
ABSTRACT
Capsule: House Sparrows Passer domesticus in the Valencian Community, where the species is declining, were more abundant in parks and urban areas where rubbish bins are available as a source of food. In heavy traffic areas, crowded streets and high-rise building zones, the species was less abundant.
Aims: To explore biotic and abiotic factors affecting the abundance of urban House Sparrows. To infer the causes that might explain the urban decline and to propose measures that might halt it and elevate population numbers.
Methods: The abundance of urban House Sparrows was analysed in relation to 17 landscape variables by carrying out 181 point counts (50 urban parks/38 schools/93 streets) per season across six locations and over five seasons (breeding and non-breeding seasons) in southeast Spain. Urban parks were also described by 21 additional variables. Hierarchical partitioning analyses were used to identify favoured or avoided urban factors by birds in each season.
Results: House Sparrows were most abundant in parks and other urban areas where rubbish bins were available as their foraging sites. The presence of rubbish bins, area and proximity to other parks favoured their abundance in parks. Furthermore, House Sparrows were less abundant in crowded streets, heavy traffic areas and high-rise buildings zones.
Conclusion: House Sparrows were present in high abundance where they feed but were scarce in polluted, crowded and heavily built-up areas of the city. Changes in the structure of these urban habitats may underlie the documented declines across Spain. Green urban planning and management are needed to ensure heterogeneous green areas with enough vegetation coverage to provide high availability of natural food versus anthropogenic food scraps. Sufficient numbers of buildings offering nest sites and reduced air pollution might ameliorate cities adequately to support larger populations of House Sparrows.
The House Sparrow Passer domesticus has been associated with humans since the Bronze Age (Ericson et al. Citation1997) and presents the widest distribution of any species in the world (Cramp & Perrins Citation1994, BirdLife International Citation2016). This is a consequence of an anthropogenic process, namely releases of birds into non-native areas and is, at least initially, partly also a consequence of its dependency and co-evolvement with human agricultural development (Sætre et al. Citation2012, Liebl et al. Citation2015). However, this species has declined in the Western Palaearctic during the last quarter of the 20th century (Summers-Smith Citation2003) and at the beginning of the 21st century (Raven et al. Citation2003, Hayhow et al. Citation2014). Moreover, these declines have occurred in both urban and farmland areas (Summers-Smith Citation2000, Crick et al. Citation2002, De Laet & Summers-Smith Citation2007), even though the decline first started in rural ones (Summers-Smith Citation2003, Robinson et al. Citation2005).
Their decline was first detected in Great Britain (Summers-Smith Citation2003) where 65% of birds have been lost in 32 years, between 1970 and 2012 (Hayhow et al. Citation2014). For instance, the species declined in London by 71% between 1994 and 2002 (Raven et al. Citation2003). On the other hand, it is still abundant in Spain (Carrascal & Palomino Citation2008), despite showing an annual decline of 0.9% (SEO/BirdLife Citation2013). However, trends in Valencian populations mirror those in Great Britain. The first signs of decline were detected in orange groves of Sagunto (Valencia) where the population decreased by more than 90% between 1975 and 2001 (Gil-Delgado et al. Citation2002). Later, in the parks of the city of Valencia, the population declined by 70% between 1998 and 2008, equivalent to an average annual decline of 15% (Murgui & Macias Citation2010). In addition, large-scale studies within the Valencian Community detected an overall decline of 22% between 1998 and 2015 (Vera & Escandell Citation2016).
Human-dependent bird species are more likely to display negative population trends in the most heavily human-modified environments (Siriwardena et al. Citation1998). Accordingly, it is appropriate to investigate declines in House Sparrow numbers in relation to patterns of temporal changes in human activities (Anderson Citation2006). Several hypotheses have been proposed to explain their declines in urban areas including: predation by Domestic Cats Felis silvestris catus or Eurasian Sparrowhawks Accipiter nisus (Woods et al. Citation2003, Bell et al. Citation2010); competition for food resources with Feral Rock Doves Columbia livia; and competition with European Starlings Sturnus vulgaris and Spotless Starlings Sturnus unicolor for nest sites (Weitzel Citation1988). Furthermore, it has also been attributed to other causes such as: structural habitat changes by the improvement of human socioeconomic status in towns and cities which means loss of green space, invertebrate and nest availability sites (McKinney Citation2002, Shaw et al. Citation2008, Mouldrá et al. Citation2018); increased exposure to electromagnetic radiation (Balmori & Hallberg Citation2007); poor-quality trophic resources availability in urban areas (Herrera-Dueñas et al. Citation2015) and increased pollution from traffic (Summers-Smith Citation2007, Herrera-Dueñas et al. Citation2017). The most favoured hypothesis to explain the decline of House Sparrows in urban areas is that it is down to a combination of factors rather than to a single one. Nevertheless, we are still some way from identifying these key urban factors (Vincent Citation2005, Anderson Citation2006).
A very useful approach to identify such factors may be provided by a fine-scale habitat study of areas where the sparrow is declining (Robinson et al. Citation2005, Chamberlain et al. Citation2007). Therefore, here, we aimed to explore the key biotic and abiotic factors affecting the abundance of House Sparrows in several locations in Valencia across different seasons. Most previous research studied the species just within its breeding season (e.g. Chamberlain et al. Citation2007, Shaw et al. Citation2011), whereas we studied birds in early and midsummer (between late June and July, late breeding season in Spain which is from mid-March to early August (Gil-Delgado et al. Citation1979, Bernis Citation1989)) and autumn and winter (their two non-breeding seasons in the study area). Taking into account that summer counts could be biased towards dispersing juvenile House Sparrows present in areas not selected by adults, it would also be interesting to know these important factors for conservation purposes of urban populations of sparrows. From previous studies in Valencia (Murgui Citation2009) and Great Britain (Shaw Citation2009), we predicted some factors determining bird abundance would be related to the availability of food and nest sites. However, the species has declined throughout the whole of eastern Spain (Vera & Escandell Citation2016). Therefore, we explored habitat factors in coastal and inland areas of the southeastern Iberian Peninsula as a whole to increase our ability to identify factors that might explain general declines in this species’ abundance. We hope that our findings may inform conservation policy to halt and reverse its decline.
Methods
Study area and bird census
The study was carried out in five locations in the Valencia area in southeastern Spain (four located in the northern area of Alicante’s province and one in the southeast of Valencia’s province) ( and ). This last location was split across two sites, Gandía and Grao/Playa, separated by orange groves ( and ). Further information about human demographics, agriculture and industry of these locations is provided by Jordà (Citation1976), Banyuls (Citation1985) and Tormo (Citation2011).
Figure 1. Maps showing selected Valencian localities (Spain, Europe) for the study of factors affecting the abundance of House Sparrows in urban areas of southeast of Spain between 2014 and 2016. The sizes of the circles show how large the urban areas are in terms of their overall number of inhabitants.
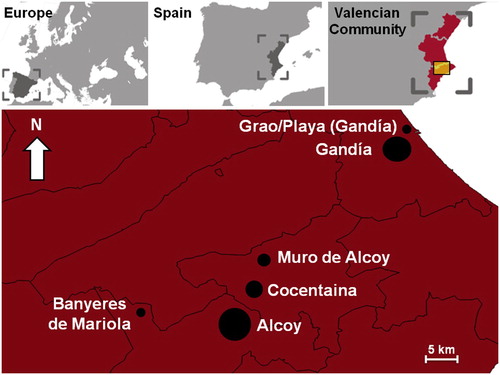
Table 1. Geographical, demographical and climatic characteristics of the selected locations for the study of factors affecting the abundance of House Sparrows in urban areas of southeast of Spain between 2014 and 2016. Table devised with data from SigPac (Citation2014), AVAMET (Citation2016) and Instituto Nacional de Estadística (Spanish National Statistics Institute) (Citation2016).
House Sparrow abundance was determined using point counts (Ralph et al. Citation1996, Gibbons & Gregory Citation2006). To avoid double counting of birds, point counts were separated by a minimum distance of 100 m. At each point count, abundance of birds within a space limited by a radius of 50 m was recorded by a single observer (EB-P) for 5 minutes. Birds were recorded by sight and vocalizations and double counting was avoided. A settlement period of 1 minute was allowed before each count.
The area of each location was calculated using the cartographic viewer SigPac (SigPac Citation2014). It was divided into several homogeneous parcels in terms of type and age of the buildings. The number of point counts in each homogeneous parcel and thus in each locality was related to its urban surface (). At each parcel, point counts were undertaken on streets, in school grounds and in parks, which were distributed randomly taking into account any inaccessible areas (those in parks and schools were randomly distributed within them) via the QGIS platform (QGIS Citation2014) (). Censusing of sparrows was carried out in the autumn of 2014 and 2015 (non-breeding seasons), the winter of 2014–15 and 2015–16 (non-breeding seasons), and the summer of 2015 (breeding season). During each field season 181 point counts were made, totalling 905. Each counting session lasted 2 hours starting at daily official sunrise when House Sparrows are most active (Anderson Citation2006). Sessions were not conducted on days of rain and/or winds. On average, a point count session comprised 10–14 counts.
Figure 2. Distribution of the 18 point counts within the location of Muro de Alcoy (Valencia, Spain) as an example of their distribution in the locations for the study of factors affecting the abundance of House Sparrows in urban areas of southeast of Spain between 2014 and 2016. Symbols represent point count locations in parks (squares), on streets (circles) and in schools (crosses).

Urban landscape and park variables
At each point count in the urban landscape, 17 variables were described (A), some through a qualitative classification of presence/absence (1/0), and others through a qualitative ranking (0, 1, 2, …) or by quantized values (0 to n). These 17 landscape variables were grouped into three classes: urban structure, human factors and food resources (A). Variables were selected paying particular attention to human-related urban factors, infrastructure and planning, which could be rather easily addressed and managed according to species’ requirements. In the resources group, seven variables related to presence of anthropogenic scrap food sources and natural food sources for sparrows were measured (Smith et al. Citation2006, Murgui Citation2009, Rega-Brodsky & Nilon Citation2016, Skórka et al. Citation2016) (A). In towns and cities, food could be in short supply and these variables may be essential for sparrow survival. The second group contained six variables related to urban structure (Murgui Citation2007, Shaw et al. Citation2008), nest sites, shelter (Savard & Falls Citation1981, Indykiewicz Citation1991, Wotton et al. Citation2002) and urban movements of birds (Bernis Citation1989, Anderson Citation2006) (A). The third group included four variables related to the human influences that can promote or depress House Sparrow numbers, such as traffic, pedestrians, socioeconomic status and altitude above sea level (Summers-Smith Citation2007, Shaw et al. Citation2008, Menon et al. Citation2013) (A).
Table 2. Summary of the environmental variables used in the Hierarchical Partitioning Analyses for the study of factors affecting the abundance of House Sparrows in urban areas of southeast of Spain between 2014 and 2016. Note that there are two different main groups of studied variables, (A) urban landscape and (B) parks. The variable geography was included in every group to control overall location effects of the sampling points.
Parks were also described according to 21 variables in four groups (B). The first contained variables related to vegetation and substrate while the second group described aspects of the location of the park (B). The third included variables related to resources provided by humans that might influence bird abundance in parks, such as rubbish bins, terraces, children’s play areas or water bodies (B), while the fourth group described variables related to urban factors including the influence of people, socioeconomic status, cats and dogs (B).
Statistical analysis
We used hierarchical partitioning (HP) to identify the set of predictor variables that best explained the abundance of House Sparrows in each season (Chevan & Sutherland Citation1991, Mac Nally Citation1996). HP does not aim to identify a best regression model as such but rather uses all models in a regression hierarchy to distinguish those variables that have high independent correlations with the dependent variable. Such variables are likely to be most influential in controlling variation in the dependent variable (Mac Nally Citation1996). Their explanatory power is segregated into the independent effect (I) and the effects caused jointly with other variables (J) (Mac Nally Citation2002). It also offers a set of negative effects that contains variables that act as suppressors of other variables, suppressing irrelevant variance of independent variables, thus enhancing the relationship between the other independent variables and the dependent one (Chevan & Sutherland Citation1991). Moreover, model outputs also provide the percentage (I%) of the total group I counted in each variable.
Hierarchical partitioning was performed with the ‘hier.part’ package in RStudio 0.99.903 (Walsh & Mac Nally Citation2015) using Poisson regression and r-squared (Rsqu) as the goodness-of-fit measure (Jongman et al. Citation1995). The significance of the contribution of environmental variables was evaluated by 999 tests based on randomizations (Mac Nally Citation2002), where the random distribution of I was generated. For the variable to make a significant contribution, the observed I had to be within the 95-percentile with Z-scores ≥ 1.65 (Hallstan et al. Citation2010).
We opted to analyse data from each single season by means of a separate model, to evaluate whether similar or different environmental factors regulated House Sparrow abundance over the same sites in different periods, according to coherent or varying patterns. This approach, coupled with the use of hierarchical partitioning within each season, would help to find stronger and more important effects.
Due to the division of the landscape variables into three groups (A), we avoided performing the HP with more than nine independent variables because higher numbers of variables produce minor rounding errors (Walsh & Mac Nally Citation2015). Prior to HP analyses of each group of variables in each season, Poisson regression models with the variables of each group were carried out using the ‘glm’ function with the canonical link ‘log’ in RStudio 0.99.903 and the variance inflation factor (VIF) among variables was calculated to detect multicollinearity, using the function ‘vif’ of the package ‘car’. When VIF was >5 in some variable, we considered multicollinearity to be high (Zuur et al. Citation2010) and in such cases the variable of the highest VIF or the second one, if the first one was considered of key biological relevance, was discarded. This proceeding was repeated to test if the reduced group of variables showed multicollinearity again.
In each of the five seasons, one HP analysis was performed for each urban landscape group: urban structure, human factors and resources (A). The overall location effects of the sampling points were controlled by including a geographical variable in all of the analyses (longitude × latitude) calculated from the UTM coordinates of each point count (López-Iborra et al. Citation2011). Significant variables from each urban landscape group analysis were then combined in a final HP analysis that assembled selected variables hierarchically for each season. The final HP analysis only retained those key variables for the sparrow abundance. Once again, prior to this, VIF values of selected variables of each group in each season were calculated to avoid multicollinearity. Finally selected categorical variables in the combined HP analysis were analysed to find those significant categories for House Sparrow abundance. This was conducted in RStudio 0.99.903 through the functions ‘kruskal.test’ and ‘kruskalmc’ of the ‘pgirmess’ package which reduce the probability of making a Type I error (Field et al. Citation2012). Prior to this, normality and homogeneity of variances of those selected variables were analysed by performing tests of Kolmogorov–Smirnoff and Levene in RStudio 0.99.903 with the functions ‘ks.test’ and ‘leveneTest’ from the packages ‘stats’ and ‘car’, respectively. Poisson regression models were carried out using the ‘glm’ function with the canonical link ‘log’ in RStudio 0.99.903 to determine whether each selected variable was negatively or positively correlated with sparrow abundance in each season. The sign of each category of categorical variables was addressed taking into account their reference level. The same analytical approaches were adopted to examine the variables of the parks (B).
To test climatic differences between both winters, a paired t-test with the minimum monthly temperatures of the localities in December, January and February of both winter seasons (AVAMET Citation2018) was conducted in RStudio 0.99.903 with the ‘t.test‘ function. Prior to this, normality and homogeneity of variances were analysed by performing the test of Shapiro–Wilk (‘shapiro.test‘ function) and F test (‘var.test‘ function).
Results
House Sparrows were present in more than 80% of point counts every season (). Every point count except one (a main street in Alcoy with heavy traffic) yielded House Sparrows in at least one season but there was great variation across them ().
Table 3. Presence during counts, percentage of occupied points and mean count of House Sparrows in the 181 point counts during five seasons in urban areas of southeast of Spain between 2014 and 2016.
Urban landscape
Outputs from initial HP analyses are described in online Appendix S1 and Table S1. Outputs from VIF analyses revealed that multicollinearity among selected variables was only present in summer counts. Presence of parks and trees had the highest values of VIF (6.53 and 3.01, respectively). Therefore, the second one was not included in the combined HP analyses of that season because the presence of a park is a better representation of an urban habitat. Outputs from combined HP analyses of urban landscape variables revealed that in four seasons of the census, urban parks had a positive significant effect on House Sparrow abundance, which was especially important in summer, with an independent effect higher than 31% (). Building height had a negative effect on House Sparrow abundance in every studied season with independent contributions (I%) between 7 and 17% (). The presence of rubbish bins had a positive significant effect on the abundance of birds with a I% between 9% and 25% in both winters and summer (). Geography had an important positive effect on House Sparrow abundance in both winters with I% of 25.47% and 35.25%, respectively. Old buildings and vacant lots had a significant positive effect on the abundance of birds in the first autumn with a I% of 20.55% and 9.29%, respectively. Old buildings were the second most important variable in that autumn after the 32.07% of I% of socioeconomic status (). In addition, the presence of trees was positively selected in the second winter with a I% of 6.91%. However, it had lower explanatory power than urban parks and rubbish bins with I% of 21.63% and 9.89%, respectively (). Both presence of lawn and street width had, respectively, a positive and negative effect of 6% on sparrow abundance during summer. However, they had less explanatory power than the presence of urban parks and rubbish bins with I% of 31.36% and 25.37%, respectively ().
Table 4. Variables identified by the combined Hierarchical Partitioning (HP) Analyses as key factors affecting the abundance of House Sparrows in urban areas of southeast of Spain between 2014 and 2016.
Socioeconomic status had a positive significant effect of 32.07% on the abundance of birds in the first autumn (), with significant differences between their five categories (χ2 2014: = 21.53, df = 4, P < 0.001). Post-hoc tests revealed that significantly more birds were present in deprived areas than in industrial and medium ones.
Number of people had a negative significant effect on sparrow abundance () in both autumns, in the first winter of study and in summer with I% of 10.53%, 18.68%, 13.62% and 5.70%, correspondingly. Traffic volume was also significant and negatively related to bird numbers in both winters and in the second autumn with I% of 11.85%, 10.11% and 13.93%, respectively (). These two variables had a medium-low explanatory power in every season but had a negative significant effect in four and three seasons respectively, which could be considered a regular effect for the abundance of the species.
In regard to joint contributions, most of the variables of both autumns and first winter presented lower joint than independent contributions, suggesting a low degree of correlation between them (). However, summer and the second winter presented the same number of variables with higher independent than joint contributions than variables with higher joint than independent contributions (). This means that these variables had a slight correlation between them. Nevertheless, multicollinearity was avoided by previous VIF analyses and that is the reason for lower joint than independent contributions in most of groups. Also, there are four variables with negative joint contributions in different seasons that act as suppressors of other variables: vacant lot, number of people, rubbish bin and number of vehicles ().
Parks
Outputs from initial HP analyses are provided in online Appendix S2 and Table S2. Multicollinearity was not found between the initial selected variables of the four groups in each season and, consequently, no variables had to be removed to perform the combined HP analyses. Only three seasons had significant variables selected for the abundance of House Sparrows in parks: summer, autumn 2015 and winter 2015–16. In summer, the surface of the park had a positive significant effect on birds of 56.49% of I% while the presence of dogs had a negative significant effect of 43.22% of it (). Moreover, the distance to the nearest park had a negative significant effect on sparrows during the second autumn and winter (). Finally, during the winter 2015–16, the presence of rubbish bins had a positive significant effect of 59.94% of I% on the abundance of birds (). In regard to joint contributions, every selected variable had lower joint than independent contributions, suggesting a low degree of correlation between them and none of them had negative joint contributions. Winter conditions differed between years because means of minimum temperatures of the studied seasons presented significant differences and winter 2015–16 was warmer than winter 2014–15 (Shapiro–Wilk normality text: W = 0.942, P = 0.403/F test to compare two variances: F = 0.796, n = 14, P = 0.675/Paired t-test: t = −3.519, df = 14, P = 0.003).
Table 5. Variables identified by the combined Hierarchical Partitioning (HP) Analyses as key factors affecting the abundance of House Sparrows in urban parks of southeast of Spain between 2014 and 2016.
Discussion
The House Sparrow is still abundant in urban and suburban habitats of the Valencian area as expected due to their close association with humans (Summers-Smith Citation1988, Gil-Delgado & Mora Citation2015), despite their well-documented decline across Europe (Summers-Smith Citation2003, Shaw et al. Citation2008). However, its abundance varied greatly across different habitats within urban areas (Chamberlain et al. Citation2007, Murgui Citation2009) and its presence in almost every studied urban sample was not necessarily a good indicator of healthy populations (Gil-Delgado & Mora Citation2015).
Key environmental factors in the urban landscape
Eight variables had a positive effect on House Sparrow abundance. The most significant ones were the presence of urban parks and rubbish bins, which are sources of food and shelter for birds (Mitschke & Mulsow Citation2003, Bokotey & Gorban Citation2005, Murgui Citation2009). Our results, in agreement with Murgui (Citation2009), showed that urban parks are the main feeding location for House Sparrows in cities of southeastern Spain. Green spaces may offer essential invertebrates for the juvenile sparrows during the breeding season and also plant resources throughout the year (Murgui Citation2009). However, without their careful management, natural food availability is likely to decline rapidly impacting negatively on the species (Peach et al. Citation2008, Peach et al. Citation2015, Weir Citation2015). We think that modern policies of urban green areas (loss of green spaces, excessive cleaning and use of pesticides) may be displacing urban sparrows to sites where food is easily available and/or of lower nutritional value. Contrary to these arguments, Skórka et al. (Citation2016) found that availability of anthropogenic foods had a negative effect on the breeding abundance of House Sparrows mediated through increased predation risks (Lima Citation1998, Krauze-Gryz et al. Citation2013). However, we found that the abundance of House Sparrows was significantly and positively influenced by the presence of rubbish bins, which is a constant and reliable but poor-quality source of human-related food that also represents a sort of proxy for anthropogenic food sources as a whole (Bokotey & Gorban Citation2005, Herrera-Dueñas et al. Citation2015). Our findings suggest the existence of seasonality because it was only not selected in autumn. In the coldest seasons, rubbish bins might be important due to the reliability of food scraps that can be found around them (Bokotey & Gorban Citation2005), while their importance in summer might be due to the easy access to the scraps for juvenile birds. The replacement of open rubbish bins by closed plastic ones has been suggested as a cause of the House Sparrow decline in Ukraine (Bokotey & Gorban Citation2005) and may have influence in the studied area where above-ground rubbish bins are being replaced by underground ones (stainless steel chutes on the pavement that feed into underground chambers). Clearly, anthropogenic food sources represent a double-edge sword to urban birds because while they may promote survival of larger proportions of populations, they lack many key dietary elements such as vitamins, minerals or essential amino acids (Herrera-Dueñas et al. Citation2015).
Variables that just had positive effect on bird abundance once or twice such as the presence of old buildings, presence of vacant lots, socioeconomic status, presence of trees, presence of lawn and geography were not considered as key factors for House Sparrows because of their irregular selection. However, geography was positively and consistently selected during both winters suggesting it as an important factor in this season. House Sparrows were more abundant in coastal locations than in inland areas in winter, as suggested by Gil-Delgado & Mora (Citation2015), and it implies that inland populations should be the priority for study and conservation of the species in the Valencian area. This result could be explained by three non-exclusive hypotheses: larger winter mortality in inland areas (Senar & Copete Citation1995), altitudinal movements (Senar & Borras Citation2004) and/or a differential predator effect (Chamberlain et al. Citation2009, Izquierdo & Sánchez Citation2015), but more research is needed. It is important to note that the positive association of the species to the presence of lawn that we only found in the breeding season reinforces the vision of the need for green urban planning that boosts the availability of invertebrates and supports the urban breeding populations of the species (Weir Citation2015).
On the other hand, four variables were negatively associated with House Sparrow abundance: building height, number of people, traffic volume and street width. Building height emerged as a consistent explanatory variable for reduced sparrow abundance across every season, which suggests that they are negatively linked to heavily built-up areas of the cities, as Murgui (Citation2009) also found in Valencia. Urban House Sparrows feed on the streets, so an increase of building height may increase the vertical distance between feeding areas and roosting sites, probably increasing their cost of foraging. Furthermore, new built-up areas of higher socioeconomic level may provide wholly unsuitable habitats for House Sparrows (Wotton et al. Citation2002, Shaw et al. Citation2008, Mouldrá et al. Citation2018). In agreement with Shaw et al. (Citation2008), we found that, at least in one autumn, socioeconomic status could be an important factor for the species because it was more abundant in deprived areas than in industrial and more developed parts of urban areas.
The number of people and volume of traffic were negatively related to House Sparrow abundance in four and three different seasons respectively, findings contrary to Menon et al. (Citation2013). They did not find any correlation between pedestrians or number of vehicles and House Sparrows abundance, probably due to urban differences between India and Spain. Our findings suggested that the disturbances to birds caused by many people may outweigh their benefits to birds in terms of food availability and the avoidance of streets with dense traffic, which will be the most polluted. This result suggests that pollution in urban areas may be a major driver of House Sparrow decline (Summers-Smith Citation2007). Herrera-Dueñas et al. (Citation2017) suggested that air pollution in cities may be increasing oxidative stress in House Sparrows, entailing negative consequences on the body condition of the individuals and on the viability and future fitness of their offspring, while noise pollution may affect negatively the reproductive fitness of sparrows (Schroeder et al. Citation2012, Meillère et al. Citation2015). A possible prediction for this situation would be that the most polluted cities within the House Sparrow’s range would see the steepest declines in bird numbers, but more research is needed.
The negative association of street width and House Sparrow abundance in summer suggests their preference by narrow streets than wide ones to breed, features that are being lost in new neighbourhoods to favour motor vehicles (European Commission Citation2004).
Variables without effect occurred in every season: distance to the nearest urbanized limit, presence of terraces of bars/cafeterias, presence of schools and presence of private gardens. However, these last three variables stood out because they seemed important sources of human-related food scraps and were never selected. It is possible that food scraps from terraces may not have been available for them at the census time because human intense activity had not started yet in those places. In addition, the terraces of the study area were cleaned at night, reducing the availability of early morning scraps. In line with Magudu & Downs (Citation2015), we found no association between school presence and House Sparrow abundance. Food scraps of school yards may not have been available for them at the census time because the point counts were carried out before break time. Similar anthropogenic resources for sparrows were found to relate negatively to sparrow abundance by Skórka et al. (Citation2016). Furthermore, we found no association between private gardens and sparrow abundance in any season. In Great Britain, private gardens are key habitats for House Sparrows (Chamberlain et al. Citation2007, Shaw et al. Citation2011), so a possible explanation for this difference might be a fundamental lack of such land use in Spain compared with Great Britain (Monclús Citation1998). Additionally, British gardens often have bird feeders, but these are not common in Spain (Chamberlain et al. Citation2005, Reynolds et al. Citation2017).
Key environmental factors in urban parks
Our results showed that park area was positively related to House Sparrow abundance. Contrary to our findings, Murgui (Citation2009) found a negative relationship between park area and abundance of House Sparrows in Valencia. However, the medium-sized parks favoured by birds in Valencia (Murgui Citation2009) were similar in size to the largest parks supporting the higher bird numbers in our study.
Greater spatial separation between urban parks reduced the abundance of birds, suggesting a positive role for connectedness between neighbouring parks. Again, Murgui (Citation2009) did not find such a relationship, probably due to urban differences between study areas, but clearly more research is required to understand how or if green connectivity promotes abundance of House Sparrows in urban areas. Rubbish bins may also provide a constant and reliable source of food for birds during the winter within the parks, so increasing their abundance. Contrary to expectations, rubbish bins were selected positively during the second winter, which was the warmer one. We expected that the species would be more dependent on these reliable food resources in the coldest seasons. However, a possible explanation for this situation could be related to the other selected variable in the second winter, the distance between parks. In this winter, closer parks had more House Sparrows than those far apart, so the movement of individuals between parks could have resulted in increased abundances of the species around key resources, such as rubbish bins.
The presence of dogs reduced the abundance of birds in summer. In this season, people walk the dog earlier, so increasing disturbance and perceived predation risk for House Sparrows (Forrest & St. Clair Citation2006), especially juvenile birds. Our results suggest that the species avoid parks with dogs, at least in summer. Peris & Montelongo (Citation2014) found no key factors for the presence or absence of House Sparrows in urban parks of Salamanca (Spain) during the 2009–10 breeding seasons. Besides, only the abundance and diversity of exotic trees (introduced from the native range of the species) was significant for the abundance of sparrows in a Mexican city during May 2004 (González-Oreja et al. Citation2012). More fine-scale research is urgently needed within their main feeding habitats to identify precisely the key factors that have led to the House Sparrow decline in urban areas.
Conclusions
We propose that the main causes of urban House Sparrow decline in southeast of Spain may be related to urban land use and infrastructure. Certainly, the replacement of above-ground rubbish bins by underground ones reduces the availability of food supplies to sparrows (Bokotey & Gorban Citation2005), but this highlights a major problem: urban areas and parks harbour scarce natural food for House Sparrows. In European cities generally, changes in urban parks such as increased paving, use of more pesticides, changes in habitat structure and use of sprinklers instead of hoses for the irrigation of lawns have decreased their suitability for the species and are responsible for its decline (Mitschke & Muslow Citation2003, Murgui Citation2009). Consequently, the Green Infrastructure Project of the Valencian Community (Cantó Citation2014) would be one urban planning option to halt and, in time, reverse the negative trend of the House Sparrow decline in the Valencian area because even small green patches provide suitable habitats for the birds (Rega et al. Citation2015). Similar projects could create sufficient heterogeneous urban green space that reduces predation of the target species in the long run (Summers-Smith Citation2003, Shaw et al. Citation2008) while also ameliorating air quality by reducing oxidative stress currently experienced by birds in polluted city areas (Herrera-Dueñas et al. Citation2017) and also promoting reproductive success of birds by increasing the availability of their natural foods (Peach et al. Citation2014). Also, the theory of ‘traffic evaporation’ (European Commission Citation2004), which proposes to reduce road capacity for cars in city centres, can represent a sustainable and efficient planning solution for European House Sparrows. Arguably, the development of high-rise buildings using modern materials and designs, increased socioeconomic status and increased urban population density are negatively impacting House Sparrow abundance (Shaw et al. Citation2008), so careful thought is required to accommodate all of these ‘forces’ to benefit the species through management options (European Commission Citation2004, Cantó Citation2014). Clearly, more fine-scale studies have an important part to play if we are to define management options that can be rolled out over large and disparate spatial scales in Spain and elsewhere.
Acknowledgements
We acknowledge Dan Chamberlain for a preliminary check of this manuscript. We kindly appreciate the constructive suggestions and comments made by James Reynolds which greatly improved previous versions of the text. We thank the editor, associate editor and two anonymous reviewers for their constructive comments, which helped us to improve the manuscript. We greatly appreciate the English review made by Debbie Berks.
References
- Anderson, T.R. 2006. Biology of the Ubiquitous House Sparrow: from genes to populations. Oxford University Press, Oxford.
- AVAMET. 2016. Meteoxarxa, Estadístiques. Associació Valenciana de Meteorologia ‘Josep Peinado’. Buñol, Spain. Available from: http://www.avamet.org/ [accessed October 2016].
- AVAMET. 2018. Meteoxarxa, Estadístiques. Temperatura mínima. Associació Valenciana de Meteorologia ‘Josep Peinado’. Buñol, Spain. Available from: https://www.avamet.org/mx-temperatura-minima.php [accessed April 2018].
- Balmori, A. & Hallberg, Ö. 2007. The urban decline of the house sparrow (Passer domesticus): a possible link with electromagnetic radiation. Electromagn. Biol. Med. 26: 141–151. doi: 10.1080/15368370701410558
- Banyuls, J. 1985. Evolución demográfica de la Safor en el siglo XX. Cuadernos de geografía. 37: 175–188.
- Bell, C.P., Baker, S.W., Parkes, N.G., Brooke, M. De L. & Chamberlain, D.E. 2010. The role of the Eurasian Sparrowhawk (Accipiter nisus) in the decline of the House Sparrow (Passer domesticus) in Britain. Auk. 127: 411–420. doi: 10.1525/auk.2009.09108
- Bernis, F. 1989. Los gorriones. Con especial referencia a su distribución y eto-ecología en las mesetas españolas. I.N.I.A., Ministerio de Agricultura, Pesca y Alimentación, Madrid.
- BirdLife International. 2016. Species factsheet: Passer domesticus. Available from: http://datazone.birdlife.org/species/factsheet/house-sparrow-passer-domesticus [accessed November 2016].
- Bokotey, A.A. & Gorban, I.M. 2005. Numbers, distribution and ecology of the House Sparrow in Lvov (Ukraine). Int. Stud. Sparrows. 30: 7–22.
- Cantó, M.T. 2014. La ordenación de la Infraestructura Verde en el sudeste Ibérico (Comunidad Valenciana, España). Cuadernos de Biodiversidad. 45: 10–22.
- Carrascal, L.M. & Palomino, D. 2008. Las aves comunes reproductoras en España. Población en 2004–2006. SEO/BirdLife, Madrid.
- Chamberlain, D.E., Vickery, J.A., Glue, D.E., Robinson, R.A., Conway, G.J., Woodburn, R.J.W. & Cannon, A.R. 2005. Annual and seasonal trends in the use of garden feeders by birds in winter. Ibis. 147: 563–575. doi:10.1111/j.1474-919x.2005.00430.x.
- Chamberlain, D.E., Toms, M.P., Cleary-McHarg, R. & Banks, A.N. 2007. House Sparrow (Passer domesticus) habitat use in urbanized landscapes. J. Ornithol. 148: 453–462. doi:10.1007/s10336-007-0165-x.
- Chamberlain, D.E., Glue, D.E. & Toms, M.P. 2009. Sparrowhawk Accipiter nisus presence and winter bird abundance. J. Ornithol. 150: 247–254. doi: 10.1007/s10336-008-0344-4
- Chevan, A. & Sutherland, M. 1991. Hierarchical partitioning. Am. Stat. 45: 90–96.
- Cramp, S. & Perrins, C.M. 1994. Handbook of the Birds of Europe, the Middle East and North Africa: the birds of the western Palearctic. Vol. 8. Crows to Finches. Oxford University Press, Oxford.
- Crick, H.Q.P., Robinson, R.A., Appleton, G.F., Clark, N.A. & Rickard, A.D. 2002. Investigation into the Causes of the Decline of Starlings and House Sparrows in Great Britain. BTO Research Report 290, British Trust for Ornithology, Thetford.
- De Laet, J. & Summers-Smith, J.D. 2007. The status of the urban house sparrow Passer domesticus in north-Western Europe: a review. J. Ornithol. 148: 275–278. doi: 10.1007/s10336-007-0154-0
- De Laet, J., Peach, W.J. & Summers-Smith, J.D. 2011. Protocol for censusing urban sparrows. Br. Birds. 104: 255–260.
- Ericson, P.G.P., Tyrberg, T., Kjellberg, A.S., Jonsson, L. & Ullén, I. 1997. The earliest record of house sparrows (Passer domesticus) in Northern Europe. J. Archaeol. Sci. 24: 183–190. doi: 10.1006/jasc.1996.0102
- European Commission. 2004. Reclaiming City Streets for People. Chaos or quality of life? EU Publications, Luxembourg.
- Field, A., Milles, J. & Field, Z. 2012. Discovering Statistics Using R. SAGE Publications, London.
- Forrest, A. & St. Clair, C.C. 2006. Effects of dog leash laws and habitat type on avian and small mammal communities in urban parks. Urban. Ecosyst. 9: 51–56. doi:10.1007%2Fs11252-006-7903-3 doi: 10.1007/s11252-006-7903-3
- Gibbons, D.W. & Gregory, R.D. 2006. Birds. In Sutherland, W.J. (ed.) Ecological Census Techniques: a handbook, 2nd edn. 324–328. Cambridge University Press, University of East Anglia, Norwich.
- Gil-Delgado, J.A. & Mora, J. 2015. Gorrión común (Passer domesticus). In: López, G.M., Bañuls, A., Zaragozí, A., Sala, J., Izquierdo, A., Martínez, J.E., Bañuls, D., Arroyo, S., Sánchez, J.A., Campos, B. & Reig, A. (eds.) Atlas de las aves nidificantes en la provincia de Alicante, 460–461. Publicacions de la Universitat d’Alacant – SEO/Alicante, Alicante.
- Gil-Delgado, J.A., Pardo, R., Bellot, J. & Lucas, I. 1979. Avifauna del naranjal valenciano II: el Gorrión Común (Passer domesticus L.). Mediterranea. 3: 69–99.
- Gil-Delgado, J.A., Vives-Ferrándiz, C. & Tapiero, A. 2002. Tendencia decreciente de una población de Gorrión Común Passer domesticus en los naranjales del este de España. Ardeola. 49: 195–210.
- González-Oreja, J.A., Barillas-Gómez, A.L., Bonache-Regidor, C., Buzo-Franco, D., García-Guzmán, J. & Hernández-Santín, L. 2012. Does habitat heterogeneity affect bird community structure in urban parks? In Lepczyk, C.A. & Warren P.S. (eds.) Urban Bird Ecology and Conservation, 1–16. Studies in avian biology, Vol. 45. University of California Press, Berkeley, CA.
- Hallstan, S., Grandin, U. & Goedkoop, W. 2010. Current and modeled potential distribution of the zebra mussel (Dreissena polymorpha) in Sweden. Biol. Invasions. 12: 285–296. doi: 10.1007/s10530-009-9449-9
- Hayhow, D.B., Conway, G., Eaton, M.A., Grice, P.V., Hall, C., Holt, C.A., Kuepfer, A., Noble, D.G., Oppel, S., Risely, K., Stringer, C., Stroud, D.A., Wilkinson, N. & Wotton, S. 2014. The State of the UK’s Birds 2014. RSPB, BTO, WWT, JNCC, NE, NIEA, NRW & SNH, Sandy, Bedfordshire.
- Herrera-Dueñas, A., Pineda J., Antonio, M.T. & Aguirre, J.I. 2015. The relationship between House Sparrow and the city: why urban populations are on decline? 10th Conference of the European Ornithologist’s Union. 24–28 August 2015. Badajoz.
- Herrera-Dueñas, A., Pineda-Pampliega, J., Antonio-García, M.T. & Aguirre, J.I. 2017. The influence of urban environments oxidative stress balance: a case study on the House Sparrow in the Iberian Peninsula. Front. Ecol. Evol. 5: 106. doi:10.3389/fevo.2017.00106.
- Indykiewicz, P. 1991. Nest and nest-sites of the House Sparrow Passer domesticus (Linnaeus, 1758) in urban, suburban and rural environments. Acta Zool. Cracov. 34: 475–495.
- Instituto Nacional de Estadística. 2016. INEbase. Demografía y población. Padrón. Población por municipios. INE, Madrid. http://www.ine.es/dynt3/inebase/index.htm?padre=525 [accessed November 2016].
- Izquierdo, A. & Sánchez, J.A. 2015. Gavilán común (Accipiter nisus). In López, G.M., Bañuls, A., Zaragozí, A., Sala, J., Izquierdo, A., Martínez, J.E., Bañuls, D., Arroyo, S., Sánchez, J.A., Campos, B. & Reig, A. (eds.) Atlas de las aves nidificantes en la provincia de Alicante, 151–153. Publicacions de la Universitat d’Alacant – SEO/Alicante, Alicante.
- Jongman, R.H.G., Ter Braak, C.J.F. & Van Tongeren, O.F.R. 1995. Data Analysis in Community and Landscape Ecology, 2nd edn. Cambridge University Press, Cambridge.
- Jordá, R.M. 1976. Alcoy: la crisis textil de 1965 y sus repercusiones. Cuadernos de geografía. 18: 27–36.
- Krauze-Gryz, D., Gryz, J.B., Goszczyński, J., Chylarecki, P. & Żmihorski, M. 2013. The good, the bad and the ugly: space use and intraguild interactions among three opportunistic predators – cat (Felis catus), dog (Canis familiaris) and fox (Vulpes vulpes) – under human pressure. Can. J. Zool. 90: 1402–1413. doi: 10.1139/cjz-2012-0072
- Liebl, A.L., Schrey, A.W., Andrew, S.C., Sheldon, E.L. & Griffith, S.C. 2015. Invasion genetics: lessons from a ubiquitous bird, the house sparrow Passer domesticus. Curr. Zool. 61: 465–476. doi: 10.1093/czoolo/61.3.465
- Lima, S. 1998. Non-lethal effects in the ecology of predator-prey interactions. BioScience. 48: 25–34. doi: 10.2307/1313225
- López-Iborra, G.M., Limiñana, R., Pavón, D. & Martínez-Pérez, J.E. 2011. Modelling the distribution of short-toed Eagle (Circaetus gallicus) in semi-arid Mediterranean landscapes: identifying important explanatory variables and their implications for its conservation. Eur. J. Wildl. Res. 57: 83–93. doi: 10.1007/s10344-010-0402-0
- Mac Nally, R. 1996. Hierarchical partitioning as an interpretative tool in multivariate inference. Aust. J. Ecol. 21: 224–228. doi:10.1111/j.1442-9993.1996.tb00602.x.
- Mac Nally, R. 2002. Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodivers. Conserv. 11: 1397–1401. doi: 10.1023/A:1016250716679
- Magudu, K. & Downs, C.T. 2015. The relative abundance of invasive House Sparrows (Passer domesticus) in an urban environment in South Africa is determined by land use. Afr. J. Wildl. Res. 45: 354–359. doi:10.3957/056.045.0354.
- McKinney, M.L. 2002. Urbanization, biodiversity and conservation. BioScience. 52: 883–890. doi: 10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2
- Meillère, A., Brischoux, F. & Angelier, F. 2015. Impact of chronic noise exposure on antipredator behavior: an experiment in breeding house sparrows. Behav. Ecol. 26: 569–577. doi:10.1093/beheco/aru232.
- Menon, M., Devi, P. & Mohanraj, R. 2013. Habitat variability and spatial assemblages of House Sparrows (Passer domesticus) along a gradient of urbanization. IOSR-JESTFT. 4: 1–11. doi: 10.9790/2402-0460111
- Mitschke, A. & Mulsow, R. 2003. Düstere Aussichten für einen haüfigen Stadtvögel-Vorkommen und Bestandsentwicklung des Haussperlings in Hamburg. Artenschutzreport. 14: 4–12.
- Monclús, F.J. 1998. La ciudad dispersa. Suburbanización y nuevas periferias. CCCB, Barcelona.
- Mouldrá, L., Zasadil, P., Mouldrý, V. & Šálek, M. 2018. What makes new housing development unsuitable for house sparrows (Passer domesticus)? Landsc. Urban Plan. 169: 124–130. doi:10.1016/j.landurbplan.2017.08.017.
- Murgui, E. 2007. Factors influencing the bird community of urban wooded streets along an annual cycle. Ornis. Fenn. 84: 66–77.
- Murgui, E. 2009. Seasonal patterns of habitat selection of the House Sparrow Passer domesticus in the urban landscape of Valencia (Spain). J. Ornithol. 150: 85–94. doi: 10.1007/s10336-008-0320-z
- Murgui, E. & Macias, A. 2010. Changes in the house sparrow Passer domesticus population in Valencia (Spain) from 1998 to 2008. Bird Study. 57: 281–288. doi: 10.1080/00063651003716762
- Peach, W.J., Mallord, J.W., Ockendon, N., Orsman, C.J. & Haines, W.G. 2015. Invertebrate prey availability limits reproductive success but not breeding population size in suburban House Sparrows Passer domesticus. Ibis. 157: 601–613. doi:10.1111/ibi.12264.
- Peach, W.J., Vincent, K.E., Fowler, J.A. & Grice, P.V. 2008. Reproductive success of house sparrows along an urban gradient. Anim. Conserv.. 11: 493–503. doi:10.1111/j.1469-1795.2008.00209.x.
- Peach, W.J., Sheehan, D.K. & Kirby, W.B. 2014. Supplementary feeding of mealworms enhances reproductive success in garden nesting House Sparrows Passer domesticus. Bird Study. 61: 378–385. doi:10.1080/00063657.2014.918577.
- Peris, S. & Montelongo, T. 2014. Birds and small urban parks: a study in a high plateau city. Turk. J. Zool. 38: 316–325. doi:10.3906/zoo-1305-20.
- QGIS. 2014. Quantum GIS Geographic Information System. Version 2.4 Chugiak. Open Source Geospatial Foundation Project. Available from: http://qgis.osgeo.org/en/site/ [accessed September 2014].
- Ralph, C.J., Geupel, G.R., Pyle, P., Martin, T.E., DeSante, D.F. & Milá, B. 1996. Métodos de censado. In Ralph, C.J., Geupel, G.R., Pyle, P., Martin, T.E., DeSante, D.F. & Milá, B. (eds.) Manual de métodos de campo para el monitoreo de aves terrestres, 32–38. Gen. Tech. Rep. PSW-GTR-159. Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture, Albany, CA.
- Raven, M.J., Noble, D.G. & Baillie, S.R. 2003. The Breeding Bird Survey 2002. BTO Research Report 334. British Trust for Ornithology, Thetford.
- Rega, C.C., Nilon, C.H. & Warren, P.S. 2015. Avian abundance patterns in relation to the distribution of small urban greenspaces. J. Urban Plan. D-ASCE. 141: A4015002. doi: 10.1061/(ASCE)UP.1943-5444.0000279
- Rega-Brodsky, C.C. & Nilon, C.H. 2016. Vacant lots as a habitat resource: nesting success and body condition of songbirds. Ecosphere. 7: e01578. doi:10.1002/ecs2.1578.
- Reynolds, S.J., Galbraith, J.A., Smith, J.A. & Jones, D.N. 2017. Garden bird feeding: insights and prospects from a north-south comparison of this global urban phenomenon. Front. Ecol. Evol. 5: 24. doi: 10.3389/fevo.2017.00024
- Robinson, R.A., Siriwardena, G.M. & Crick, H.Q.P. 2005. Size and trends of the House Sparrow Passer domesticus population in Great Britain. Ibis. 147: 552–562. doi:10.1111/j.1474-919x.2005.00427.x.
- Sætre, G–P., Riyahi, S., Aliabadian, M., Hermansen, J.S., Hogner, S., Olsson, U., Gonzalez, M.F., Sæther, S.A., Trier, C.N. & Elgvin, T.O. 2012. Single origin of human commensalism in the house sparrow. J. Evol. Biol. 25: 788–796. doi:10.1111/j.1420-9101.2012.02470.x.
- Savard, J-P.L. & Falls, J.B. 1981. Influence of habitat structure on the nesting height of birds in urban areas. Can. J. Zool. 59: 924–932. doi:10.1139/z81-132.
- Schroeder, J., Nakagawa, S., Cleasby, I.R. & Burke, T. 2012. Passerine birds breeding under chronic noise experience reduced fitness. Plos. One. 7(7): e39200. doi:10.1371/journal.pone.0039200.
- Senar, J.C. & Borras, A. 2004. Sobrevivir al invierno: estrategias de las aves invernantes en la Península Ibérica. Ardeola. 51: 133–168.
- Senar, J.C. & Copete, J.L. 1995. Mediterranean house sparrows (Passer domesticus) are not used to freezing temperatures: an analysis of survival rates. J. Appl. Statist. 22: 1069–1074. doi: 10.1080/02664769524829
- SEO/BirdLife. 2013. Resultados del programa SACRE de SEO/BirdLife. SEO/BirdLife, Madrid.
- Shaw, L.M. 2009. Investigating the Role of Socioeconomic Status in Determining Urban Habitat Quality for the House Sparrow, Passer domesticus. PhD Thesis, Exeter University.
- Shaw, L.M., Chamberlain, D. & Evans, M. 2008. The house sparrow Passer domesticus in urban areas: reviewing a possible link between post-decline distribution and human socioeconomic status. J. Ornithol. 149: 293–299. doi:10.1007/s10336-008-0285-y.
- Shaw, L.M., Chamberlain, D., Conway, G. & Toms, M. 2011. Spatial Distribution and Hábitat Preferences of the House Sparrow, Passer domesticus in Urbanised Landscapes. BTO Research Report 599, British Trust for Ornithology, Thetford.
- SigPac. 2014. Visor SigPac. Ministerio de Agricultura, Alimentación y Medio Ambiente. Gobierno de España, Madrid. Available from: http://sigpac.mapa.es/fega/visor [accessed September 2014].
- Siriwardena, G.M., Baillie, S.R., Buckland, S.T., Fewster, R.M., Marchant, J.H. & Wilson, J.D. 1998. Trends in the abundance of farmland birds: a quantitative comparison of smoothed common bird censuses. J. Appl. Ecol. 35: 24–23. doi: 10.1046/j.1365-2664.1998.00275.x
- Skórka, P., Sierpowska, K., Haidt, A., Myczko, L., Ekner-Grzyb, A., Rosin, Z.M., Kwiecinski, Z., Suchodolska, J., Takacs, V., Jankowiak, L., Wasierlewski, O., Graclik, A., Krawczyk, A.J., Kasprzak, A., Szwajkowski, P., Wylegala, P., Malecha, A.W., Mizera, T. & Tryjanowski, P. 2016. Habitat preferences of two sparrow species are modified by abundances of other birds in an urban environment. Curr. Zool. 62: 357–368. doi:10.1093/cz/zow069.
- Smith, R.M., Warren, P.H., Thompson, K. & Gaston K.J. 2006. Urban domestic gardens (VI): environmental correlates of invertebrate species richness. Biodivers. Conserv. 15: 2415–2438. doi:10.1007%2Fs10531-004-5014-0 doi: 10.1007/s10531-004-5014-0
- Summers-Smith, J.D. 1988. The Sparrows. T. & A.D. Poyser Ltd, Calton.
- Summers-Smith, J.D. 2000. Decline of House-Sparrows in large towns. Br. Birds. 93: 256–257.
- Summers-Smith, J.D. 2003. The decline of the House Sparrow: a review. Br. Birds. 96: 439–446.
- Summers-Smith, J.D. 2007. Is unleaded petrol a factor in urban house sparrow decline? Br. Birds. 100: 558–559.
- Tormo, J. 2011. La industria y la especialización productiva. In Tormo, J. (ed.) Mariola: sistema productivo y estrategia territorial, 95–130. Publicacions Universitat d’Alacant, Alicante.
- Vera, P. & Escandell, V. 2016. Ciència ciutadana al servei de la conservació de les aus comunes: el programa SACRE, de la Societat Espanyola d’Ornitologia, a la Comunitat Valenciana. Quaderns de Voluntariat i Participació Social, Vol. 2. Conselleria d’Agricultura, Medi Ambient, Canvi Climàtic i Desenvolupament Rural. Generalitat Valenciana, València.
- Vincent, K.E. 2005. Investigating the Causes of the Decline of the Urban House Sparrow Passer domesticus Population in Britain. PhD Thesis, DeMontfort University, Leicester.
- Walsh, C. & Mac Nally, R. 2015. Package ‘hier.part’. Available from: https://cran.r-project.org/web/packages/hier.part/hier.part.pdf [accessed November 2016].
- Weir, J.E.S. 2015. Urban Green Space Management for Invertebrates and House Sparrows. PhD Thesis, Imperial College London, London.
- Weitzel, N.H. 1988. Nest-site competition between the European starling and native breeding birds in northwestern Nevada. Condor. 90: 515–517. doi: 10.2307/1368590
- Woods, M., McDonald, R.A. & Harris, S. 2003. Predation by domestic cats Felis catus in Great Britain. Mamm. Rev. 33: 174–188. doi: 10.1046/j.1365-2907.2003.00017.x
- Wotton, S.R., Field, R., Langston, R.H.W. & Gibbons, D.W. 2002. Homes for birds: the use of houses for nesting by birds in the UK. Br. Birds. 95: 586–592.
- Zuur, A.F., Ieno, E.N. & Elphick, C.S. 2010. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 1: 3–14. doi: 10.1111/j.2041-210X.2009.00001.x