
ABSTRACT
Using geolocator-immersion loggers, we tracked for the first time the migration of one Cory’s Shearwater Calonectris borealis fledgling, from its breeding colony in the Canary Islands, and along its first year of life. The juvenile bird initially followed the same migratory path as the adults but visited different areas of the Central and the South Atlantic Ocean.
Since the nineties, the number of tracking studies has increased exponentially, and those directly connected to conservation have also increased steadily (Fraser et al. Citation2018). However, most studies focused on adult ecology and behaviour, whereas juvenile and immature individuals have been relatively neglected, mainly due to the difficulty in studying them before their first breeding attempt (Hazen et al. Citation2012).
In the marine environment, the distribution and abundance of seabirds, during the breeding or non-breeding seasons, are increasingly used to identify and manage relevant areas for the conservation of marine ecosystems (Lascelles et al. Citation2012). However, this process is mainly based on the movements of adults, because the spatial distribution of juvenile seabirds is often difficult to obtain due to an extended multi-year at-sea period following fledging (Phillips et al. Citation2017), which complicates the recapture of tagged birds. Nevertheless, seabirds in earlier life stages can differ significantly from adults in the spatial distribution at sea (Phillips et al. Citation2017) and may be more exposed to threats and experience higher mortality than adults in fisheries because of their naïve behaviour (Riotte-Lambert & Weimerskirch Citation2013, Fay et al. Citation2015).
Among pelagic seabirds, Calonectris shearwaters are relatively well-studied regarding the spatial ecology of their adult populations (details of the species are provided in online Appendix S1). In the Atlantic and the Mediterranean, they undertake long and rapid transoceanic migrations from their breeding grounds to their numerous wintering areas spread along the North, Central and South Atlantic (Reyes-González et al. Citation2017). Fledglings usually leave the colony from the middle to the end of October and do not return to the natal colony for breeding until they reach about nine years of age (Mougin et al. Citation2000). However, our current understanding of postnatal dispersion of these birds and their at-sea behaviour is rather limited to fledglings tracked for short periods of time (Raine et al. Citation2011, Péron & Grémillet Citation2013, Afán et al. Citation2019) and a single track of a 4 year old immature bird (Dias et al. Citation2011). Here, using miniaturized light-level geolocators, we report the first-year migration of one Cory’s Shearwater Calonectris borealis that fledged from its natal breeding population in the Canary Islands (Spain) in 2017. Our main aim is to understand whether the annual movements, the schedule and the at-sea activity patterns of this unique 10-month track of the juvenile differ from those of adults tracked in the same colony and year, thus providing the first evidence of individual exploratory behaviour in juvenile shearwaters.
The study was conducted in Gran Canaria, Canary Islands, Spain (15°47ʹ18ʺ N 27°50ʹ41ʺ W). In early October 2017, geolocaTION-immersion loggers (Intigeo-C330 from Migrate Technology Ltd, Cambridge, UK) were leg-mounted on 43 fledgling birds (online Figure S1). The weight of these geolocators (2.6 g) corresponded to 0.3–0.4% of bird body mass, thus well below the threshold of 3.0% above which detrimental effects are more likely to occur (Phillips et al. Citation2003). Despite their relative lack of accuracy (Phillips et al. Citation2004), geolocators are useful for identifying broad foraging areas throughout the annual cycle. Previously, and for comparative purposes, geolocator deployment on breeding adults was carried out during the incubation period (June–July). On 5 July 2018, one tagged juvenile Cory’s Shearwater was unexpectedly recovered dead on a beach in Florida, USA (29°12ʹ11ʺ N 80°59ʹ42ʺ W). Throughout the breeding season of 2018, we also recovered geolocators from 33 breeding adults at the colony.
The geolocators measured light intensity every minute, recording the maximum at 5-minute intervals. Twilight events from raw light intensities were visually supervised and computed with Intiproc software from Migrate Technology Ltd (Coton, Cambridge, UK; Fox Citation2015). To compute and model realistic positions for birds, we applied a probabilistic algorithm designed in R (probGLS package; Merkel et al. Citation2016), which used a four-step filter (including a land mask, travel speed [max 80 m s−1], behaviour [inferred from saltwater immersion data], and environmental [Sea Surface Temperature; SST] data). The algorithm computed stable results with uncertainty estimates, including around the equinoxes, and did not require calibration of solar angles. From migratory birds (i.e. we excluded resident birds), we estimated: (1) departure date from breeding area (i.e. the Canary Current), (2) date of crossing the Equator on their outward migrations and (3) date of crossing the Equator on their return migrations (). All dates were estimated using routines written in R, and then confirmed by visual inspection on the reconstructed tracks. Additionally, geolocators had saltwater immersion sensors recording contact with saltwater every 3 s. From these immersion values, we derived the activity patterns and matched them with the light data to calculate the proportion of daily time spent on the water and in flight during darkness and daylight throughout the tracking period (Dias et al. Citation2012, Ramos et al. Citation2016). For plotting purposes, activity budgets were modelled using generalized additive mixed models (GAMMs) through the library mgcv in R (Wood Citation2017). While modelling daily resting time year-round for adults, we included migratory strategy (migratory or resident) as a fixed term, timing throughout the annual cycle as a smooth term, and bird identity as a random term. Similarly, for modelling circadian activity, we included timing throughout the day as a smooth term, and bird identity as a random term. This allowed us to determine for the juvenile and adult birds the general behavioural patterns throughout the annual cycle, as well as circadian behavioural patterns in specific areas of the Atlantic Ocean.
Table 1. Comparison of migratory parameters between one juvenile and 26 adults of Cory’s Shearwaters from Gran Canaria, Canary Islands, Spain. Departure from the colony is only estimated for the juvenile as it relies on light data, which was not available for adults because, excluding incubation, they only attend burrows during night-time.
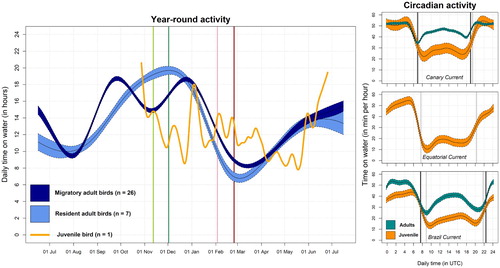
The tagged juvenile Cory’s Shearwater left its natal colony on 30 October 2017 and moved southward, spending a few days in the Canary Current and around Cape Verdean waters ((a)). This juvenile bird shared habitat in space and time with adult birds in the Canary Current, but only for a short period of time (for details see Appendix S1). Adult shearwaters use this productive upwelling area for moulting a number of wing feathers at the end of the breeding season (around 1 October; Ramos et al. Citation2009). This moult affects the flying capacity of birds during September and October (Alonso et al. Citation2009) as also revealed by the daily time spent on the water for adults (). On the contrary, fledgling birds, with fresh plumage, do not have such flight impairment, and can, therefore, afford to spend more time flying (). These differences become more obvious during daylight, when inexperienced birds should benefit from daylight for foraging and finding prey in a novel environment. Similar differences between the juvenile and adults can be seen in the other shared area, the Brazil Current. Thus, the higher flight activity of the tagged juvenile, compared with that of the adults in the two common foraging areas, suggests that the absence of wing moult reported on juveniles of most Procellariform species during the first year of life (Huettmann & Diamond Citation2000, Bugoni & Furness Citation2009) may enable juveniles to invest more hours foraging to compensate for their low foraging efficiency.
Figure 1. Spatial distribution of one juvenile (a) and adult bird (b) of Cory’s Shearwaters from Gran Canaria, Canary Islands, Spain, tracked with geolocators throughout the annual cycle. (a) Track of the juvenile from its natal colony to the beach where it was found dead (Florida, USA: 29°12′11″N 80°59′42″W). The bird fledged on 30 October 2017, and beached on 06 July 2018. (b) Schematic annual phenology and distribution of 33 adult Cory’s Shearwaters. Main phenological events are plotted clockwise in the upper circle: breeding period in dark red, outward migration in light green, wintering period in dark green, and return migration in pink. Coloured areas are minimum convex polygons of bird positions during the breeding and wintering periods (in dark red and dark green, respectively). The main wintering areas were associated with Benguela and Agulhas Currents (n = 22 individuals), Brazil-Malvinas confluence region (n = 2), and Canary Current (n = 7). Additionally, outward individual trajectories of the migratory adult birds (modelled using positions 6 days after leaving the breeding area) are plotted as green lines. Exclusive Economic Zones (EEZs) are also shown in white dashed lines and sampled colony is depicted with a yellow star on both figures.
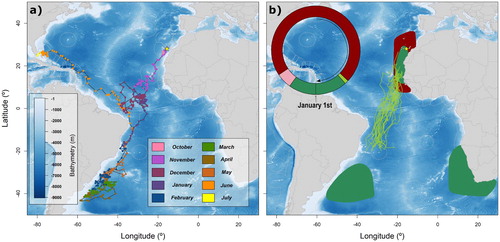
Figure 2. Daily resting time year-round (left-hand panel) and circadian activity (right-hand panels) of one juvenile and 33 adult Cory’s Shearwaters. Estimates of daily time on the water are plotted separately for each migratory strategy of adults (resident in light blue and migratory in dark blue), and for the juvenile bird (orange). For the curves of adult daily activity budgets, the solid lines correspond to the mean for each treatment group estimated using GAMMs and the coloured regions around the means show the associated 95% confidence intervals. The juvenile activity is represented by a smoothed curve. As references, coloured lines corresponding to average dates of the main phenological events estimated for those migratory adult birds are also plotted in the background: onset of the outward migration (light green), start of the wintering period (dark green), onset of the return migration (pink), and staging in the breeding ground (dark red). Circadian resting activity of the juvenile shearwater (in orange) in the three specific areas used by the bird: the Canary Current (25 days), the Equatorial Atlantic Current (59 days), and the Brazil Current (182 days). Circadian activity is also modelled using GAMMs and shown as time spent on the water. For reference, we plotted (when available) adult activity in those respective areas (in blue). Note that juvenile and adult activity do not represent the same periods of the annual cycle. Vertical lines correspond to mean of daily sunrise and sunset timings of each dataset (in light and dark grey for the juvenile and adults, respectively).
Our results indicate that juvenile and adult Cory’s Shearwaters could leave the breeding grounds simultaneously (), although previous reports on Scopoli’s Shearwaters Calonectris diomedea suggested differently (Péron & Grémillet Citation2013). Previous studies already pointed out that Cory’s Shearwaters cross the Equator in their outward migrations through a narrow corridor centred at 20° W that, in terms of energetic costs due to wind intensity, represents the lowest cost pathway (Felicísimo et al. Citation2008). In our case, both the juvenile and adult outward trajectories closely resembled that minimum wind cost trajectory moving southwest (). Therefore, our results suggest that although juveniles have limited initial orientational abilities, they might compensate for this by either following the adults during the first days post-fledging, or at least following the same general cues than adults do while migrating.
After its first long-distance movement, the juvenile bird stayed, between November and January, in a vast area of the Central Atlantic Ocean, clearly influenced by the Equatorial Counter-current and limited by Atlantic North and South Equatorial Currents. These tropical oceanic waters, only inhabited by smaller shearwater and petrel species, such as Pterodroma petrels (Ramos et al. Citation2017), Bulwer’s Petrel Bulweria bulwerii (Ramos et al. Citation2015), and Boyd’s Shearwater Puffinus boydi (Zajková et al. Citation2017), show typically low productivity and resource predictability. In addition, this area represents a narrow belt of calm winds around the Equator (Felicísimo et al. Citation2008), something that could act as an environmental trap for inexperienced seabirds. Indeed, no adult Calonectris shearwater uses this area throughout their annual cycles. Thus, according to the ‘exploration-refinement’ hypothesis (Guilford et al. Citation2011), we believe this might be the first challenge this young bird faced, and the first evidence that naïve young shearwaters could learn progressively the optimal migration routes towards more profitable wintering areas.
After crossing the Equator months later than the adults did, the juvenile bird kept moving southwards, following the edge of the South American continental shelf. From February to May, long after the breeding adults had returned to the Northern Hemisphere, the bird stayed in the Brazil Current area (including part of the South Atlantic Current). This warm water current was also exploited by the adult population (four tracked birds spent different periods of time in the area), although none of the adults temporally overlapped with the long stay of the juvenile shearwater. This area is also exploited by Scopoli’s Shearwaters (González-Solís et al. Citation2007), and by the entire population of Cape Verde Shearwater Calonectris edwardsii (González-Solís et al. Citation2009), highlighting the habitat suitability for the juvenile. However, adult populations of such closely-related species did not temporally overlap with this juvenile shearwater. This temporal mismatch might represent further evidence of the occurrence of an early learning process of juvenile birds to understand the best wintering spots through an individual exploratory behaviour of the Atlantic (Guilford et al. Citation2011).
In late May, when breeders start laying their egg at the colony, the juvenile bird returned to the Northern Hemisphere along the coast of Brazil ((a)). Contrary to what happened on its way south, the juvenile moved north against the prevailing winds, thus travelling rather inefficiently in terms of the energetic costs (Felicísimo et al. Citation2008, González-Solís et al. Citation2009). The decision to return to the Northern Hemisphere, albeit long after all adults had left the area, could be attributed to the annual migratory instinct of the species.
Once in the Northern Hemisphere, the bird spent June moving west towards the Lesser and Greater Antilles, where, apparently, no adult bird occurs throughout the year (). Typically, adults returning from their wintering grounds and moving towards their breeding grounds close a figure-of-eight migratory pattern, performing the clockwise loop in the Northern Hemisphere, sometimes spending a few days in the North Atlantic Ocean, but earlier in the year and far from the area the juvenile was apparently moving towards (González-Solís et al. Citation2007). Significant numbers of non-breeding birds can be seen regularly along North American waters from June to August, and most of them are assumed to be immature birds (Haney & McGillivary Citation1985, Gjerdrum et al. Citation2018). However, because the juvenile appeared dead a few weeks later on the Florida Peninsula, this last movement should be interpreted with caution, as its poor body condition (Appendix S1) could have influenced its last movements. The low level of activity during this last period further supports the hypothesis of a bird in a deteriorating body condition (). In addition, the tropical storm affecting the western Atlantic at that time (Hurricane Chris, in early July 2018) could have also played a definitive role in the death of such an inexperienced bird. This casualty and its associated uncertainty highlight the need for a much better understanding of the distribution and abundance of juvenile seabirds at sea, and especially the factors influencing their early mortality (Daunt et al. Citation2007, Fay et al. Citation2015).
Here, we report, for the first time, the trans-equatorial movements of a juvenile Cory’s Shearwater during its first year of life. This represents the first evidence of potential segregation between juvenile and adult populations of the species, something that has tremendous relevance for marine conservation. Finally, the dissimilarities found between the spatial ecology of this inexperienced juvenile bird with that of the adult population provide the first evidence of the early learning process of individual exploratory behaviour in juvenile shearwaters during their first years of life, supporting the exploration-refinement hypothesis.
Supplemental Material
Download MS Word (17.9 MB)Acknowledgements
We thank the Marine Science Center for recovering the geolocator in Florida and for performing the necropsy on the juvenile bird. RR was supported by a postdoctoral contract of the PLEAMAR program from the Spanish MCI (2017/2349), VMP was supported by a predoctoral contract of the Spanish MINECO (BES-2014-068025) and MCF was supported by a predoctoral grant from Universitat de Barcelona (D/134000500/P2017G/G00). We also thank Consejería de Medio Ambiente of Cabildo de Gran Canaria for permission to conduct the experiments (2015/1170).
ORCID
Raül Ramos http://orcid.org/0000-0002-0551-8605
Additional information
Funding
Related Research Data
References
- Afán, I., Navarro, J., Grémillet, D., Coll, M. & Forero, M.G. 2019. Maiden voyage into death: are fisheries affecting seabird juvenile survival during the first days at sea? R. Soc. Open. Sci. 6: 181151. doi: 10.1098/rsos.181151
- Alonso, H., Matias, R., Granadeiro, J.P. & Catry, P. 2009. Moult strategies of Cory’s Shearwaters Calonectris diomedea borealis: The influence of colony location, sex and individual breeding status. J. Ornithol. 150: 329–337. doi: 10.1007/s10336-008-0354-2
- Bugoni, L. & Furness, R.W. 2009. Ageing immature Atlantic Yellow-nosed Thalassarche chlororhynchos and Black-browed T. melanophris albatrosses in wintering grounds using bill colour and moult. Mar. Ornithol. 37: 249–252.
- Daunt, F., Afanasyev, V., Adam, A., Croxall, J.P. & Wanless, S. 2007. From cradle to early grave: juvenile mortality in European shags Phalacrocorax aristotelis results from inadequate development of foraging proficiency. Biol. Lett. 3: 371–374. doi: 10.1098/rsbl.2007.0157
- Dias, M.P., Granadeiro, J.P., Phillips, R.A., Alonso, H. & Catry, P. 2011. Breaking the routine: individual Cory’s shearwaters shift winter destinations between hemispheres and across ocean basins. Proc R. Soc. B: Biol. Sci 278: 1786–1793. doi: 10.1098/rspb.2010.2114
- Dias, M.P., Granadeiro, J.P. & Catry, P. 2012. Working the day or the night shift? Foraging schedules of Cory’s shearwaters vary according to marine habitat. Mar. Ecol. Prog. Ser. 467: 245–252. doi: 10.3354/meps09966
- Fay, R., Weimerskirch, H., Delord, K. & Barbraud, C. 2015. Population density and climate shape early-life survival and recruitment in a long-lived pelagic seabird. J. Anim. Ecol. 84: 1423–1433. doi: 10.1111/1365-2656.12390
- Felicísimo, A.M., Muñoz, J. & González-Solis, J. 2008. Ocean surface winds drive dynamics of transoceanic aerial movements. Plos One 3: e2928. doi: 10.1371/journal.pone.0002928
- Fox, J. 2015. Intiproc geolocation processing software. In User Reference Manual Intiproc v1.03 pMigrate Technology Ltd, Cambridge.
- Fraser, K.C., Davies, K.T.A., Davy, C.M., Ford, A.T., Flockhart, D.T.T. & Martins, E.G. 2018. Tracking the conservation promise of movement ecology. Front. Ecol. Evol. 6: 150. doi: 10.3389/fevo.2018.00150
- Gjerdrum, C., Loch, J. & Fifield, D.A. 2018. The recent invasion of Cory’s shearwaters into Atlantic Canada. Northeast. Nat. 25: 532–544. doi: 10.1656/045.025.0402
- González-Solís, J., Croxall, J.P., Oro, D. & Ruiz, X. 2007. Trans-equatorial migration and mixing in the wintering areas of a pelagic seabird. Front. Ecol. Environ. 5: 297–301. doi: 10.1890/1540-9295(2007)5[297:TMAMIT]2.0.CO;2
- González-Solís, J., Felicísimo, A., Fox, J.W., Afanasyev, V., Kolbeinsson, Y. & Muñoz, J. 2009. Influence of sea surface winds on shearwater migration detours. Mar. Ecol. Prog. Ser. 391: 221–230. doi: 10.3354/meps08128
- Guilford, T., Freeman, R., Boyle, D., Dean, B., Kirk, H., Phillips, R. & Perrins, C. 2011. A dispersive migration in the Atlantic puffin and its implications for migratory navigation. Plos One 6: e21336. doi: 10.1371/journal.pone.0021336
- Haney, J.C. & McGillivary, P.A. 1985. Aggregations of Cory’s shearwaters (Calonectris diomedea) at Gulf Stream fronts. Wilson Bull. 97: 191–200.
- Hazen, E.L., Maxwell, S.M., Bailey, H., Bograd, S.J., Hamann, M., Gaspar, P., Godley, B.J. & Shillinger, G.L. 2012. Ontogeny in marine tagging and tracking science: Technologies and data gaps. Mar. Ecol. Prog. Ser. 457: 221–240. doi: 10.3354/meps09857
- Huettmann, F. & Diamond, A.W. 2000. Seabird migration in the Canadian northwest Atlantic Ocean: moulting locations and movement patterns of immature birds. Can. J. Zool. 78: 624–647. doi: 10.1139/z99-239
- Lascelles, B.G., Langham, G.M., Ronconi, R.A. & Reid, J.B.. 2012. From hotspots to site protection: identifying Marine Protected Areas for seabirds around the globe. Biol. Conserv. 156: 5–14. doi: 10.1016/j.biocon.2011.12.008
- Merkel, B., Phillips, R.A., Descamps, S., Yoccoz, N.G., Moe, B. & Strøm, H. 2016. A probabilistic algorithm to process geolocation data. Mov. Ecol. 4: 26. doi: 10.1186/s40462-016-0091-8
- Mougin, J.-L., Jouanin, C. & Roux, F. 2000. Démographie du puffin cendré Calonectris diomedea de Selvagem Grande (30° 09’ N, 15° 52’ W). Revue d’Ecologie (La Terre et la Vie) 55: 275–290.
- Péron, C. & Grémillet, D. 2013. Tracking through life stages: adult, immature and juvenile autumn migration in a long-lived seabird. Plos One 8: e72713. doi: 10.1371/journal.pone.0072713
- Phillips, R.A., Xavier, J.C. & Croxall, J.P. 2003. Effects of satellite transmitters on albatrosses and petrels. Auk 120: 1082–1090. doi: 10.1093/auk/120.4.1082
- Phillips, R.A., Silk, J.R.D., Croxall, J.P., Afanasyev, V. & Briggs, D.R. 2004. Accuracy of geolocation estimates for flying seabirds. Mar. Ecol. Prog. Ser. 266: 265–272. doi: 10.3354/meps266265
- Phillips, R.A., Lewis, S., González-Solís, J. & Daunt, F. 2017. Causes and consequences of individual variability and specialization in foraging and migration strategies of seabirds. Mar. Ecol. Prog. Ser. 578: 117–150. doi: 10.3354/meps12217
- Raine, A., Borg, J.J. & Raine, H. 2011. First description of post-fledging migration of Maltese Cory’s Shearwaters Calonectris diomedea diomedea. Ringing Migr. 26: 114–117. doi: 10.1080/03078698.2011.587241
- Ramos, R., González-Solís, J. & Ruiz, X. 2009. Linking isotopic and migratory patterns in a pelagic seabird. Oecologia 160: 97–105. doi: 10.1007/s00442-008-1273-x
- Ramos, R., Sanz, V., Militão, T., Bried, J., Neves, V.C., Biscoito, M., Phillips, R.A., Zino, F. & González-Solís, J. 2015. Leapfrog migration and habitat preferences of a small oceanic seabird, Bulwer’s petrel (Bulweria bulwerii). J. Biogeogr. 42: 1651–1664. doi: 10.1111/jbi.12541
- Ramos, R., Ramírez, I., Paiva, V.H., Militão, T., Biscoito, M., Menezes, D., Phillips, R.A., Zino, F. & González-Solís, J. 2016. Global spatial ecology of three closely-related gadfly petrels. Sci. Rep. 6: 23447. doi: 10.1038/srep23447
- Ramos, R., Carlile, N., Madeiros, J., Ramírez, I., Paiva, V.H., Dinis, H.A., Zino, F., Biscoito, M., Leal, G., Bugoni, L., Jodice, P.G.R., Ryan, P.G. & González-Solís, J. 2017. It is the time for oceanic seabirds: tracking year-round distribution of gadfly petrels across the Atlantic Ocean. Divers. Distrib. 23: 794–805. doi: 10.1111/ddi.12569
- Reyes-González, J.M., Zajková, Z., Morera-Pujol, V., de Felipe, F., Militão, T., Dell’Ariccia, G., Ramos, R., Igual, J.M., Arcos, J.M. & González-Solís, J. 2017. Migración y ecología espacial de las poblaciones españolas de pardela cenicienta. SEO/BirdLife, Madrid, Spain.
- Riotte-Lambert, L. & Weimerskirch, H. 2013. Do naive juvenile seabirds forage differently from adults? Proc. R. Soc., Ser. B, Biol. Sc. 280: 20131434. doi: 10.1098/rspb.2013.1434
- Wood, S. 2017. mgcv: Mixed GAM Computation Vehicle with Automatic Smoothness Estimation. R package version 1.8-22. https://cran.r-project.org/web/packages/mgcv/mgcv.pdf.
- Zajková, Z., Militão, T. & González-Solís, J. 2017. Year-round movements of a small seabird and oceanic isotopic gradient in the tropical Atlantic. Mar. Ecol. Prog. Ser. 579: 169–183. doi: 10.3354/meps12269