
ABSTRACT
Capsule: Pairs of White-throated Dippers Cinclus cinclus which defended winter territories bred earlier than non-territorial individuals, but there was no difference in reproductive success.
Aims: The effect of winter territoriality on breeding ecology has rarely been studied in resident birds. We carried out a preliminary investigation of whether winter territorial behaviour and territory size affect the timing of reproduction, breeding territory size and reproductive success in a riverine bird, the White-throated Dipper.
Methods: We monitored an individually marked population of White-throated Dippers in the UK. Wintering individuals were classified as either territorial or ‘floaters’ according to their patterns of occurrence and behaviour, and their nesting attempts were closely monitored in the subsequent months. Winter and breeding territory sizes were measured by gently ‘pushing’ birds along the river and recording the point at which they turned back.
Results: All birds defending winter territories did so in pairs, but some individuals changed partners before breeding. Territorial pairs that were together throughout the study laid eggs significantly earlier than pairs containing floaters and those comprising territorial birds that changed partners. However, there were no significant differences in clutch size, nestling mass or the number of chicks fledged. There was no relationship between winter territory length and lay date or any measure of reproductive success, although sample sizes were small. Winter territories were found to be significantly shorter than breeding territories.
Conclusion: Winter territoriality may be advantageous because breeding earlier increases the likelihood that pairs will raise a second brood, but further study is needed. Territories are shorter in winter as altitudinal migrants from upland streams increase population density on rivers, but this may also reflect seasonal changes in nutritional and energetic demands.
Territoriality, the defence of a distinct area from conspecifics, is a widespread and well-studied behaviour in birds. Territories are usually secured for reproductive purposes and provide individuals with exclusive access to limited resources, such as nesting sites or food (Brown Citation1964, Maher & Lott Citation2000). The quality of an individual’s territory is often a major determinant of mate choice (Alatalo et al. Citation1986, Eckerle & Thompson Citation2006) and reproductive success (van de Pol et al. Citation2006, Sergio et al. Citation2009b), and most field studies of territoriality have therefore focused on the breeding season. By contrast, relatively little is known about the fitness consequences of territoriality at other times of the year.
Territorial behaviour during winter has been documented in a variety of birds (Pulliam & Millikan Citation1982, Snow & Snow Citation1988, Cuadrado Citation1997). In some species, individuals establish a territory in order to monopolize a particular food source (Salomonson & Balda Citation1977, Snow & Snow Citation1988, Kelsey Citation1989). In others, it is thought that foraging strategies are sufficiently diverse to enable individuals to remain in a limited area and defend it economically (Brown Citation1964, Holmes et al. Citation1989, Salewski et al. Citation2002). Consequently, the overwinter survival of individuals which do not or cannot defend a territory may be reduced due to lower food availability or the costs associated with covering greater distances to feed (Jenkins et al. Citation1963, Holmes et al. Citation1989, Cuadrado Citation1995). In a study of wintering European Robins Erithacus rubecula, the feeding rates and body condition of territory holders were similar to those of non-territorial birds, but the former were able to feed in less exposed sites, suggesting that territories provided refuge from predators rather than access to food (Cuadrado Citation1997).
Winter territories are sometimes defended by birds in pairs or groups and this must offer fitness benefits to offset the costs of intraspecific competition for local resources. In Mute Swans Cygnus olor, established pairs may maintain winter territories to ensure access to high-quality breeding sites and prevent rivals from taking over the area (Scott Citation1984). In other species, pairs are formed during the autumn and defend a winter territory, either as a single pair or collectively in groups, before breeding together (Logan & Hyatt Citation1991, Hogstad Citation2014), and early pair bond formation may increase reproductive success (Fowler Citation1995). However, it has been shown in other species that individuals which defend a winter territory as a pair are unlikely to breed together and instead may benefit from reduced energetic expenditure associated with defence or increased rates of detecting rivals and predators (Davies & Houston Citation1981, Gwinner et al. Citation1994). Further work is needed to understand this variation in winter territorial strategies within and between species. In particular, very few studies have investigated the consequences of winter territoriality for the subsequent breeding season; the quality of winter territories in long-distance migrants is known to have so-called carry-over effects on the timing and success of reproduction (Norris et al. Citation2004, Paxton & Moore Citation2015), but little is known about similar effects in species which remain in the same area throughout the year.
White-throated Dippers Cinclus cinclus (hereafter ‘Dippers’) are aquatic songbirds found almost exclusively along rivers and streams where they feed on freshwater invertebrates and sometimes small fish (Tyler & Ormerod Citation1994). They provide an ideal system for studying winter territoriality because their territories are linear and therefore simple to survey and measure (D’Amico & Hemery Citation2003, Feck & Hall Citation2004, Chen & Wang Citation2010), but also because individuals vary in whether or not they defend a territory outside of the breeding season. Dippers are strongly territorial when breeding, but while some birds are thought to defend a territory throughout the winter, often in pairs, other individuals apparently become ‘floaters’ and are mobile, foraging in a territory until being evicted when encountered by the territory holder (Cramp Citation1988, Tyler & Ormerod Citation1994). Furthermore, birds breeding on upland streams often descend onto rivers in the winter where they compete for territories with local residents (Tyler & Ormerod Citation1994). However, the consequences of variation in winter territorial behaviour for reproductive success in the following season have never been studied.
Holding a territory in the winter may allow individuals to enter the breeding season in better condition, which may in turn lead to earlier or more successful reproduction. Dippers breeding earlier in the season typically have larger clutches, larger broods and are more likely to raise a second brood (Tyler & Ormerod Citation1985, Ormerod & Tyler Citation1993, Wilson Citation1996), although offspring survival may be lower in early broods and second broods (Ormerod & Tyler Citation1993). The size of the winter territory and whether or not it is defended by a pair may also impact on the timing and success of reproduction, especially if territorial pairs are future breeding partners. We test these hypotheses using field observations of an individually marked population of Dippers to: (1) quantify winter territorial behaviour and determine whether territorial pairs are future breeding partners; (2) compare the timing of breeding and reproductive success between winter territory holders and floaters; (3) investigate the relationship between winter territory size and breeding territory size and (4) investigate the relationship between winter territory size and the timing of breeding and reproductive success.
Methods
Study site and winter surveys
We monitored a population of Dippers in the River Lune catchment within four miles of Sedbergh, Cumbria, UK (54.3236°N 2.5282°W), between January and July 2016. This population of 40–50 breeding pairs has been individually marked and intensively studied since 2013. Each year, all unringed birds were caught in mist nets or in hand nets placed over the nest and fitted with a British Trust for Ornithology (BTO) metal ring together with a unique combination of three colour rings (under BTO licence). Individuals were sexed according to their wing length because there is minimal overlap between males and females (Demongin Citation2016); no discrepancies ever occurred when confirming sex from observations of breeding behaviour (e.g. incubation is carried out solely by females, Tyler & Ormerod Citation1994). Individuals were aged as either first-year or older (‘adults’) according to the presence or absence of unmoulted juvenile coverts, respectively (Demongin Citation2016). All nesting attempts were monitored through to fledging or failure (see below) and every nestling was metal and colour ringed (using the same system as for adults) when between one and two weeks old. Breeding began in March and the laying of first clutches typically took place between mid-March and mid-April each year.
The rivers and associated streams within the site were surveyed from 15 January to 12 February 2016 in order to identify territory holders and floaters. Access restrictions and flooding due to a severe storm in December 2015 meant that only approximately 60% of the site could be reliably surveyed; birds present on the remaining waterways were excluded from the study. Each stretch of river was surveyed once per week and three or four times in total. Surveys involved walking slowly alongside each section of water, identifying all colour ringed birds using binoculars and recording the location where each was first encountered using a Garmin eTrex H global positioning system (GPS) unit (Garmin Ltd., Switzerland). Birds were classified as territorial if they were recorded within 500 m of the same location on each survey (typical winter territory sizes in this study population are 500–1000 m in length, SPS unpubl. data; see Results). All of these birds showed behaviour typical of territoriality, including singing, chasing of intruders and consistent ‘double backing’ when pushed in a particular direction (see below). Birds were considered to be in territorial pairs if the same two individuals were recorded together (i.e. within 1–50 m of each other) each time; members of a pair were occasionally found further apart but always reunited when disturbed by the observer or to chase away another bird which had entered their territory. Individuals were classified as floaters if recorded at different locations more than 1 km apart on at least three surveys; these birds were never seen in a pair and did not show any territorial behaviour. All other individuals recorded during the winter survey period were only seen once or twice, in the same or different locations, and were not classified. This included six colour ringed individuals and what we estimate to be a similar number of unmarked birds.
Territory size
To measure territory size in the winter and in the breeding season, upstream and downstream territory limits were identified using similar methods to researchers working with other Cinclus species (Feck & Hall Citation2004, Chen & Wang Citation2010). Birds were gently pushed upstream or downstream by walking behind them until they doubled back in the opposite direction; the turning point was recorded with a GPS. The focal bird was whichever member of a pair the observer first encountered, and visits were abandoned if the focal bird interacted with a conspecific other than its partner (who was rarely more than 15 m away). Two upstream and two downstream limits were identified for each territory, all on separate visits; all pairs of upstream or downstream limits were within 20 m of each other, so no further visits were considered necessary. No territories contained forks in the river or major tributaries. Small tributaries were rarely used by the birds in our study area other than in times of severe flooding, and these streams were therefore excluded from our measurements because it was not possible to push birds onto them. The furthest upstream and downstream limits for each territory were then inputted into ArcGIS version 10.2 (Esri, USA) in order to calculate the length of the territory, measured along the watercourse rather than as the Euclidean distance between the two limits. Winter territories were measured from 15 February to 6 March and, for any given pair, at least four weeks before laying began (median first egg lay date for winter territorial pairs = 2 April, range = 20 March to 12 April); breeding territories were measured between 4 and 29 April after the pair had begun building a nest. It is possible that territory limits may have shifted between the time when winter territories were first identified and when they were measured, however, our observations suggest that the limits remain unchanged until floaters start to pair up and this was not recorded until at least a month after measurements were completed. The breeding territory size of some pairs could not be measured due to access restrictions.
Reproductive success
All nests were found by following birds carrying nest material or by regularly inspecting traditional sites, and then closely monitored until fledging or failure. Once a pair had been observed lining their nest with leaves, the nest was checked every two to three days for eggs in order to determine lay date (i.e. the date on which the first egg of the clutch was laid). Dippers lay one egg per day and do not start incubating until the penultimate or final egg has been laid (Tyler & Ormerod Citation1994); if fresh eggs are cold it is therefore possible to determine lay date by back calculation. Clutch size was measured approximately one week after incubation had begun by counting the total number of eggs present. After a typical incubation period of 16 days (Tyler & Ormerod Citation1994), nests were checked daily for hatching; the date of hatching was occasionally missed by up to two days but the nestlings could then be aged from their appearance. All nestlings were ringed and weighed using an electronic balance when they were nine days old. Nests were then checked at least once per week until fledging was due, at which point daily checks were made to confirm fledging and to inspect the nest for dead nestlings. For all successful nests, the number of chicks fledged was taken to be the brood size at ringing as no cases of mortality after this point were recorded.
Statistical analysis
The following variables were measured for each nest: lay date, converted into a Julian date as the number of days since the end of February (i.e. 1st March = 1); clutch size; mean nestling mass (the mean body mass of all chicks in the brood when nine days old); and the number of chicks fledged (zero for failed nests). All of these variables and territory sizes were non-normally distributed, so non-parametric tests were used throughout. All analyses were carried out in SPSS version 20 (IBM Corp., USA).
Results
All birds classified as territorial in winter defended their territories in heterosexual pairs (n = 14 pairs); six pairs contained at least one first year bird but all eight pairs comprising two adults had bred together in the previous season (S.P. Sharp, unpubl. data). Most pairs (9/14 = 64.3%) also bred together in the following season. In three of the other pairs, one member was not seen at all after winter and presumed dead, while the remaining member paired up with a bird which had not been recorded at all during winter surveys. The final two pairs switched partners following the breakdown of a bigamous relationship: one female abandoned her part-built nest and the male with which she had defended a winter territory, instead breeding with a bird which had not been classified and leaving her original male to continue breeding with his other female. All floaters (n = 14) bred with other floaters or with birds which had either not been classified or not been seen at all during winter (n = 10 pairs). There was no clear evidence of any lone birds defending a winter territory, although some of the unclassified birds were occasionally seen chasing other unidentified individuals.
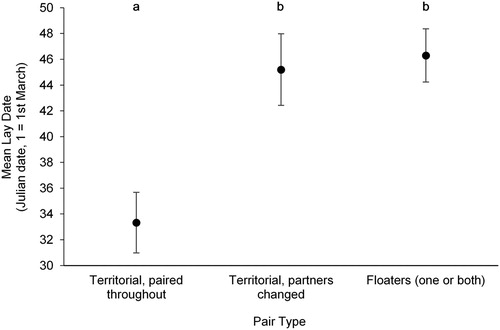
The lay dates of pairs which were territorial during winter and remained together (n = 9) were significantly earlier than those of birds which were territorial but had changed partners since winter (n = 5) and those of pairs containing floaters (n = 10; Kruskal–Wallis test: χ2 = 11.719, df = 2, P = 0.003; ). The earliest lay date for a winter territorial pair was day 20 (20th March); the earliest lay dates of pairs with new partners or pairs containing floaters were day 41 (10th April) and day 35 (4th April), respectively. By contrast, pairs from the three groups did not differ significantly in their clutch size (χ2 = 0.943, df = 2, P = 0.624), mean nestling mass (χ2 = 0.788, df = 2, P = 0.674) or number of chicks fledged (χ2 = 1.372, df = 2, P = 0.504).
Figure 1. The mean lay dates (± se) of breeding pairs of Dippers with different territorial behaviour in the previous winter. Territorial pairs are divided into those which remained together in the breeding season and those containing birds which changed partners between winter and the breeding season. The remaining pairs are those which contained at least one floater. Letters (a and b) denote significant differences (P < 0.05) determined using Dunn–Bonferroni post hoc comparisons following a Kruskal–Wallis test.
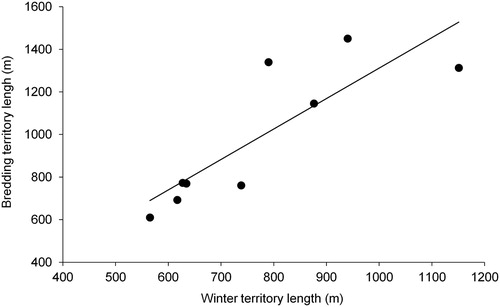
For the pairs which remained together throughout the study period, winter territories were significantly shorter than breeding territories (Wilcoxon signed rank test: Z = 2.666, n = 9, P = 0.008; ) but there was a significant positive correlation between the length of winter and breeding territories (Spearman correlation: rs = 0.850, n = 9, P = 0.004; ). There was no significant correlation between winter territory length and lay date (rs = –0.167, n = 9, P = 0.667), clutch size (rs = 0.608, n = 9, P = 0.083), mean nestling mass (rs = 0.095, n = 8, P = 0.823) or number of chicks fledged (rs = –0.059, n = 9, P = 0.879).
Figure 2. The mean winter and breeding territory lengths (± se) of pairs of Dippers which remained together throughout the study period.
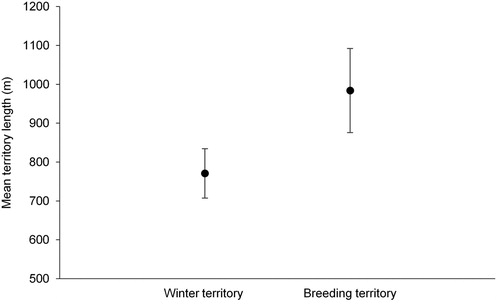
Figure 3. The relationship between winter territory length and breeding territory length in pairs of Dippers which remained together throughout the study period.
Discussion
Winter territorial behaviour varied between individuals in our study site. This may help to explain the conflicting nature of previously published observations of wintering Dippers; these include reports of highly territorial birds in some regions but an apparent absence of winter territories in others, though in many cases the birds were not individually marked (reviewed in Tyler & Ormerod Citation1994). All territorial birds remained in heterosexual pairs throughout the winter, many of them established pairs. While it is possible that some of the unclassified birds or those wintering in areas that were not surveyed may have been territorial and single, such birds are unlikely to be common as more than half of the wintering population was classified and solitary birds seen once were, by definition, absent from that location on other visits.
Pairs which were territorial in winter and remained together to breed began laying significantly earlier than those comprising birds which had changed partners or pairs containing floaters. This may be selectively advantageous because breeding earlier increases the likelihood that pairs will be able to raise a second brood (Tyler & Ormerod Citation1985, Ormerod & Tyler Citation1993, Wilson Citation1996). Double brooding can substantially increase annual and lifetime reproductive success in birds (Hoffmann et al. Citation2015, Cornell & Williams Citation2016), although in Dippers these benefits may be offset by lower offspring survival in early and second broods (Ormerod & Tyler Citation1993). The 2016 breeding season was later than previous seasons (S.P. Sharp, unpubl. data) and only four of the pairs in this study attempted to raise a second brood. It is worth noting that all of these were winter territorial pairs, but further study over multiple seasons is needed to investigate the long-term fitness consequences of winter territoriality.
So-called carry-over effects have been widely reported in long-distance migrants, with higher quality winter territories enabling individuals to maintain better body condition, arrive on their breeding grounds earlier and have higher reproductive success (Norris et al. Citation2004, Paxton & Moore Citation2015). Similar effects may be operating in Dippers with birds occupying winter territories securing access to good feeding sites, bringing them into breeding condition earlier in the year. If this is the case, then it is perhaps surprising that other reproductive measures did not differ between the groups, although Wilson (Citation1996) also found that earlier breeders raised more second broods yet did not have larger clutches or broods. It may be that the lateness of the 2016 season masked any effects, or that winter territoriality yields fitness benefits not measured here, for example offspring or adult survival. Alternatively, it may simply be that because winter territorial birds had paired up sooner, they began breeding earlier (Zack & Stutchbury Citation1992). Many had bred together in previous years, and a longer pair bond can also lead to earlier reproduction (Fowler Citation1995).
Floating may be an alternative winter strategy with fitness benefits we were unable to detect offsetting any costs associated with later reproduction. However, given the greater distances covered by floaters and the high levels of aggression they experience, it seems more likely that these are younger or poorer quality individuals which are unable to compete for winter territories; this is often true of floaters (Smith & Arcese Citation1989, Sergio et al. Citation2009a) including in other winter territorial species (Stutchbury Citation1994, Hogstad Citation1999). Floating may therefore be ‘making the best of a bad situation’, for example by monitoring multiple territories in order to identify vacancies when they arise (Smith Citation1984, Hogstad Citation1999). The exact age and body condition of many birds in our study was unknown, but ongoing work will shed more light on the age and quality of floaters relative to territorial birds as well as investigating whether individuals adopt the same strategy in successive winters.
Birds that changed partners between winter and the breeding season also laid their clutches later than pairs which remained together throughout. Again, this is likely due to the time spent finding a new partner and the shorter period of pair bonding (Fowler Citation1995), and there is experimental evidence in other passerines that remaining with the same partner allows pairs to breed sooner (Adkins-Regan & Tomaszycki Citation2007). Most of the birds in our study changed partner after the disappearance and presumed death of their previous partner. However, it is also possible that some birds ‘divorce’ to acquire a more compatible or higher quality mate (Choudhury Citation1995).
The smaller size of winter territories than breeding territories held by the same pair is likely a consequence of increases in population density during winter. Birds breeding on upland streams where winter conditions are harsh often move lower to the adjoining rivers and remain there until spring, competing with local territory holders (Tyler & Ormerod Citation1994). Similar findings have been reported in American Dippers Cinclus mexicanus, in which altitudinal migration increases winter population densities and reduces the size of available territories (Morrissey et al. Citation2004). However, winter territories may also be shorter than breeding territories because birds spend less time performing energetically expensive behaviour such as flying or territory defence (Bryant et al. Citation1985, Ormerod & Tyler Citation1991), and make greater use of fish in their diet (Ormerod & Tyler Citation1991); this may be important at a time when environmental conditions are typically harsher. The finding that pairs with larger winter territories also have larger breeding territories may reflect the quality of these birds and their ability to defend a longer stretch of river while still foraging optimally. Breeding territory quality is often related to individual quality, although it is notoriously challenging to determine the direction of any causal relationship (Germain & Arcese Citation2014).
Sample sizes were small in our study, especially when testing the relationship between territory length and breeding success. Furthermore, it is difficult to assess how the lateness of the 2016 breeding season or the exceptional severity of the storm which preceded the study may have impacted the results. However, it is possible that territory length is a poor measure of territory quality in Dippers. Wilson (Citation1996) found that in areas of deep, slow moving water where food is scarce, Dippers defend longer territories than in wider areas of fast-flowing and well-oxygenated water that provide preferred feeding sites such as riffles. The correlation between winter and breeding territory sizes may simply reflect the number of and distance between riffles in that area; territory length may not predict lay date or breeding success if unrelated to quality. Further work on winter territoriality in this species should therefore include alternative measures of territory quality such as food availability.
Acknowledgements
We would like to thank the many local landowners and Paul Wilkinson at Yorkshire Dales National Park for providing access and supporting our work; Tess Moore, Wesley Payne and Rebecca Simpson for their invaluable help with fieldwork; and Duncan Whyatt for assistance with ArcGIS.
ORCID
Lucy Magoolagan http://orcid.org/0000-0002-1961-1114
Stuart P. Sharp http://orcid.org/0000-0002-3059-2532
Additional information
Funding
References
- Adkins-Regan, E. & Tomaszycki, M. 2007. Monogamy on the fast-track. Biol. Lett. 3: 617–619. doi: 10.1098/rsbl.2007.0388
- Alatalo, R.V., Lundberg, A. & Glynn, C. 1986. Female pied flycatchers choose territory quality and not male characteristics. Nature 323: 152–153. doi: 10.1038/323152a0
- Brown, J.L. 1964. The evolution of diversity in avian territorial systems. Wilson Bull. 76: 160–169.
- Bryant, D.M., Hails, C.J. & Prys-Jones, R. 1985. Energy expenditure by free-living dippers (Cinclus cinclus) in winter. Condor 87: 177–186. doi: 10.2307/1366880
- Chen, C.C. & Wang, Y. 2010. Relationships between stream habitat and breeding territory length of the brown dipper (Cinclus pallasii) in Taiwan. J. Ornithol. 151: 87–93. doi: 10.1007/s10336-009-0429-8
- Choudhury, S. 1995. Divorce in birds: a review of the hypotheses. Anim. Behav. 50: 413–429. doi: 10.1006/anbe.1995.0256
- Cornell, A. & Williams, T.D. 2016. Individual quality and double-brooding in a highly synchronous songbird population. Auk 133: 251–260. doi: 10.1642/AUK-15-165.1
- Cramp, S. (ed) 1988. The Birds of the Western Palearctic, Vol. 5. Oxford University Press, Oxford.
- Cuadrado, M. 1995. Winter territoriality in migrant Black Redstarts Phoenicurus ochrurus in the Mediterranean area. Bird Study 42: 232–239. doi: 10.1080/00063659509477172
- Cuadrado, M. 1997. Why are migrant robins (Erithacus rubecula) territorial in winter?: the importance of the anti-predatory behaviour. Ethol. Ecol. Evol. 9: 77–88. doi: 10.1080/08927014.1997.9522904
- D’Amico, F. & Hemery, G. 2003. Calculating census efficiency for river birds: a case study with the white-throated dipper Cinclus cinclus in the Pyrénées. Ibis 145: 83–86. doi: 10.1046/j.1474-919X.2003.00108.x
- Davies, N.B., & Houston, A.I. 1981. Owners and satellites: the economics of territory defence in the pied wagtail, Motacilla alba. J. Anim. Ecol. 50: 157–180. doi: 10.2307/4038
- Demongin, L. 2016. Identification Guide to Birds in the Hand. Beauregard-Vendon, Privately published.
- Eckerle, K.P. & Thompson, C.F. 2006. Mate choice in house wrens: nest cavities trump male characteristics. Behaviour 143: 253–271. doi: 10.1163/156853906775900694
- Feck, J. & Hall, R.O. 2004. Response of American dippers (Cinclus mexicanus) to variation in stream water quality. Freshwater Biol. 49: 1123–1137. doi: 10.1111/j.1365-2427.2004.01254.x
- Fowler, G.S. 1995. Stages of age-related reproductive success in birds: simultaneous effects of age, pair-bond duration and reproductive experience. Amer. Zool. 35: 318–328. doi: 10.1093/icb/35.4.318
- Germain, R.R. & Arcese, P. 2014. Distinguishing individual quality from habitat preference and quality in a territorial passerine. Ecology 95: 436–445. doi: 10.1890/13-0467.1
- Gwinner, E., Rödl, T. & Schwabl, H. 1994. Pair territoriality of wintering stonechats: behaviour, function and hormones. Behav. Ecol. Sociobiol. 34: 321–327. doi: 10.1007/BF00197002
- Hoffmann, J., Postma, E. & Schaub, M. 2015. Factors influencing double brooding in Eurasian hoopoes Upopa epops. Ibis 157: 17–30. doi: 10.1111/ibi.12188
- Hogstad, O. 1999. Territory acquisition during winter by juvenile willow tits Parus montanus. Ibis. 141: 615–620. doi: 10.1111/j.1474-919X.1999.tb07369.x
- Hogstad, O. 2014. Pair-bond formation and mate fidelity among juvenile willow tits. Ornis Norv. 37: 7–10. doi: 10.15845/on.v37i0.607
- Holmes, R.T., Sherry, T.W. & Reitsma, L. 1989. Population structure, territoriality and overwinter survival of two migrant warbler species in Jamaica. Condor 91: 545–561. doi: 10.2307/1368105
- Kelsey, M.G. 1989. A comparison of the song and territorial behaviour of a long-distance migrant, the marsh warbler Acrocephalus palustris, in summer and winter. Ibis 131: 403–414. doi: 10.1111/j.1474-919X.1989.tb02788.x
- Jenkins, D., Watson, A. & Miller, G.R. 1963. Population studies on red grouse, Lagopus lagopus scoticus (Lath.) in north-east Scotland. J. Anim. Ecol. 32: 317–376. doi: 10.2307/2598
- Logan, C.A. & Hyatt, L.E. 1991. Mate attraction by autumnal song in the Northern mockingbird (Mimus polyglottos). Auk 108: 429–432.
- Maher, C.R. & Lott, D.F. 2000. A review of ecological determinants of territoriality within vertebrate species. Am. Midl. Nat. 143: 1–29. doi: 10.1674/0003-0031(2000)143[0001:AROEDO]2.0.CO;2
- Morrissey, C.A., Bendell-Young, L.I. & Elliott, J.E. 2004. Seasonal trends in population density, distribution, and movement of American dippers within a watershed of southwestern British Columbia, Canada. Condor 106: 815–825. doi: 10.1650/7455
- Norris, D.R., Marra, P.P., Kyser, T.K., Sherry, T.W. & Ratcliffe, L.M. 2004. Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. Proc. R. Soc. Lond. B 271: 59–64. doi: 10.1098/rspb.2003.2569
- Ormerod, S.J. & Tyler, S.J. 1991. Exploitation of prey by a river bird, the dipper Cinclus cinclus (L.), along acidic and circumneutral streams in upland Wales. Freshwater Biol. 25:105–116. doi: 10.1111/j.1365-2427.1991.tb00477.x
- Ormerod, S.J. & Tyler, S.J. 1993. The adaptive significance of brood size and time of breeding in the dipper Cinclus cinclus (Aves: Passeriformes) as seen from post-fledging survival. J. Zool. 231: 371–381. doi: 10.1111/j.1469-7998.1993.tb01925.x
- Paxton, K.L. & Moore, F.R. 2015. Carry-over effects of winter habitat quality on en route timing and condition of a migratory passerine during spring migration. J. Avian Biol. 46: 495–506. doi: 10.1111/jav.00614
- Pulliam, H.R. & Millikan, C. 1982. Social organization in the nonreproductive season. In Farner, D.S., King, J.R. & Parkers, K.C. (eds) Avian Biology, 169–197. Academic Press, New York.
- Salewski, V., Bairlein, F. & Leisler, B. 2002. Different wintering strategies of two Palearctic migrants in West Africa – a consequence of foraging strategies? Ibis 144: 85–93. doi: 10.1046/j.0019-1019.2001.00007.x
- Salomonson, M.G. & Balda, R.P. 1977. Winter territoriality of Townsend's solitaires (Myadestes townsendi) in a pinon-juniper-ponderosa pine ecotone. Condor 79: 148–161. doi: 10.2307/1367157
- Scott, D.K. 1984. Winter territoriality of mute swans Cygnus olor. Ibis 126: 168–176. doi: 10.1111/j.1474-919X.1984.tb07996.x
- Sergio, F., Blas, J. & Hiraldo, F. 2009a. Predictors of floater status in a long-lived bird: a cross-sectional and longitudinal test of hypotheses. J. Anim. Ecol. 78: 109–118. doi: 10.1111/j.1365-2656.2008.01484.x
- Sergio, F., Blas, J., Baos, R., Forero, M.G., Donázar, J.A. & Hiraldo, F. 2009b. Short- and long-term consequences of individual and territory quality in a long-lived bird. Oecologia 160: 507–514. doi: 10.1007/s00442-009-1314-0
- Smith, S.M. 1984. Flock switching in chickadees: why be a winter floater? Amer. Nat. 123: 81–98. doi: 10.1086/284188
- Smith, J.N. & Arcese, P. 1989. How fit are floaters? Consequences of alternative territorial behaviours in a non-migratory sparrow. Amer. Nat. 166: 830–845.
- Snow, B. & Snow, D. 1988. Birds and Berries. T & AD Poyser, London.
- Stutchbury, B.J. 1994. Competition for winter territories in a neotropical migrant: the role of age, sex and color. Auk 111: 63–69. doi: 10.2307/4088505
- Tyler, S.J. & Ormerod, S.J. 1985. Aspects of the breeding biology of Dippers Cinclus cinclus in the southern catchment of the River Wye, Wales. Bird Study 32: 164–169. doi: 10.1080/00063658509476874
- Tyler, S.J. & Ormerod, S.J. 1994. The Dippers. T & AD Poyser, London.
- van de Pol, M., Bakker, T., Saaltink, D-J. & Verhulst, S. 2006. Rearing conditions determine offspring survival independent of egg quality: a cross-foster experiment with oystercatchers Haematopus ostralegus. Ibis 148: 203–210. doi: 10.1111/j.1474-919X.2006.00479.x
- Wilson, J.D. 1996. The breeding biology and population history of the Dipper Cinclus cinclus on a Scottish river system. Bird Study 43: 108–118. doi: 10.1080/00063659609461001
- Zack, S. & Stutchbury, B.J. 1992. Delayed breeding in avian social systems: the role of territory quality and “floater” tactics. Behaviour 123: 194–219. doi: 10.1163/156853992X00020