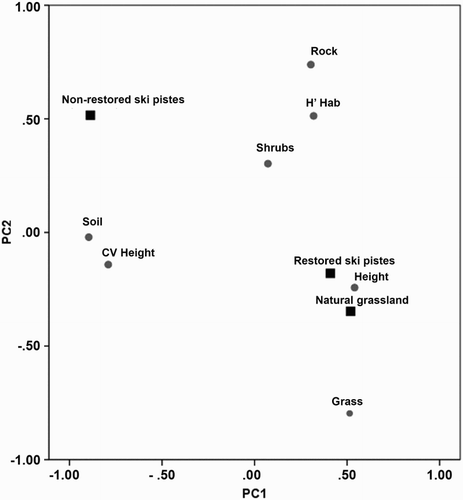
FA showed that the first two principal components (PC1, PC2) accounted for 70.06% of the total variation in the habitat structure matrix, with eigenvalues > 1. The percentage of soil-rubble and the heterogeneity of the vertical component had a negative contribution on PC1 (suggesting a gradient of grass cover from bare ground to pastures) and the percentage of rocks and Shannon index of vegetation cover provided the major positive loading on PC2 (suggesting a gradient from grassland-dominated plots to more diversified plots with shrubs and rocks). The relative position of centroids (i.e. the average location of survey plots in ordination space) in the biplot determined by the first two principal components showed that natural grassland and restored ski-pistes were structurally quite similar, and very different from the non-restored ski-piste category, which was identified mainly with soil-rubble cover ().
Figure 1. Biplot of a principal component analysis (PC1 vs. PC2) where both environmental descriptors and survey plots are plotted together. As a matter of clarity, to avoid plotting too many confounding points (i.e. 49 survey plots plus 7 descriptors), the distribution of survey plots for each plot type is synthetically represented by centroids (i.e. the weighted mean of survey plots). Note: Soil: percentage of soil-rubble cover; Rock: percentage of stone-rock cover; Shrub: percentage of shrub cover; Grass: percentage of grass cover; H’hab: Shannon diversity of the habitat; CV Height: heterogeneity of the vertical component; Height: mean height of the vertical component. Dots indicate environmental descriptors (habitat structure variables) and squares indicate centroids of survey plots.
Bird communities
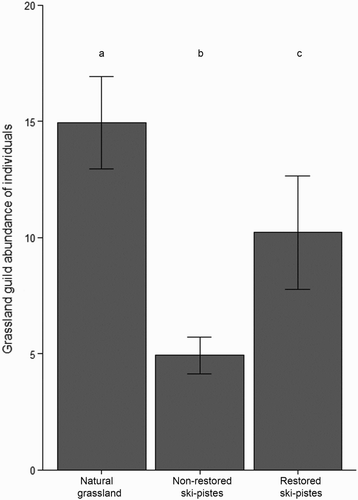
Non-restored ski-pistes, restored ski-pistes and natural grassland plots showed significant differences in terms of bird species richness and abundance of individuals (). Plots located in natural grasslands supported the highest bird species richness and abundance, whereas those set in non-restored ski-pistes had the lowest values. However, differences between restored ski-pistes and natural grasslands depended on whether all species or only grassland species were taken into account. When all species were considered, post-hoc tests showed that species richness was not significantly different between plots in natural grasslands and those in restored ski-pistes (even though values were lower in the latter), whilst the abundance of individuals was significantly greater in natural grassland. When only grassland species were considered, pairwise post-hoc tests showed that both ecological parameters (species richness and abundance) were significantly lower on restored ski-pistes than in natural grassland (, & ).
Figure 2. Mean species richness of grassland species per point count in natural grasslands, non-restored ski-pistes and restored ski-pistes. Bars are standard errors.
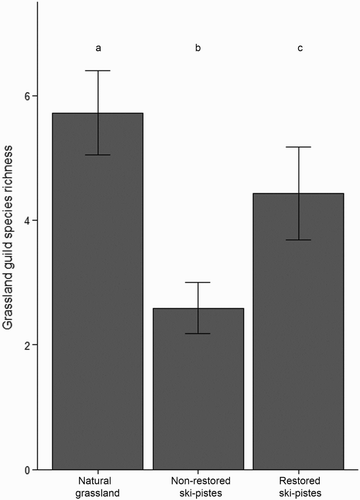
Figure 3. Mean number of individuals of grassland species per point count in natural grasslands, non-restored ski-pistes and restored ski-pistes. Bars are standard errors.
Relationships between bird community and habitat
GLMs showed that bird community parameters (species richness and abundance) of both non-restored and restored ski-pistes were significantly lower than those of natural grassland (reference category) independently of the community composition (). Notably, differences in the abundance of individuals of grassland species of restored ski-pistes and of natural grasslands were highly significantly different (P < 0.001), whereas the differences regarding the overall community were less striking (P < 0.05).
Table 2. Summary of GLM of (a) bird species richness and abundance of individuals of the overall community, and of (b) species richness and abundance of individuals of grassland species in relation to non-restored ski-pistes, restored ski-pistes and natural grasslands (reference level), habitat parameters and altitude.
Bird communities were also influenced by two habitat structure variables, shrub cover and vegetation diversity; grassland species were also positively influenced by altitude. As a rule, community parameters increased with vegetation diversity (3 instances out of 3) and decreased with shrub cover (3 out of 4). In general, separate models with only plot type carried more information than those with only habitat cover and structure variables (Total species richness: plot type only model AIC 233.64, vegetation only model AIC 254.54; Grassland species richness: plot type only model AIC 223.91, vegetation only model AIC 233.66; Abundance of grassland species: plot type only model AIC 325.53, vegetation only model AIC 385.17).
Discussion
The efforts to restore ski-pistes have changed considerably since the demands for sustainable erosion control arose in the 1970s. Restoration technology has made considerable progress in recent years, and specific revegetation measures are available that make use of local seeds and plants that are adapted to and suited for any elevation. In the long-term, in fact, sufficient protection against erosion can only be guaranteed if stable, enduring and ecologically adapted sub-alpine and alpine plant species become established (Klug et al. Citation2013, Krautzer et al. Citation2013, Rixen Citation2013). Nevertheless, several thousand kilometres of ski-pistes still require restoration in the European Alps. Above the tree line in particular, vegetation cover on the ski-pistes remains extremely low for long periods after restoration (at least 10–12 years), despite the use of modern techniques such as hydroseeding (Barni et al. Citation2007). Worse still, the vegetation cover on high altitude machine-graded pistes may deteriorate over time, illustrating that natural recovery may not occur in these managed alpine habitats (Roux-Fouillet et al. Citation2011). Only at elevations of several hundred meters below the tree line does re-establishment of vegetation occur rapidly and reliably (Rixen Citation2013).
Despite the amount of research conducted, very little is known about the effect of ski-piste restoration on animal communities. To our knowledge, no study on the effect of restoration on bird communities has been published so far. In this paper, we therefore tested for the first time the hypothesis that revegetation of ski-pistes of open habitat zones goes hand-in-hand with bird community recovery. In terms of habitat structure, restored ski-pistes were in fact not significantly different from natural grassland, thus suggesting that a successful restoration level was achieved.
Previous studies carried out on non-restored ski-pistes in open habitat areas have shown that plots located in natural grasslands supported the greatest bird species richness and diversity and the greatest grassland species density, whereas those in ski-pistes had the lowest values; moreover, plots located beside ski-pistes did not support smaller numbers of bird species and diversity than plots in natural areas, but they supported a significantly lower bird density (Rolando et al. Citation2007). There was no difference in the overall bird community between restored ski-pistes and natural grasslands, but there was a significant difference in grassland specialist species, suggesting that habitat quality for this group in particular is affected by ski-piste type. More broadly, these results show that the guild approach can reveal patterns not evident when considering the community as a whole (Bishop & Mayers Citation2005, Caprio et al. Citation2008).
The present results suggest that restoration of ski-pistes may partially promote the recovery of local bird communities. Species richness and abundance of birds were in fact significantly higher on restored than on non-restored ski-pistes, independently of the species considered and the analyses carried out. Nevertheless, bird community parameters (especially those of grassland species) of restored ski-pistes were still lower than those of natural grassland, despite the fact that the presence of shrubs (i.e. rhododendrons) in grasslands tended to lower bird diversity. This suggests therefore that an apparently successful restoration of ski-pistes may not be enough to promote a complete recovery of bird communities. These results are likely driven by the vegetation of the restored ski-pistes. Several studies have demonstrated that grass cover of ski-pistes is a major determinant of local animal diversity (Negro et al. Citation2010, Caprio et al. Citation2011). However, equal grass cover of restored ski-pistes and natural grassland does not necessarily prove they are ecologically equivalent, because differences may still be great and significant in terms of density of grass species, which remains lower on restored ski-pistes, or in terms of occurrence of alien plant species, which is higher on ski-pistes, especially when hydroseeded (Barni et al. Citation2007). This means that, even though grass cover is high, the vegetation of restored ski-pistes remains different from that of adjacent pastures and, therefore, less attractive to local birds. Previous studies have shown that responses of ground-dwelling arthropods to ski-piste restoration (as a consequence of hydroseeding with commercial mixtures) were contrasting. Restored ski-pistes were colonized by grasshoppers Orthoptera, which were more abundant on ski-pistes than on the adjacent grassland plots, but ski-pistes and adjacent grassland plots were used equally by ground beetles Carabidae, and ski-pistes were avoided by spiders Araneae (Negro et al. Citation2013). Birds feeding on epigeic invertebrates might be influenced by these changes (e.g. Alpine Choughs Pyrrhocorax graculus were seen feeding on grasshoppers in ski-restored ski-pistes), but present results suggest that overall attractiveness of restored ski-pistes was lower than that of natural grassland.
This study indicates, therefore, that restoration of ski-pistes of open habitat areas may promote the complete recovery of local bird communities only if an integral recolonization of the original vegetal communities (which is essential to host invertebrates) is achieved. Otherwise, the recovery will be significant (e.g. higher values of bird species richness and abundance than non-restored ski-pistes) but nevertheless, partial (e.g. lower values of species richness and abundance than natural grassland). Considering that thousands of kilometres of ski-pistes have been already constructed in the Alps, and that climate change will probably increase the potential conflict between skiing and high-elevation bird species, the best conservation choice will be that of abstaining from, or at least deferring, the construction of new ski-pistes. If construction of new ski-pistes is unavoidable, it is vitally important that restoration measures follow restoration guidelines that represent today’s state-of-the-art (Rixen Citation2013) and that original grasslands which are compatible with skiing activities are preserved.
We wish to stress that successful restoration of ski-pistes will not solve eventual problems connected with snow cover management. Snow cover of ski-pistes is very different from that of natural grassland because of the use of artificial snow produced by snow-making facilities and/or the snow compression caused by skiers and heavy machinery. In both cases, the main effect is that of postponing the time of melt-out (Rixen Citation2013). This causes a delay in vegetative growth and flowering, which has been demonstrated to affect alpine butterfly communities (Rolando et al. Citation2013). Hence, it cannot be excluded that, all other things being equal, this phenological delay may also affect bird communities, irrespective of the restoration status of the ski-pistes. Even nature-friendly management does not necessarily guarantee animal conservation. In ski-pistes of an alpine open habitat zone whose vegetation cover never experienced any disturbance, a noticeable decline in the abundance of most epigeic beetle species in patches with artificially increased accumulation of snow was found (Kašák et al. Citation2013).
Much published research (as here) is based on models that take into account habitat and vegetation cover (Caprio et al. Citation2011, Chamberlain et al. Citation2013). This kind of approach is adequate in depicting and identifying major responses and processes, but much effort should also be made in order to better understand underlying mechanisms, including fine-scale studies of the relationship between birds, vegetation, snow cover and invertebrate availability. This in-depth analysis is needed to drive management interventions to improve habitat conditions in alpine areas which are also already threatened by climate change (Chamberlain et al. Citation2013). This suggests that, in addition to research on the effects of ski-restoration, more studies on the effect of winter snow on birds and invertebrate communities of ski-pistes are desirable. For example, species such as Alpine Chough, Red-billed Chough Pyrrhocorax pyrrhocorax and Snowfinch Montifringilla nivalis are dependent on invertebrates during the spring and summer, whose availability is likely to be affected by vegetation structure and snow cover. The effect of snow melt (including artificial snow) on plant and invertebrate phenology, both on and off pistes, and consequently feeding ecology of high altitude grassland birds, should be a priority research topic in this field.
Supplementary Appendix
Download MS Word (14.4 KB)Acknowledgements
We thank Sonja Wipf and one anonymous reviewer for their comments on earlier drafts of the manuscript, and Luca Giraudo and Marta De Biaggi for their support.
ORCiD
Enrico Caprio http://orcid.org/0000-0002-5997-5959
Dan Chamberlain http://orcid.org/0000-0002-5381-2024
Additional information
Funding
Related Research Data
References
- Amacher-Hoppler, A. & Schoch, R. 2008. Remontées Mécaniques Suisses: Faits et Chiffres. Remontées mécaniques, Berne.
- Barni, E., Freppaz, M. & Siniscalco, C. 2007. Interactions between vegetation, roots, and soil stability in restored high-altitude ski runs in the Alps. Arct. Antarc. Alp. Res. 39: 25–33. doi: 10.1657/1523-0430(2007)39[25:IBVRAS]2.0.CO;2
- Bayfield, N.G. 1996. Long-term changes in colonization of bulldozed ski pistes at Cairn Gorm, Scotland. J. Appl. Ecol. 33: 1359–1365. doi:10.2307/2404776.
- Bicknell, S. & Mcmanus, P. 2006. The canary in the coalmine: Australian ski resorts and their response to climate change. Geogr. Res. 44: 386–400. doi:10.1111/j.1745-5871.2006.00409.x.
- Bishop, J.A. & Mayers, W.L. 2005. Associations between avian functional guild response and regional landscape properties for conservation planning. Ecol. Indic. 5: 33–48. doi:10.1016/j.ecolind.2004.10.001.
- Caprio, E., Ellena, I. & Rolando, A. 2008. Assessing habitat/landscape predictors of bird diversity in managed deciduous forests: a seasonal and guild-based approach. Biodivers. Conserv. 18: 1287–1303. doi:10.1007/s10531-008-9478-1.
- Caprio, E., Chamberlain, D.E., Isaia, M. & Rolando, A. 2011. Landscape changes caused by high altitude ski-pistes affect bird species richness and distribution in the Alps. Biol. Conserv. 144: 2958–2967. doi:10.1016/j.biocon.2011.08.021.
- Cernusca, A., Angerer, H., Newesely, C. & Tappeiner, U. 1990. Auswirkungen von Kunstschnee – Eine Kausalanalyse der Belastungsfaktoren. Verhandlungen der Gesellschaft für Ökologie 19: 746–757.
- Chamberlain, D.E., Negro, M., Caprio, E. & Rolando, A. 2013. Assessing the sensitivity of alpine birds to potential future changes in habitat and climate to inform management strategies. Biol. Conserv. 167: 127–135. doi:10.1016/j.biocon.2013.07.036.
- Delarze, R., Gonseth, Y. & Galland, P. 1998. Guide des milieux naturels de Suisse: écologie-menaces-espèces caractéristiques. Delachaux et Niestle, Lausanne.
- Elsasser, H. & Messerli, P. 2001. The vulnerability of the snow industry in the Swiss Alps. Mt. Res. Dev. 21: 335–339. doi: 10.1659/0276-4741(2001)021[0335:TVOTSI]2.0.CO;2
- Fukushima, T., Kureha, M., Ozaki, N., Fujimori, Y. & Harasawa, H. 2002. Influences of air temperature change on leisure industries – case study on ski activities. Mitigation Adapt. Strateg. Glob. Chang. 7: 173–189. doi:10.1023/A:1022803405470.
- Gaunch, H.G. Jr 1984. Multivariate Analysis in Community Ecology. Cambridge University Press, Cambridge.
- Hadley, G.L. & Wilson, K.R. 2004. Patterns of small mammal density and survival following ski-run development. J. Mammal. 85: 97–104. doi: 10.1644/1545-1542(2004)085<0097:POSMDA>2.0.CO;2
- Isselin-Nondedeu, F. & Bédécarrats, A. 2007. Influence of alpine plants growing on steep slopes on sediment trapping and transport by runoff. CATENA 71: 330–339. doi:10.1016/j.catena.2007.02.001.
- Kašák, J., Mazalová, M., Šipoš, J. & Kuras, T. 2013. The effect of alpine ski-slopes on epigeic beetles: does even a nature-friendly management make a change? J. Insect. Conserv. 17: 975–988. doi:10.1007/s10841-013-9579-3.
- Keßler, T., Cierjacks, A., Ernst, R. & Dziock, F. 2012. Direct and indirect effects of ski run management on alpine Orthoptera. Biodivers. Conserv. 21: 281–296. doi:10.1007/s10531-011-0184-z.
- Klug, B., Markart, G., Meier, J., Krautzer, B. & Kohl, B. 2013. Ski run re-vegetation: a never-ending story of trial and error? In Rixen, C. & Rolando, A. (eds.) The Impacts of Skiing and Related Winter Recreational Activities on Mountain Environments, 155–183. Bentham Science, Sharjah.
- Koenig, U. & Abegg, B. 1997. Impacts of climate change on winter tourism in the Swiss Alps. J. Sustainable Tour. 5: 46–58. doi:10.1080/09669589708667275.
- Körner, C. 2003. Alpine Plant Life. Springer, Heidelberg.
- Krautzer, B., Graiss, W. & Klug, B. 2013. Ecological restoration of ski runs. In Rixen, C. & Rolando, A. (eds.) The Impacts of Skiing and Related Winter Recreational Activities on Mountain Environments, 184–209. Bentham Science, Sharjah.
- Laiolo, P. & Rolando, A. 2005. Forest bird diversity and ski-runs: a case of negative edge effect. Anim. Conserv. 8: 9–16. doi:10.1017/S1367943004001611.
- Mackenzie, D. 1989. Alpine Countries Seek Controls on Skiers, Builders and Roads. New Scientist Publ Expediting Inc 200 Meacham Ave, Elmont, NY.
- Mosimann, T. 1985. Geo-ecological impacts of ski piste construction in the Swiss Alps. Appl. Geogr. 5: 29–37. doi:10.1016/0143-6228(85)90004-9.
- Negro, M., Isaia, M., Palestrini, C. & Rolando, A. 2009. The impact of forest ski-pistes on diversity of ground-dwelling arthropods and small mammals in the Alps. Biodivers. Conserv. 18: 2799–2821. doi:10.1007/s10531-009-9608-4.
- Negro, M., Isaia, M., Palestrini, C. & Rolando, A. 2010. The impact of high-altitude ski pistes on ground-dwelling arthropods in the Alps. Biodivers. Conserv. 19: 1853–1870. doi:10.1007/s10531-010-9808-y.
- Negro, M., Novara, C., Bertolino, S. & Rolando, A. 2012. Ski-pistes are ecological barriers to forest small mammals. Eur. J. Wildl. Res. 59: 57–67. doi:10.1007/s10344-012-0647-x.
- Negro, M., Rolando, A., Barni, E., Bocola, D., Filippa, G., Freppaz, M. & Isaia, M. 2013. Differential responses of ground dwelling arthropods to ski-piste restoration by hydroseeding. Biodivers. Conserv. 22: 2607–2634. doi:10.1007/s10531-013-0544-y.
- Patthey, P., Wirthner, S., Signorell, N. & Arlettaz, R. 2008. Impact of outdoor winter sports on the abundance of a key indicator species of alpine ecosystems. J. Appl. Ecol. 45: 1704–1711. doi:10.1111/j.1365-2664.2008.01547.x.
- Pechlaner, H. & Tschurtschenthaler, P. 2003. Tourism policy, tourism organisations and change management in Alpine regions and destinations: a European perspective. Curr. Issues Tour. 6: 508–539. doi:10.1080/13683500308667967.
- R Core Team. 2014. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/.
- Rixen, C. 2013. Skiing and vegetation. In Rixen, C. & Rolando, A. (eds.) The Impacts of Skiing and Related Winter Recreational Activities on Mountain Environments, 65–78. Bentham Science, Sharjah.
- Rixen, C., Haeberli, W. & Stoeckli, V. 2004. Ground temperatures under ski pistes with artificial and natural snow. Arct. Antarc. Alp. Res 36: 419–427. doi: 10.1657/1523-0430(2004)036[0419:GTUSPW]2.0.CO;2
- Rolando, A., Caprio, E., Rinaldi, E. & Ellena, I. 2007. The impact of high-altitude ski-runs on alpine grassland bird communities. J. Appl. Ecol. 44: 210–219. doi:10.1111/j.1365-2664.2006.01253.x.
- Rolando, A., Negro, M., D’Entrèves, P.P., Balletto, E. & Palestrini, C. 2013. The effect of forest ski-pistes on butterfly assemblages in the Alps. Insect Conserv. Divers. 6: 212–222. doi:10.1111/j.1752-4598.2012.00204.x.
- Roux-Fouillet, P., Wipf, S., & Rixen, C. 2011. Long-term impacts of ski piste management on alpine vegetation and soils. J. Appl. Ecol. 48: 906–915. doi:10.1111/j.1365-2664.2011.01964.x.
- Sanecki, G.M., Cowling, A., Green, K., Wood, H. & Lindenmayer, D. 2006. Winter distribution of small mammals in relation to snow cover in the subalpine zone, Australia. J. Zool. (London) 269: 99–110. doi:10.1111/j.1469-7998.2006.00074.x.
- Sato, C.F., Wood, J.T. & Lindenmayer, D.B. 2013. The effects of winter recreation on alpine and subalpine fauna: a systematic review and meta-analysis. PLoS ONE 8: e64282. doi:10.1371/journal.pone.0064282.
- Sato, C.F., Wood, J.T., Schroder, M., Green, K., Osborne, W.S., Michael, D.R. & Lindenmayer, D.B. 2014a. An experiment to test key hypotheses of the drivers of reptile distribution in subalpine ski resorts. J. Appl. Ecol. 51: 13–22. doi:10.1111/1365-2664.12168.
- Sato, C.F., Wood, J.T., Schroder, M., Michael, D.R., Osborne, W.S., Green, K. & Lindenmayer, D.B. 2014b. Designing for conservation outcomes: the value of remnant habitat for reptiles on ski runs in subalpine landscapes. Landscape Ecol. 29: 1225–1236. doi:10.1007/s10980-014-0058-3.
- Scott, D., Dawson, J. & Jones, B. 2007. Climate change vulnerability of the US Northeast winter recreation – tourism sector. Mitig. Adapt. Strateg. Glob. Change. 13: 577–596. doi:10.1007/s11027-007-9136-z.
- Simons, P. 1988. Après ski le déluge. New Scientist. 117: 49–52.
- Titus, J.H. & Tsuyuzaki, S. 1999. Ski slope vegetation of Mount Hood, Oregon, U.S.A. Arct. Antarc. Alp. Res. 31: 283–292. doi:10.2307/1552259.
- Urbanska, K.M. & Fattorini, M. 2000. Seed rain in high-altitude restoration plots in Switzerland. Restor. Ecol. 8: 74–79. doi:10.1046/j.1526-100x.2000.80010.x.
- Verner, J. 1984. The guild concept applied to management of bird populations. Env. Manag. 8: 1–13. doi:10.1007/BF01867868.
- Wipf, S., Rixen, C., Fischer, M., Schmid, B. & Stoeckli, V. 2005. Effects of ski piste preparation on alpine vegetation. J. Appl. Ecol. 42: 306–316. doi:10.1111/j.1365-2664.2005.01011.x.