
ABSTRACT
Capsule: Fine-scale use of climate-sensitive habitats by White-winged Snowfinches Montifringilla nivalis is affected by landscape composition, which thus modulates the potential impact of climate change.
Aims: To explore the fine-scale habitat use for foraging by breeding White-winged Snowfinches, with the aim of identifying the potential impacts of climate change on their dependence on habitat characteristics in the wider landscape, which could shape the species’ response to climate change. Pairs mostly relying on snow could be at higher risk due to a decrease in spring snow-cover, but the reliance on snow may be mediated by the presence of alternative habitats in the landscape.
Methods: The White-winged Snowfinch uses alpine grassland, snowfields and melting snow margins for capturing prey during nestling rearing and is threatened by earlier snowmelt induced by climate change. We investigated habitat use in 17 Snowfinch pairs in relation to landscape composition within a 300 m buffer around their nests.
Results: The use of snowfields and snow-bare margins was positively associated with wind intensity (probably due to greater wind-borne arthropod fallout with wind), and negatively associated with grassland cover (i.e. the main alternative habitat) and date within the season.
Conclusions: Fine-scale habitat selection was affected by landscape composition: the availability of patches of alternative habitats decreased the reliance on more climate-sensitive resources, modulating the potential impact of climate change. Coupling assessments of fine-scale resource selection with broader habitat descriptors, which are easier to assess over broader scales, may help understand and predict climate change impacts.
Climatic changes are impacting species of every taxonomic group across all continents, but their impacts are largely modulated by habitat characteristics (Otto et al. Citation2014, Bowden et al. Citation2015, Selwood et al. Citation2015). Local habitat may actually buffer the effects of climate change (Scherrer & Körner Citation2011, Nieto-Sánchez et al. Citation2015), although habitat management is unlikely to compensate for losses due to climatic alterations (Wessely et al. Citation2017; but see Braunisch et al. Citation2014, Brambilla et al. Citation2018). Microhabitat selection and, more generally, fine-scale habitat use, plays a fundamental role in a species’ capacity to buffer changes due to climate variation (Higgins et al. Citation2010, Suggitt et al. Citation2011, Frey et al. Citation2016, Brambilla et al. Citation2017b), and the investigation of fine-scale ecological patterns may increase our mechanistic understanding of how extensively organisms might adapt to environmental change (Baudier et al. Citation2015, Bennett et al. Citation2015, Brambilla et al. Citation2018). This may be crucial for accurate predictions of species’ responses to environmental or climatic impacts, and hence for the identification of conservation priorities. Studies on fine-scale habitat use by animal species are therefore required to better understand how species respond to climate change impacts (Scheffers et al. Citation2014, Pincebourde et al. Citation2016). However, so far only a few studies have investigated responses regulated by microhabitat/microclimate and their relative modifications, despite growing evidence highlighting their fundamental role (Turlure et al. Citation2010, Fedy & Martin Citation2011, Suggitt et al. Citation2011, Frey et al. Citation2016).
Fine-scale habitat selection and microhabitat use may indeed buffer against climate change impacts, but at the same time it can vary according to various biological and environmental factors (Valdez et al. Citation2017), to seasonality (Lunghi et al. Citation2015) or according to individual traits (Whiteman & Côté Citation2004, Morales et al. Citation2008), eventually resulting in different impacts of climate change according to different microhabitat use (Bowden et al. Citation2015). Hence, an understanding of fine-scale habitat selection is often essential to predict the potential impact of climate change on a species, and it can vary according to habitat composition on a broader scale (i.e. landscape). Therefore, assessing variations in the use of fine-scale habitats (with different sensitivity to climate change) according to landscape may also help to identify landscape-dependent vulnerability to climate change. This would greatly facilitate the identification of areas where species are more likely to be threatened or, vice-versa, to persist, on the basis of the landscape characteristics, which are easier to assess on a large-scale. Fine-scale habitat assessment requires in-depth investigation which is not always feasible over broad areas.
Mountain regions are particularly sensitive to climate change (Gobiet et al. Citation2014), and mountain bird species are considered to be at higher risk compared to birds from other habitats (Chamberlain et al. Citation2016, Scridel et al. Citation2017, Citation2018, Brambilla et al. Citation2017a). Earlier snowmelt, due to global warming, is one of the main threats for several species inhabiting areas at high-elevation or latitude, including plants (Kudo & Hirao Citation2006) and animals (Rosvold Citation2016). One of the species most threatened by climate change in the European Alps is the White-winged Snowfinch (hereafter Snowfinch) Montifringilla nivalis; according to correlative distribution models, it is projected to lose large portions of its current range due to climate warming (Brambilla et al. Citation2016, Citation2017a). Snowfinches make abundant use of snowfields and of their melting margins for prey collection: during the critical phase of nestling rearing, Snowfinches often forage on snow margins and snowfields (Strinella et al. Citation2007, Brambilla et al. Citation2017b, Citation2018). Snow margins provide abundant and accessible insect prey (Muscio et al. Citation2005), whereas on snowfields invertebrate detectability is highest, and Snowfinches forage on wind-blown arthropod fallout (Antor Citation1995). Snow cover and its duration are decreasing in mountain systems, including the Alps (Klein et al. Citation2016) and particularly in springtime (IPCC Citation2007, Brown & Robinson Citation2011, Derksen & Brown Citation2012). This implies that fine-scale habitats linked to snow cover (and the species relying on these) are particularly vulnerable. In addition, Snowfinches frequently forage on alpine grassland, especially on sites with short grass sward (Brambilla et al. Citation2017b), and correct grassland management is a key feature for preserving foraging habitat suitability for Snowfinches (Brambilla et al. Citation2018). The importance of snow for the species represents one of the potential mechanistic links with climate and the increasingly earlier snow-melt date in the Alps (Klein et al. Citation2016; see also Brown & Robinson Citation2011), suggests that climate-induced modifications of foraging habitats may be one of the most critical impacts of climate change on this species. Snowfinch populations or individuals relying mostly on snowfields and their margins could thus be at greater risk than those relying on other fine-scale habitats (e.g. grassland). With this study, we investigated whether landscape composition surrounding Snowfinch nests affected their fine-scale habitat selection during the capture of prey items when provisioning their nestlings. We hypothesized that the availability of alternative suitable habitats may determine a lower reliance on snow-related habitats, and thus that landscape composition may modulate extinction risk posed by habitat modifications induced by climate change.
Based on previous knowledge (Brambilla et al. Citation2017b), we identified five main foraging fine-scale habitats for Snowfinches: snow, grassland, bare substrate, snow-grassland margin, snow-bare substrate margin. These fine-scale habitats are likely subjected to different intensities of threat imposed by climate change. The availability of snow cover and margins is clearly affected by climate and impacted by rising temperatures and changes in precipitation regimes. Indeed, snow cover in the Alps has reduced over time, including its duration, with an approximately 6-day advance in snowmelt date per decade (Klein et al. Citation2016). These fine-scale habitats are most likely to decrease or disappear in future decades. On the other hand, grassland and bare substrate are likely to be less subject to the potential impact of climate change; nevertheless, climate change will affect grassland structure and composition (Theurillat & Guisan Citation2001; but see Cannone et al. Citation2007, Harsch et al. Citation2009). In this study, we focused on the use of snowfields and their margins in relation to landscape composition.
Methods
Study area and fieldwork
The study was carried out in the Italian Alps, in the provinces of Sondrio, Brescia, Trento and Bolzano, across five study sites located at different altitudes (1900–3200 m above sea level) and within different landscape characteristics. The average cover of habitats and snow (2–21%) varied greatly across study sites: grassland was dominant in two sites, bare substrates at another two, whereas the remaining one was characterized by a mixture of different habitats ().
Table 1. Number of surveyed pairs, foraging locations and surveys (number of days-per-pair at a given site) and average landscape composition (300 m radius around the nests) for the different study areas; mean values are shown because within-study variations occurred because of snowmelt progression. The category ‘other’ includes man-made areas and water bodies.
We investigated foraging habitat selection by breeding Snowfinches between the 7th June and the 19th July 2017. Seventeen breeding pairs were included within this study, 8 of which were followed for one day and 9 pairs for two days, with the interval between subsequent visits ranging between 2 and 10 days. During each survey, we monitored breeding Snowfinches, visually tracking adults when moving from the nest to a foraging location. We recorded habitat exploited at approximately 20 foraging events per pair/day (); for each pair we included a mean (± sd) of 28 ± 13 foraging events. Each foraging event was attributed to one of the following fine-scale habitat types: grassland, bare substrate, snowfield, margin snow-grassland, margin snow-bare substrate. Bare substrate included rocks, sand, gravel, scree, mud or bare soil.
For each breeding pair/date we estimated the proportional cover of different habitat types (i.e. grassland, snow, bare substrate, other – mostly man-made areas and waterbodies) within a 300 m radius around the nest, as available studies indicate that adults largely forage within this distance from the nest (Strinella et al. Citation2007, Brambilla et al. Citation2017b, Citation2018). A 300 m fixed radius was used for all pairs to get daily and comparable data on landscape composition around the nests. Habitat coverage was estimated visually in the field with the help of aerial photographs. We adopted this approach as available databases did not provide enough details to distinguish key habitat features at such a fine scale, such as small areas of grass or snow patches. Further to this, snow cover had to be assessed periodically and in some cases on a daily basis. To verify the accuracy of these visual estimates, we performed checks of coverage estimates using a GIS software (Geographical Information System; QGIS Development Team Citation2018) by redrawing habitat plots, and the results were similar to the visual estimates. For each foraging event, we also recorded meteorological conditions (sky cover: 0: clear, 1: partly overcasted, 2: overcasted (cover > 95%); wind: 0: absent, 1: weak, 2: moderate or strong), as weather conditions may affect invertebrate activity and availability to Snowfinches (Losapio et al. Citation2016).
Data analysis
We built generalized linear mixed models (GLMMs) to evaluate whether landscape composition, meteorological conditions and season progression affected fine-scale habitat selection for foraging Snowfinches. We built models to quantify the effects of the above factors on the use of snowfields and of snow-bare substrate margins versus all other habitats. We selected snowfields and snow-bare substrate margins as they are the most sensitive to climate change, being directly dependent only on spring snow cover, which is steadily decreasing in the Alps (Klein et al. Citation2016). Snowfinch pair identity was entered as random factor to take into account the non-independence of records. Factors entered in the model were: date (as Julian date), sky cover (factorial), wind (factorial), percentage cover in the 300 m radius of grassland, snow, bare substrate and other habitats. All variables were standardized before analysis. To avoid overfitting and possible biases due to multicollinearity, we first built three models, one for meteorological factors and date, and two for habitat cover variables. Given that some pairs of land-cover variables were correlated (snow and grass: r = −0.67, grass and bare substrate: r = −0.54), we tested their effect by means of two separate models, the first including snow and bare ground cover (r = −0.11), and the second grassland and other habitats (r = −0.41). For each one of the three groups, we performed a model selection procedure based on an information-theoretic approach (Burnham & Anderson Citation2002), ranking all possible models within a group according to the Akaike Information Criterion corrected for small sample sizes (AICc). Then, from each of the three procedures of model selection, we selected the variables comprised in the most parsimonious (ΔAICc < 2) models (excluding the ‘uninformative parameters’ sensu Arnold Citation2010), as done in other studies (Jedlikowski et al. Citation2016). With these variables we built a single GLMM relating the use of a given fine-scale habitat (snowfields or margins snow-bare) to the selected meteorological and landscape factors (Assandri et al. Citation2016). All possible models were again ranked according to AICc. We then calculated marginal (variance explained by fixed effects only) and conditional R2 (variance explained also by the random factor). All analyses were performed in R (R Development Core Team Citation2016) with packages lme4 (Bates et al. Citation2015) and MuMIn (Bartoń Citation2016). Variable effects according to the GLMM model were plotted with the package Visreg (Breheny & Burchett Citation2018).
Results
Data on 455 foraging locations were collected () of which 130 (28.6%) occurred on snowfields or snow-bare substrate margins (68 and 62, respectively). Most of the other records occurred in grassland habitat (36.5%) or at snow-grassland margins (30.5%).
The use of different fine-scale habitats was related to the landscape composition. A single most supported model was identified at the final step (all other models ΔAICc > 2; ). The use of snowfields and snow-bare margins decreased as the season progressed () and with grassland cover (), while it was positively affected by wind (proportionally to wind intensity; ). Marginal R2 of the model was equal to 0.29 (conditional R2 = 0.67) and there was no overdispersion.
Figure 1. The probability of use of snowfields and snow-bare margins by breeding Snowfinches during the nestling rearing period in relation to season progression.
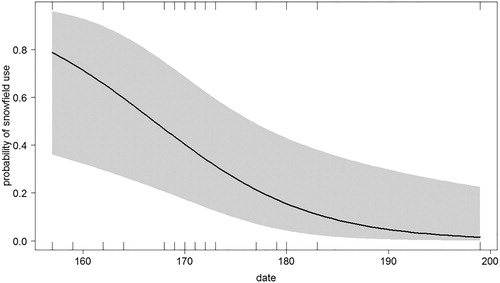
Figure 2. The probability of use of snowfields and snow-bare margins by breeding Snowfinches during the nestling rearing period in relation to grassland cover.
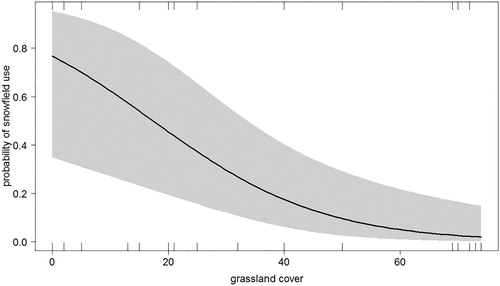
Figure 3. The probability of use of snowfields and snow-bare margins by breeding Snowfinches during the nestling rearing period in relation to wind intensity (moderate and strong wind considered together; see text).
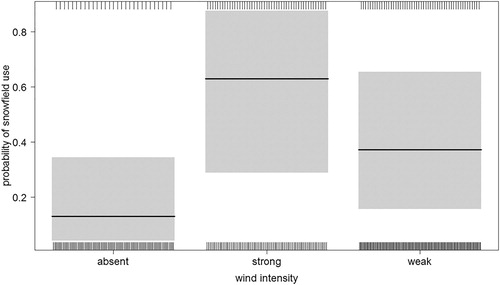
Table 2. Most supported (ΔAICc < 2) models for the use of snowfields and snow-bare substrate margins by breeding Snowfinches during the nestling rearing phase. For the categorical variables sky cover and wind, + indicates the inclusion in the model.
Discussion
Climate change is one of the main threats to biodiversity and represents a major pressure on several ecosystems and species over the coming decades (Scheffers et al. Citation2016). However, even for the species most likely to be negatively affected by climate change (such as mountain birds, Scridel et al. Citation2018), the impact is potentially multi-faceted and affected by abiotic or biotic factors, such as habitat/microhabitat characteristics, or the availability of key resources largely not dependent on climate conditions (Scheffers et al. Citation2014, Bowden et al. Citation2015, Brambilla et al. Citation2018). Species, populations or individuals that mostly rely on climate-dependent resources are the ones most at risk because of the direct impact of climate change. However, this topic has so far received little investigation, and we are not aware of studies focusing on the use of climate-dependent resources by animal species at a fine scale in relation to the broader environment they inhabit. Nevertheless, different degrees of reliance on such resources would result in different levels of resilience and vulnerability: species or individuals using a larger amount of other resources, which are not strictly dependent on climate, would display a higher resilience and a lower vulnerability to climate change.
In several Arctic and alpine environments, earlier snowmelt is a major driver of ecological change and could affect several species and ecosystems. Animal species relying on snowfields are thus particularly at risk (Rosvold Citation2016); they include the Snowfinch, which typically forages on or at the margin of snowfields, especially during the critical phase of nestling rearing (Brambilla et al. Citation2017b). The shorter duration of snow cover (Klein et al. Citation2016) and the lower availability of snowfields will dramatically reduce the availability of three of the fine-scale habitats exploited by Snowfinches during the crucial phase of its life cycle (Brambilla et al. Citation2018). Snowfields and the two types of snow margins hosted more than half of the foraging events recorded in our study. The negative effect of date on the probability of foraging on snowfields/snow-substrate margins could be due to changes in prey abundance or availability, or to a decrease in snow patches and/or an increase in grassland, with consequent increased use of the latter at the expense of snowfields. Other passerine species are known to use upland grassland at increasing elevation as the breeding season progresses, likely because of an increase in invertebrate availability in those habitats (Brambilla & Rubolini Citation2009).
According to our results, this impact of climate change will not affect all pairs/populations homogeneously, but the effects will be particularly important for those inhabiting areas dominated by bare substrates. In Snowfinches, the probability of foraging on snowfields or on snow-bare substrate margins rather than in other fine-scale habitats decreased with increasing grassland cover. Grassland occurrence reduces the dependency on snow, being a high-suitability alternative foraging habitat (Brambilla et al. Citation2017b, Citation2018), but in rocky landscapes, where grassland is scarcer or even absent, Snowfinches have to rely on snow-related fine-scale habitats. In fact, snowfields are likely to be suitable for foraging Snowfinches even in rocky environments, as they ‘trap’ invertebrate fallout. The importance of wind-carried invertebrates on snowfields (Antor Citation1995) is confirmed by the higher use Snowfinches make of snowfields with weak and, especially, moderate or strong wind (), when invertebrates are more likely to be blown by wind and deposited on snow. Data from the site dominated by bare substrate investigated in our study (Rosetta area) confirmed the importance of grassland and snow for Snowfinches. Thirty-eight out of 40 (95%) foraging events recorded for the two pairs breeding there were in grassland, snow or margin snow-grassland, even though the average cover of grassland and snow totalled only 19%. Only two foraging events occurred on bare substrates, despite an 81% cover of the landscape. Even grassland could be subjected to climate change impacts, for example, because of treeline upshift (Harsch et al. Citation2009), potentially affecting Snowfinch pairs breeding at the lowest parts of the species’ elevational range, or because of change in structural parameters, in particular sward height (Brambilla et al. Citation2018). Nevertheless, grassland is clearly less likely to be affected than snowfields by the direct impacts of climate change.
Pair identity explained a substantial portion of deviance in the model (conditional R2 definitely higher than marginal one; see Results): also local characteristics, social factors and/or individual traits are likely to contribute to the process of foraging habitat selection (Aplin et al. Citation2014), and hence may contribute to further modulate the potential response to environmental changes. Local characteristics may include height of the grassland sward (Brambilla et al. Citation2018), invertebrate abundance/availability in different habitats, habitat-specific disturbance or individual habituation to some specific patches (pers. obs.).
In conclusion, rock-dwelling Snowfinches will pay a higher cost to climate change, as the shortening of snow cover duration will result in the loss of most or all the suitable foraging fine-scale habitats during the most critical part of their life cycle. The conservation (and correct management, Brambilla et al. Citation2018) of alpine grassland, including the scarce grassland patches within sites dominated by bare substrates, is pivotal for the preservation of suitable foraging habitat for Snowfinches, under scenarios of predicted reduction in snow cover (Brown & Robinson Citation2011, Derksen & Brown Citation2012, Klein et al. Citation2016). However, if the overarching changes imposed by climate change in the long-term will disrupt ecological and food networks, even these small-scale habitats will lose suitability. Nevertheless, fine-scale environmental variations are essential for species’ responses to climate change and should be included in adaptation strategies (Bradbury et al. Citation2011), exactly as local and landscape traits must be considered in management plans (Buchanan et al. Citation2017).
We highlight how coupling patterns of resource selection at fine scales with broader scale habitat descriptors may help understand and predict the potential impact of climate change on wild species, by combining a mechanistic assessment of species-habitat relationships with landscape characteristics. This approach could be used to define and evaluate the potential effectiveness of planned conservation strategies, such as the identification of priority sites for conservation or useful measures to increase species resilience to climate change.
Acknowledgements
We are very grateful to M. Ramanzin, R. Ambrosini, E. Bassi, D. Faifer, G. Volcan, A. Franzoi, A. Iemma, L. Zane and to CAI/SAT, Baita Segantini, Rifugio Capanna Cervino, Rifugio G. Volpi al Mulaz, Rifugio Predotti alla Rosetta, Rifugio Pradidali, Rifugio Velo della Madonna e Funivia Rosetta, Hotel Perego, Hotel Pirovano Quarto, Rifugio Aldo Arnaldo Berni, Rifugio Bonetta, Albergo Folgore, for helpful collaboration. The study was funded by MUSE and Paneveggio-Pale di San Martino Natural Park as part of DS’s PhD. AF was supported by the Autonomous Province of Trento (Italy). D. Hayhow, an anonymous reviewer and the associate editor provided very helpful comments on a first draft of the manuscript.
ORCID
Mattia Brambilla http://orcid.org/0000-0002-7643-4652
References
- Antor, R.J. 1995. The importance of arthropod fallout on snow patches for the foraging of high-alpine birds. J. Avian Biol 26: 81–85. doi: 10.2307/3677216
- Aplin, L.M., Farine, D.R., Mann, R.P. & Sheldon, B.C. 2014. Individual-level personality influences social foraging and collective behaviour in wild birds. Proc. R. Soc. London B 281: 20141016. doi: 10.1098/rspb.2014.1016
- Arnold, T.W. 2010. Uninformative parameters and model selection using Akaike’s information criterion. J. Wildl. Manage 74: 1175–1178. doi: 10.1111/j.1937-2817.2010.tb01236.x
- Assandri, G., Bogliani, G., Pedrini, P. & Brambilla, M. 2016. Diversity in the monotony? Habitat traits and management practices shape avian communities in intensive vineyards. Agric. Ecosyst. Environ 223: 250–260. doi: 10.1016/j.agee.2016.03.014
- Bartoń, K. 2016. MuMIn: multi-model inference. R package version 1.15.6.
- Bates, D., Mächler, M., Bolker, B. & Walker, S. 2015. Fitting linear mixed-effects models using lme4. J. Stat. Softw 67: 1–48. doi: 10.18637/jss.v067.i01
- Baudier, K.M., Mudd, A.E., Erickson, S.C. & O’Donnell, S. 2015. Microhabitat and body size effects on heat tolerance: implications for responses to climate change (army ants: Formicidae, Ecitoninae). J. Anim. Ecol 84: 1322–1330. doi: 10.1111/1365-2656.12388
- Bennett, N.L., Severns, P.M., Parmesan, C. & Singer, M.C. 2015. Geographic mosaics of phenology, host preference, adult size and microhabitat choice predict butterfly resilience to climate warming. Oikos 124: 41–53. doi: 10.1111/oik.01490
- Bowden, J.J., Hansen, R.R., Olsen, K. & Høye, T.T. 2015. Habitat-specific effects of climate change on a low-mobility Arctic spider species. Polar Biol. 38: 559–568. doi: 10.1007/s00300-014-1622-7
- Bradbury, R.B., Pearce-Higgins, J.W., Wotton, S.R., Conway, G.J. & Grice, P.V. 2011. The influence of climate and topography in patterns of territory establishment in a range-expanding bird. Ibis (Lond. 1859) 153: 336–344. doi: 10.1111/j.1474-919X.2011.01106.x
- Brambilla, M., Caprio, E., Assandri, G., Scridel, D., Bassi, E., Bionda, R., Celada, C., Falco, R., Bogliani, G., Pedrini, P., Rolando, A. & Chamberlain, D. 2017a. A spatially explicit definition of conservation priorities according to population resistance and resilience, species importance and level of threat in a changing climate. Divers. Distrib 23: 727–738. doi: 10.1111/ddi.12572
- Brambilla, M., Cortesi, M., Capelli, F., Chamberlain, D., Pedrini, P. & Rubolini, D. 2017b. Foraging habitat selection by Alpine White-winged Snowfinches Montifringilla Nivalis during the nestling rearing period. J. Ornithol 158: 277–286. doi: 10.1007/s10336-016-1392-9
- Brambilla, M., Pedrini, P., Rolando, A. & Chamberlain, D.E. 2016. Climate change will increase the potential conflict between skiing and high-elevation bird species in the Alps. J. Biogeogr 43: 2299–2309. doi: 10.1111/jbi.12796
- Brambilla, M., Resano-Mayor, J., Scridel, D., Anderle, M., Bogliani, G., Braunisch, V., Capelli, F., Cortesi, M., Horrenberger, N., Pedrini, P., Sangalli, B., Chamberlain D., Arlettaz, R. & Rubolini, D. 2018. Past and future impact of climate change on foraging habitat suitability in a high-alpine bird species: management options to buffer against global warming effects. Biol. Conserv 221: 209–218. doi: 10.1016/j.biocon.2018.03.008
- Brambilla, M. & Rubolini, D. 2009. Intra-seasonal changes in distribution and habitat associations of a multi-brooded bird species: implications for conservation planning. Anim. Conserv 12: 71–77. doi: 10.1111/j.1469-1795.2008.00226.x
- Braunisch, V., Coppes, J., Arlettaz, R., Suchant, R., Zellweger, F. & Bollmann, K. 2014. Temperate mountain forest biodiversity under climate change: compensating negative effects by increasing structural complexity. PLoS One 9: e97718. doi: 10.1371/journal.pone.0097718
- Breheny, P. & Burchett, W. 2018. visreg: visualization of regression models.
- Brown, R.D. & Robinson, D.A. 2011. Northern hemisphere spring snow cover variability and change over 1922–2010 including an assessment of uncertainty. Cryosphere 5: 219–229. doi: 10.5194/tc-5-219-2011
- Buchanan, G.M., Pearce-Higgins, J.W., Douglas, D.J.T. & Grant, M.C. 2017. Quantifying the importance of multi-scale management and environmental variables on moorland bird abundance. Ibis (Lond. 1859) 159: 744–756. doi: 10.1111/ibi.12488
- Burnham, K.P. & Anderson, D.R. 2002. Model Selection and Multimodel Inference. Springer, New York, NY.
- Cannone, N., Sgorbati, S. & Guglielmin, M. 2007. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ 5: 360–364. doi: 10.1890/1540-9295(2007)5[360:UIOCCO]2.0.CO;2
- Chamberlain, D.E., Pedrini, P., Brambilla, M., Rolando, A. & Girardello, M. 2016. Identifying key conservation threats to alpine birds through expert knowledge. PeerJ. 4: e1723. PeerJ Inc. doi: 10.7717/peerj.1723
- Derksen, C. & Brown, R. 2012. Spring snow cover extent reductions in the 2008–2012 period exceeding climate model projections. Geophys. Res. Lett 39: n/a-n/a. doi: 10.1029/2012GL053387
- Fedy, B. & Martin, K. 2011. The influence of fine-scale habitat features on regional variation in population performance of alpine white-tailed ptarmigan. Condor 113: 306–315. doi: 10.1525/cond.2011.100070
- Frey, S.J.K., Hadley, A.S., Betts, M.G. & Robertson, M. 2016. Microclimate predicts within-season distribution dynamics of montane forest birds. Divers. Distrib 22: 944–959. doi: 10.1111/ddi.12456
- Gobiet, A., Kotlarski, S., Beniston, M., Heinrich, G., Rajczak, J. & Stoffel, M. 2014. 21st century climate change in the European Alps – A review. Sci. Total Environ 493: 1138–1151. doi: 10.1016/j.scitotenv.2013.07.050
- Harsch, M.A., Hulme, P.E., McGlone, M.S. & Duncan, R.P. 2009. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett 12: 1040–1049. doi: 10.1111/j.1461-0248.2009.01355.x
- Higgins, J.W.P., Dennis, P., Whittingham, M.J. & Yalden, D.W. 2010. Impacts of climate on prey abundance account for fluctuations in a population of a northern wader at the southern edge of its range. Glob. Chang. Biol 16: 12–23. Wiley/Blackwell (10.1111). doi: 10.1111/j.1365-2486.2009.01883.x
- IPCC. 2007. Impacts, Adaptation and Vulnerability, Working Group II Contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge.
- Jedlikowski, J., Chibowski, P., Karasek, T. & Brambilla, M. 2016. Multi-scale habitat selection in highly territorial bird species: exploring the contribution of nest, territory and landscape levels to site choice in breeding rallids (Aves: Rallidae). Acta Oecol. 73: 10–20. doi: 10.1016/j.actao.2016.02.003
- Klein, G., Vitasse, Y., Rixen, C., Marty, C. & Rebetez, M. 2016. Shorter snow cover duration since 1970 in the Swiss Alps due to earlier snowmelt more than to later snow onset. Clim. Change 139: 637–649. doi: 10.1007/s10584-016-1806-y
- Kudo, G. & Hirao, A.S. 2006. Habitat-specific responses in the flowering phenology and seed set of alpine plants to climate variation: implications for global-change impacts. Popul. Ecol 48: 49–58. doi: 10.1007/s10144-005-0242-z
- Losapio, G., Gobbi, M., Marano, G., Avesani, D., Boracchi, P., Compostella, C., Pavesi, M., Schöb, C., Seppi, R., Sommaggio, D., Zanetti, A. & Caccianiga, M. 2016. Feedback effects between plant and flower-visiting insect communities along a primary succession gradient. Arthropod. Plant. Interact 10: 485–495. doi: 10.1007/s11829-016-9444-x
- Lunghi, E., Manenti, R. & Ficetola, G.F. 2015. Seasonal variation in microhabitat of salamanders: environmental variation or shift of habitat selection? PeerJ. 3: e1122. PeerJ Inc. doi: 10.7717/peerj.1122
- Morales, M.B., Traba, J., Carriles, E., Delgado, M.P. & de la Morena, E.L.G. 2008. Sexual differences in microhabitat selection of breeding little bustards Tetrax tetrax: ecological segregation based on vegetation structure. Acta Oecol. 34: 345–353. doi: 10.1016/j.actao.2008.06.009
- Muscio, G., Pellegrini, G., Solari, M., Tomaselli, M., Vanin, S. & Zanetti, A. 2005. Ambienti nivali. La vita in un ambiente estremo, Quaderni H. Ministero dell’Ambiente e della Tutela del Territorio, Museo Friulano di Storia Naturale, Comune di Udine.
- Nieto-Sánchez, S., Gutiérrez, D. & Wilson, R.J. 2015. Long-term change and spatial variation in butterfly communities over an elevational gradient: driven by climate, buffered by habitat. Divers. Distrib 21: 950–961. doi: 10.1111/ddi.12316
- Otto, S.A., Diekmann, R., Flinkman, J., Kornilovs, G. & Möllmann, C. 2014. Habitat heterogeneity determines climate impact on Zooplankton community structure and dynamics. PLoS One 9: e90875. Public Library of Science. doi: 10.1371/journal.pone.0090875
- Pincebourde, S., Murdock, C.C., Vickers, M. & Sears, M.W. 2016. Fine-scale microclimatic variation can shape the responses of organisms to global change in both natural and urban environments. Integr. Comp. Biol, 56: 45–61.
- QGIS Development Team. 2018. QGIS geographic information system. QGIS geographic information system. Open source geospatial foundation project.
- R Development Core Team. 2016. A language and environment for statistical computing. R foundation for statistical computing, Vienna.
- Rosvold, J. 2016. Perennial ice and snow-covered land as important ecosystems for birds and mammals. J. Biogeogr 43: 3–12. doi: 10.1111/jbi.12609
- Scheffers, B.R., De Meester, L., Bridge, T.C.L., Hoffmann, A.A., Pandolfi, J.M., Corlett, R.T., Butchart, S.H.M., Pearce-Kelly, P., Kovacs, K.M., Dudgeon, D., Pacifici, M., Rondinini, C., Foden, W.B., Martin, T.G., Mora, C., Bickford, D. & Watson, J.E.M. 2016. The broad footprint of climate change from genes to biomes to people.
- Scheffers, B.R., Edwards, D.P., Diesmos, A., Williams, S.E. & Evans, T.A. 2014. Microhabitats reduce animal’s exposure to climate extremes. Glob. Chang. Biol 20: 495–503. doi: 10.1111/gcb.12439
- Scherrer, D. & Körner, C. 2011. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr 38: 406–416. doi: 10.1111/j.1365-2699.2010.02407.x
- Scridel, D., Bogliani, G., Pedrini, P., Iemma, A., Von Hardenberg, A. & Brambilla, M. 2017. Thermal niche predicts recent changes in range size for bird species. Clim. Res 73: 207–216. doi: 10.3354/cr01477
- Scridel, D., Brambilla, M., Martin, K., Lehikoinen, A., Iemma, A., Matteo, A., Jahnig, S., Caprio, E., Bogliani, G., Pedrini, P., Rolando, A., Arlettaz, R. & Chamberlain, D. 2018. A review and meta-analysis of the effects of climate change on Holarctic mountain and upland bird populations. Ibis 160: 489–515. doi: 10.1111/ibi.12585
- Selwood, K.E., Thomson, J.R., Clarke, R.H., McGeoch, M.A. & Mac Nally, R. 2015. Resistance and resilience of terrestrial birds in drying climates: do floodplains provide drought refugia? Glob. Ecol. Biogeogr 24: 838–848. doi: 10.1111/geb.12305
- Strinella, E., Ricci, F. & Vianale, P. 2007. Uso dell’habitat nel Fringuello alpino (Montifringilla nivalis) in periodo riproduttivo in un’area sub-antropizzata: Campo Imperatore (Gran Sasso-Abruzzo). Alula 14: 107–114.
- Suggitt, A.J., Gillingham, P.K., Hill, J.K., Huntley, B., Kunin, W.E., Roy, D.B. & Thomas C.D. 2011. Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos 120: 1–8. doi: 10.1111/j.1600-0706.2010.18270.x
- Theurillat, J.-P. & Guisan, A. 2001. Potential impact of climate change on vegetation in the European Alps: a review. Clim. Change 50: 77–109. doi: 10.1023/A:1010632015572
- Turlure, C., Choutt, J., Baguette, M. & Van Dyck, H.A.N.S. 2010. Microclimatic buffering and resource-based habitat in a glacial relict butterfly: significance for conservation under climate change. Glob. Chang. Biol 16: 1883–1893. doi: 10.1111/j.1365-2486.2009.02133.x
- Valdez, J.W., Klop-Toker, K., Stockwell, M.P., Fardell, L., Clulow, S., Clulow, J. & Mahony, M.J. 2017. Differences in microhabitat selection patterns between a remnant and constructed landscape following management intervention. Wildl. Res 44: 248. doi: 10.1071/WR16172
- Wessely, J., Hülber, K., Gattringer, A., Kuttner, M., Moser, D., Rabitsch, W., Schindler, S., Dullinger, S. & Essl, F. 2017. Habitat-based conservation strategies cannot compensate for climate-change-induced range loss. Nat. Clim. Chang 7: 823–827. doi: 10.1038/nclimate3414
- Whiteman, E.A. & Côté, I.M. 2004. Individual differences in microhabitat use in a Caribbean cleaning goby: a buffer effect in a marine species? J. Anim. Ecol 73: 831–840. doi: 10.1111/j.0021-8790.2004.00858.x