
ABSTRACT
Capsule: Atlantic Puffins Fratercula arctica from Scottish and Norwegian populations were significantly heavier in winter than when rearing chicks.
Aims: To compare body masses of Atlantic Puffins on their wintering grounds off the Faroe Islands with those of birds rearing chicks at colonies in Scotland and Norway.
Methods: We took standardized measurements of wing length and body mass of Atlantic Puffins during the summer chick-rearing period and on the wintering grounds near the Faroe Islands. These measurements were used to estimate seasonal changes in body mass for the two breeding populations. In three cases data were available for individuals weighed both at the colony and on the wintering grounds.
Results: On average, Atlantic Puffins breeding in Scotland and Norway increased their body mass by 20–30% between the chick-rearing period and winter. The very limited individual level data accorded well with the population level estimates.
Conclusions: Our results provide the first estimates of the order of magnitude in mass change between two key life history stages in this species. They indicate that gains in body mass between chick-rearing and winter are at least double the decline in mass previously recorded between incubation and chick-rearing. Given the Atlantic Puffin’s deteriorating conservation status, improved information on seasonal changes in body condition should help determine the underlying causes of die-offs in major wreck incidents such as those reported in recent years.
Many seabirds, including most species of tubenoses (Procellariiformes) and auks (Alcidae) and a few species of gulls (Laridae), spend the entire non-breeding season at sea, often thousands of kilometres away from their breeding colonies (Schreiber & Burger Citation2002). Over the last decades, the over-wintering behaviour of several species of seabirds breeding in the North Atlantic has been documented in detail through the use of various tracking devices, particularly light loggers (geolocators) (Frederiksen et al. Citation2012, Fayet et al. Citation2017, and results of SEATRACK at www.seapop.no/en/seatrack/). Results from these studies indicate that under normal conditions, many seabird species never come to land in autumn and winter.
Most pelagic seabirds are long-lived and invest more in survival than reproduction (Schreiber & Burger Citation2002). Consequently, populations are potentially vulnerable to poor conditions outside the breeding season, when most adult mortality is expected to occur. To increase their chances of surviving periods of poor food supply and, for the Atlantic species of auks, also reduced flight and diving abilities when they moult their main wing feathers more-or-less synchronously, seabirds are known to increase their body mass outside of the breeding season by storing fat in sub-cutaneous and visceral deposits (Camphuysen et al. Citation2007). Thus, the body mass of Common Guillemots Uria aalge taken as bycatch in fishing gear in the Skagerrak during late autumn and winter (950−1050 g), was much higher than that of breeding birds in nearby colonies in Scotland and South Norway (650−920 g; Cramp Citation1985, Anker-Nilssen & Lorentsen Citation1995, Barrett et al. Citation2008). However, the increase in body size with breeding latitude (Jones Citation1988, Barrett et al. Citation2008) and the uncertain origins of wintering birds because of their high dispersal outside the breeding season, make it difficult to assess how much weight individuals of any species add during the non-breeding season. Furthermore, such studies are also limited to the few species that are regularly reported as victims of hunting or bycatch in fishing gear. In both cases, sampling of the morphological size parameters needed to assess body condition and origins of the corpses requires close cooperation with the fishers or hunters, which is often difficult to establish.
Despite recent concerns about its population status, the Atlantic Puffin Fratercula arctica (hereafter Puffin) is probably still the most numerous seabird in the Northeast Atlantic (Mitchell et al. Citation2004, Barrett et al. Citation2006, IUCN Citation2017), but little is known about the body condition of adults in winter when they disperse over a huge part of the North Atlantic (Fayet et al. Citation2017). Puffins generally occur at low densities outside the breeding season but, although not highlighted by the survey of Fayet et al. (Citation2017), there is a notable concentration in the seas around the Faroe Islands in autumn and winter where it forms the basis of an annual legal hunt. Ringing recoveries indicate that many of these birds come from colonies in Scotland and Norway (). In this study, we use a 36-year long data set of body measurements of Puffins killed near the Faroes in winter to compare their body condition with birds of the same size and age in two breeding colonies, one in southeast Scotland and one in northern Norway from which many of these ringed birds originated. We predict that Puffins will be heavier in winter because they store fat to be able to buffer against larger-scale periods of reduced accessibility to food, whereas they can afford not to store fat in the chick period when events of food shortages are generally fewer and can be buffered against by abandoning the offspring at any stage. In addition, a leaner body condition will allow them to save energy when commuting between feeding and breeding grounds and act to increase breeding success (Nordberg Citation1981).
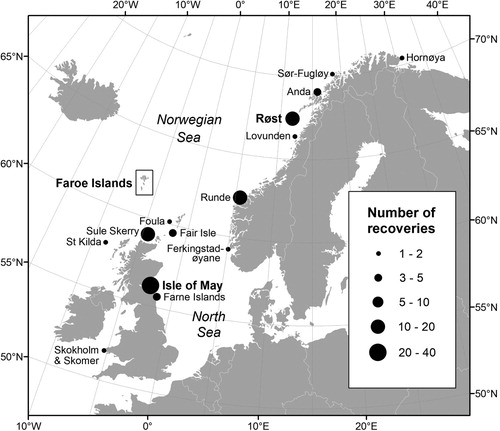
Figure 1. Ringing locations of 106 Atlantic puffins recovered around the Faroe Islands between October and March, as reported by December 2017. Totals for the Isle of May (38) and Røst (17) include a few birds ringed at nearby colonies. Names of study sites are indicated in bold.
Methods
Field work and morphological measurements
Puffins killed in Faroese waters, Northwest Europe between October and March in eight winters in 1981−2017 were examined by JKJ. Here, winters are referred to with their starting year, i.e. 1981 is winter 1981–82. Except for a few birds entangled in fishing nets and one individual hooked on a salmon drift line, all had been shot at sea by local hunters. Birds were collected on an opportunistic basis and aged by the number of bill grooves, sexed by gonadal inspection, weighed after being dried (but presumably still containing some lead shots) and had one wing (maximum flattened chord) measured (Camphuysen et al. Citation2007). For the birds classified as adults, i.e. those in their fourth calendar year or older, wing length and body mass were measured for 133 and 54 birds, respectively. The remainder had been plucked and/or eviscerated for human consumption or had incompletely grown primaries. Both measurements were taken for 47 individuals.
In 21 summers in 1973−2017 and 34 summers in 1980−2017, a total of 1469 and 2391 adults with chicks were captured on the Isle of May (56°11′N 02°33′W) in the Firth of Forth, Scotland and on Hernyken (67°26′N 11°52′E) in the Røst archipelago, Norway, respectively. The birds were either caught in a burrow with a chick (Isle of May) or in a mist net when carrying a food load (both sites). Thus, the age of the chick was most often unknown, but at both sites the sampling was usually spread throughout the chick period. In all cases, the adults were measured and/or weighed following the same protocol as the birds collected in winter. Both measurements were taken for 1449 and 1940 individuals, respectively. Some of these birds (186 from the Isle of May and 375 from Røst) were sexed by molecular DNA analysis or by using colony-specific discriminant functions based on the head + bill and wing lengths or bill measurements derived from adults sexed by DNA or gonadal inspection (Harris & Wanless Citation2011, Østvik Citation2017).
Data analysis
All data were analysed in IBM SPSS Statistics (ver. 24.0, IBM Corp. Citation2016). Differences in means of wing length and body mass were tested with one-way analysis of variance (ANOVA), whereas a simple 2 × 1 χ2 test was used as described by Zar (Citation1984) and corrected for continuity as described by Yates (Citation1934) to test if sex distributions deviated from equality. Differences in elevation and slope of the relationships between body mass and wing length were tested using the univariate general linear model (GLM) function with type III sum of squares, body mass as the dependent variable, wing length and year as covariates and site as a fixed factor, including also the interaction term between wing length and site. All test results with two-tailed P < 0.05 were considered significant.
Results
There was no evidence of a skewed sex distribution among adult birds in any of the samples (male–female ratio: Røst 183:192, χ2 = 0.171, df = 1, P = 0.678; Isle of May 87:99, χ2 = 0.651, df = 1, P = 0.420; Faroes 36:33, χ2 = 0.058, df = 1, P = 0.810). After pooling sexed and unsexed birds, body size of adults differed significantly between the three sites (one-way ANOVA, wing length: F2,3535 = 2604.7, P < 0.001, body mass: F2,3895 = 2463.8, P < 0.001; ). As expected, body mass was significantly and positively related to wing length among the wintering birds in the Faroes (Pearson r = 0.372, n = 47, P = 0.010) as well as among the chick-rearing adults at each of the two colonies (; Isle of May: r = 0.467, n = 1449, P < 0.001; Røst: r = 0.471, n = 1940, P < 0.001). When applying univariate GLM analyses using wing length and year as covariates to control for differences in body size and inter-annual differences in environmental conditions, we found significant differences in body mass of Puffins between the three sites (, ). This was mainly because the winter birds were much heavier than equal-sized breeders, although body mass varied significantly between years and the birds from the two colonies differed somewhat in mass. The significant interaction between wing and site was caused by the slope of the mass-wing relationship being less steep for the Isle of May birds than for birds at Røst or in the Faroes in winter, which did not differ significantly in slope.
Figure 2. Linear regressions (± 95% confidence intervals) of body mass as a function of wing length for adult Puffins killed in the Faroes (large white circles) in winter (October–March) and for chick-rearing adults at the colonies of Isle of May (southeast Scotland; black dots) and Røst (northern Norway; grey dots). Symbols indicate individual birds. For clarity, wing lengths of birds in the colonies are plotted 0.25 mm below (Isle of May) or above (Røst) the true values.
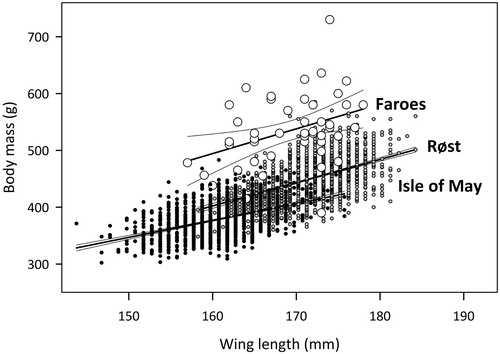
Table 1. Descriptive statistics of body mass and wing length for adult Puffins killed in Faroese waters in eight winters (October–March 1981–2018) and for chick-rearing adults at the colonies of Isle of May (Southeast Scotland) in 21 summers (1973–2017) and Røst (North Norway) in 34 summers (1980–2017). The sexed individuals are included in the ‘all’ categories.
Table 2. Univariate GLM tests of differences in body mass between adult Puffins killed in Faroese waters in winter (October–March) and chick-rearing adults at the colonies of Isle of May (southeast Scotland) and Røst (northern Norway), respectively, when controlling for differences in body size by entering wing length as a covariate.
The estimated marginal means of the pairwise comparisons indicate that adult birds from the Isle of May increased their body mass outside the breeding season by close to a third of their body mass in the chick period, whereas the equivalent increase for Røst was approximately a fifth of body mass ().
Table 3. Estimated differences in mean body mass between adult Puffins killed in Faroese waters in winter (October–March) and chick-rearing adults of the same size (as indicated by wing length) at the colonies of Isle of May (southeast Scotland) and Røst (northern Norway), respectively. The results are based on the estimated marginal means from the corresponding pairwise GLM analyses reported in .
Three Puffins were weighed both as adults in the colony and after being killed in Faroes waters in winter, one from each of the study colonies and one from Runde (62°24′N 05°36′E) in southwest Norway, situated approximately half-way between the Isle of May and Røst. The relative mass change of these three birds (mean 23.5%, range 10.9–32.5%; ) accorded well with our population level comparisons for the Isle of May and Røst and those killed in the Faroes. The bird from Røst was dissected and had 180 g of sub-cutaneous fat and 11 g of fat stored around its intestines, together amounting to 34% of the total winter body mass.
Table 4. Body mass (g) and wing length (mm) of three Atlantic puffins that were weighed as adults both at the colony in the chick period and when killed in Faroese waters in winter (October–March).
Two individuals from the Faroes were 20% heavier than the maximum published mass for this species (Lowther et al. Citation2002, Harris & Wanless Citation2011) and 70 g more than any other bird in our sample. One had a mass of 720 g, but no wing length was available since the primaries were incompletely grown. At its examination it was noted as being the size of a Razorbill Alca torda, a species that when feeding chicks is about 60% heavier than the Puffin (personal data), and its massive bill suggested it belonged to one of the high-arctic populations (the so-called Fratercula arctica naumanni). The other had a mass of 730 g and a wing of 174 mm and could thus also have come from a northern population.
Discussion
The non-breeding movements of Puffins breeding on the Isle of May and Røst have been followed using geolocator technology since 2007 and 2012, respectively (Harris et al. Citation2013, Fayet et al. Citation2017). Although birds from both colonies have been documented in waters around the Faroes, in neither instance were these areas the main wintering grounds, the former occurring mainly within the North Sea with only a minority entering the Atlantic, the latter remaining mostly around Iceland after most visiting the Barents Sea in autumn. Similar geolocator data from 14 other colonies throughout the species range showed that the Faroes was also used by birds from Skomer in Wales and from Runde (and by inference colonies in between where birds have not been tracked) but not by birds from three Norwegian colonies further north than Røst (data for 179 bird-winters), from Iceland or from colonies further west (Fayet et al. Citation2017, results of SEATRACK at www.seapop.no/en/seatrack/). However, earlier ringing recoveries () show that birds from the far north of Norway do sometimes occur in Faroese waters outside the breeding season.
The body size of Puffins in Europe increases with latitude of their breeding colony, with adults from Skomer and Runde having mean wing lengths of 159 and 169.5 mm, respectively (Harris & Wanless Citation2011). The differences in wing length, body mass and the slope of the relationship between these two size variables for Puffins breeding on the Isle of May and Røst, were therefore as expected and likely reflect similar trends for Puffins colonies north of Skomer and south of Røst where similar data for chick-feeding birds were not available.
Without more data for individual birds in both summer and winter, which are very difficult to obtain, we can only indirectly estimate the mass gain of Puffins between the two seasons. Consequently, the results of our analysis are likely to be biased by a number of factors that we cannot easily control for. First, the winter distribution of birds from any individual colony (Fayet et al. Citation2017) is unlikely to match the localized distribution of our sample from the Faroes, where environmental conditions in winter may well differ considerably from what most birds from the colony will experience. Second, the sample from the Faroes will inevitably include birds from other colonies as well as the Isle of May and Røst (). Third, we have not been able to control for the very variable conditions involved over such a long study period, which are also likely to have affected the hunting success in the Faroes, although such effects are dampened by including data from eight winters and 34 summers in our analysis. Fourth, the very variable timing of wing feather moult in Puffins (Harris et al. Citation2014, Anker-Nilssen et al. Citation2017) may also have acted to increase the variation in body condition of the birds collected, even if we excluded those that apparently were in active moult. Finally, the body mass measurements of adults in the colonies will be biased by the very different conditions for breeding documented in the two colonies over the study period (Harris et al. Citation2005, Harris & Wanless Citation2011). This may have induced differences in carry-over effects on movements outside the breeding season (Bogdanova et al. Citation2017; Fayet et al. Citation2017) and, thereby, also differences in body condition. Breeding success of Puffins on Røst was extremely variable from many years of total failures at the population level to 0.91 chicks fledged per egg hatched (Anker-Nilssen & Aarvak Citation2006), but less so on the Isle of May (0.30–0.93 fledged per pair; Harris & Wanless Citation2011). The seasons of nil success on Røst included seven of the eight summers immediately prior to the winters covered by the Faeroese data set as well as the four years when most ringed birds were recovered in the Faroes. Nevertheless, at neither colony was there a significant correlation between annual measures of breeding success and the masses of adults provisioning chicks in years when at least five weights were available (Røst: r² < 0.001, n = 30 years; Isle of May r² = 0.079, n = 27 years), which suggests that conditions on the breeding grounds were unlikely to influence body masses 3–6 months later.
Like most seabirds, Puffins are extremely mobile and characterized by high metabolism (Ellis & Gabrielsen Citation2002). This is, of course, especially true for the adults when they commute between often distant foraging grounds at sea and the colony in mid-summer to provision their chick (Harris et al. Citation2012). As activity levels and energy expenditure are likely to be much lower outside the breeding season, increasing body mass in winter has likely developed as a favourable adaptation allowing the birds to draw on their own fat stores to increase their chances of surviving unpredictable periods of poor food availability. Although increased body mass comes with an obvious cost in terms of reduced mobility, it will also act to reduce heat loss and, thereby, overall energy demand.
Our estimates could suggest a somewhat lower mass gain for Puffins from Røst (21%) than those from the Isle of May (31%), yet we cannot claim to have documented a true difference between the two study colonies. Our samples may also have been affected by an apparent difference in the timing of their presence in Faroese waters, as indicated by the significant difference in the average finding date of ringed Puffins, over the age of 5 years, originating from the UK compared to those from Norwegian colonies (UK: 28 October, n = 29, Norway: 9 December, n = 18; t = 3.851, df = 45, P < 0.001). Birds of different origins could also differ as to where and when they undertake the synchronous moult of the primary flight feathers (Harris et al. Citation2014, Anker-Nilssen et al. Citation2017), with further implications for their body mass.
Despite all these sources of bias, our results provide the only estimate of the average mass gain of Puffins outside the breeding season currently available. They indicate that on average, Puffins increase their body mass by 20–30% between the end of the breeding season and winter, i.e. at least twice the drop in mass recorded between incubation and chick feeding in other studies (Barrett et al. Citation1985, Harris & Wanless Citation2011). Although a more thorough analysis of such differences would require data at the individual level, we believe our results provide informative estimates of the order of magnitude in mass change between crucial life history stages in this species. Such knowledge is valuable for a number of purposes, for example, when assessing the underlying causes of Puffin die-offs in major wreck incidents such as those reported in recent years and which may have had significant impact at the population level (Anker-Nilssen et al. Citation2003, Citation2017, Harris & Elkins Citation2013, Morley et al. Citation2016). Our findings are also relevant to modelling of Puffin energetics outside the breeding season, for example, for assessing the impacts of prolonged periods of starvation on overwinter survival. This is critical for improving predictions of the effects of extreme weather events in the core areas of this species, which may increase in both frequency and intensity with global warming (Hanna et al. Citation2017 and references therein).
Acknowledgements
We thank Brett Sandercock at the Norwegian Institute for Nature Research (NINA) for statistical guidance, Geir Systad at NINA for making the map and all the people that assisted in the field work on the Isle of May National Nature Reserve and on Hernyken in Nykan Nature Reserve in Røst over the years – far too many to be mentioned by name. The Danish Bird Ringing Centre at the Natural History Museum of Denmark, University of Copenhagen provided the ringing and recovery data. Permits to work in the two nature reserves were granted by the County Governor of Nordland and the Scottish Natural Heritage, respectively. Sarah Wanless and two reviewers greatly improved the manuscript with their comments.
Additional information
Funding
References
- Anker-Nilssen, T. & Aarvak, T. 2006. Long-term studies of seabirds in the municipality of Røst, Nordland. Results with focus on 2004 and 2005. NINA Report 133. Trondheim: Norwegian Institute for Nature Research. (In Norwegian with English abstract and legends).
- Anker-Nilssen, T. & Lorentsen, S.-H. 1995. Size variation of Common Guillemots Uria aalge wintering in the northern Skagerrak. Seabird 17: 64−73.
- Anker-Nilssen, T., Aarvak, T. & Bangjord, G. 2003. Mass mortality of Atlantic Puffins Fratercula arctica off Central Norway, spring 2002: causes and consequences. Atlantic Seabirds 5: 57−71.
- Anker-Nilssen, T., Harris, M.P., Kleven, O. & Langset, M. 2017. Status, origin and population level impacts of Atlantic Puffins killed in a mass mortality event in SW Norway early 2016. Seabird 30: 1−14.
- Barrett, R.T., Anker-Nilssen, T., Bakken, V., Strøm, H., Krasnov, Y. & Aarvak, T. 2008. Biometrics as a determinant of the origins of seabirds killed in oil spills and other incidents. Bird Conserv. Int. 18: 229−241. doi: 10.1017/S0959270908000233
- Barrett, R.T., Chapdelaine, G., Anker-Nilssen, T., Mosbech, A., Montevecchi, W.A., Reid, J.R. & Veit, R.R. 2006. Seabird numbers and prey consumption in the North Atlantic. ICES J. Mar. Sci. 63: 1145−1158.
- Barrett, R.T., Fieler, R., Anker-Nilssen, T. & Rikardsen, F. 1985. Measurements and weight changes of Norwegian adult Puffins Fratercula arctica and Kittiwakes Rissa tridactyla during the breeding season. Ringing Migr. 6: 102−112. doi: 10.1080/03078698.1985.9673865
- BirdLife International 2017. Fratercula arctica. The IUCN Red List of Threatened Species 2017: e.T22694927A117606911.
- Bogdanova, M.I., Butler, A., Wanless, S., Moe, B., Anker-Nilssen, T., Frederiksen, M., Boulinier, T., Chivers, L.S., Christensen-Dalsgaard, S., Descamps, S., Harris, M.P., Newell, M., Olsen, B., Phillips, R.A., Shaw, D., Steen, H., Strøm, H., Thórarinsson, T.L. & Daunt, F. 2017. Multi-colony tracking reveals spatio-temporal variation in carry-over effects between breeding success and winter movements in a pelagic seabird. Mar. Ecol. Prog. Ser. 578: 167–181.
- Camphuysen, C.J., Bao, R., Nijkamp, H., Escuer, R.G. & Heubeck, M. (eds) 2007. Handbook on Oil Impact Assessment. http://www.zeevogelgroep.nl/downloads/handbook-oil-impact-assessment.
- Cramp, S. (ed) 1985. The Birds of the Western Palearctic, Vol. IV. Oxford University Press, Oxford.
- Ellis, H.I. & Gabrielsen, G.W. 2002. Energetics of free-ranging seabirds. In Schreiber, E.A. & Burger, J. (eds) Biology of Marine Birds, 360–407. CRC Press, Boca Raton, FL.
- Fayet, A.L., Freeman, R., Anker-Nilssen, T., Diamond, A., Erikstad, K.E., Fifield, D., Fitzsimmons, M., Hansen, E.S., Harris, M.P., Jessopp, M., Kouwenberg, A.-L., Kress, S., Mowat, S., Perrins, C.M., Petersen, A., Petersen, I.K., Þórarinsson, Þ.L., Reiertsen, T.K., Robertson, G., Sigurðsson, I.A., Shoji, A., Wanless, S. & Guilford, T. 2017. Ocean-wide drivers of migration strategies and their influence on population breeding performance in a declining seabird. Curr. Biol. 27: 1−8. doi: 10.1016/j.cub.2017.11.009
- Frederiksen, M., Moe, B., Daunt, F., Phillips, R.A., Barrett, R.T., Bogdanova, M.I., Boulinier, T., Chardine, J.W., Chastel, O., Chivers, L.S., Christensen-Dalsgaard, S., Clément-Chastel, C., Colhoun, K., Freeman, R., Gaston, A.J., González-Solís, J., Goutte, A., Grémillet, D., Guilford, T., Jensen, G.H., Krasnov, Y., Lorentsen, S.-H., Mallory, M.L., Newell, M., Olsen, B., Shaw, D., Steen, H., Strøm, H., Systad, G.H., Thórarinsson, T.L. & Anker-Nilssen, T. 2012. Multi-colony tracking reveals the winter distribution of a pelagic seabird on an ocean basin scale. Diversity Distrib. 18: 530−542. doi: 10.1111/j.1472-4642.2011.00864.x
- Hanna, E., Hall, R.J. & Overland, J.E. 2017. Can Arctic warming influence UK extreme weather? Weather 72: 346−352 doi: 10.1002/wea.2981
- Harris, M.P., Anker-Nilssen, T., McCleery, R.H., Erikstad, K.E., Shaw, D.N. & Grosbois, V. 2005. Effect of wintering area and climate on the survival of adult Atlantic puffins Fratercula arctica in the eastern Atlantic. Mar. Ecol. Prog. Ser. 297: 283−296. doi: 10.3354/meps297283
- Harris, M.P., Bogdanova, M.I., Daunt, F. & Wanless, S. 2012. Using GPS technology to assess feeding areas of Atlantic Puffins Fratercula arctica. Ringing Migr. 27: 43−49. doi: 10.1080/03078698.2012.691247
- Harris, M.P., Daunt, F., Bogdanova, M.I., Lahoz-Monfort, J.J., Newell, M.A., Phillips, R.A. & Wanless, S. 2013. Inter-year differences in survival of Atlantic puffins Fratercula arctica are not associated with winter distribution. Mar. Biol. 160: 2877−2889. doi: 10.1007/s00227-013-2278-5
- Harris, M.P. & Elkins, N. 2013. An unprecedented wreck of Puffins in eastern Scotland in March and April 2013. Scott. Birds 32: 157–159.
- Harris, M.P. & Wanless, S. 2011. The Puffin. T. & A.D. Poyser, London.
- Harris, M.P., Wanless, S. & Jensen, J.-K. 2014. When are Atlantic Puffins Fratercula arctica in the North Sea and around the Faroe Islands flightless? Bird Study 61: 182−192. doi: 10.1080/00063657.2014.909382
- IBM Corp. 2016. IBM SPSS Statistics for Windows, Version 24.0. IBM Corp, Armonk, NY.
- Jones, P.H. 1988. The European cline in wing-length of Guillemots Uria aalge. Seabird 11: 19−21.
- Lowther, P.E., Diamond, A.W., Kress, S.W., Robertson, G.J. & Russell, K. 2002. Atlantic Puffin (Fratercula arctica). In Poole, A. & Gill, F. (eds) The Birds of North America, Vol. 709: 1–24. The Birds of North America, Inc, Philadelphia, PA.
- Mitchell, I.P., Newton, S.F., Ratcliffe, N. & Dunn, T.E. 2004. Seabird Populations of Britain and Ireland. Results of the Seabird 2000 Census (1998−2002). T. & A.D. Poyser, London.
- Morley, T.I., Fayet, A.L., Jessop, H., Veron, P., Veron, M., Clark, J. & Wood, M.J. 2016. The seabird wreck in the Bay of Biscay and South-Western approaches in 2014: A review of reported mortality. Seabird 29: 22−28.
- Nordberg, R.A. 1981. Temporary weight decrease in breeding birds may result in more fledged young. Am. Nat. 118: 838−850. doi: 10.1086/283874
- Østvik, A.N. 2017. Effects of a fluctuating climate on the body condition of Atlantic Puffins Fratercula arctica at Røst, Norway. MSc Thesis, Norwegian University of Life Sciences, Ås.
- Schreiber, E.A. & Burger, J. 2002. Biology of Marine Birds. CRC Press, Boca Raton, FL.
- Yates, F. 1934. Contingency tables involving small numbers and the χ² test. J. R. Stat. Soc. Suppl. 1: 217–235. doi: 10.2307/2983604
- Zar, J. H. 1984. Biostatistical Analysis, 2nd edn. Prentice-Hall, Inc, Englewood Cliffs, NJ.