
ABSTRACT
Capsule: There is seasonal variation in the benefits that birds can take from restored farmland ponds.
Aims: To evaluate season-specific benefits of farmland pond restoration for local bird communities.
Methods: Bird communities were recorded at unmanaged overgrown and managed open-canopy farmland ponds over the breeding, post-breeding and winter seasons. Results were compared and related to seasonal variation in environmental conditions of within-pond and marginal habitats to identify predictors of local bird communities.
Results: Bird communities at managed open-canopy ponds showed a higher abundance and species richness over all seasons and displayed pronounced seasonal shifts in composition. Warblers and other specialised bird species were frequently observed at open-canopy sites over the breeding and post-breeding seasons but were generally absent from overgrown ponds. While pond management and landscape connectivity had a consistent positive influence on bird communities over all seasons, the importance of other predictors such as bramble area varied seasonally.
Conclusions: Our study highlights a key role of pond management for farmland bird conservation. In addition, the identified seasonal predictors of bird assemblages provide valuable lessons for the design of agri-environment prescriptions for farmland ponds, highlighting the importance of bramble-dominated patches and pond marginal habitat over the breeding season and of a strong connectivity between pond margins and surrounding semi-natural habitats throughout the year.
Across Europe, many farmland bird species have declined (Tucker & Heath Citation1994) with an estimated loss of 420 million individuals since 1980 (Inger et al. Citation2015). In line with European declines, since 1970, UK farmland bird populations have decreased by 56% (Defra Citation2018). Agricultural intensification has impacted on farmland bird populations through a variety of mechanisms, ranging from changes in farming practices, such as the transition from spring-sown to autumn-sown crops (Evans et al. Citation2004, Newton Citation2004), intensified grassland management (Butler et al. Citation2010), increased use of agricultural chemicals (Bright et al. Citation2008, Hallmann et al. Citation2014), reduction of non-cropped features (Robinson & Sutherland Citation2002, Bright et al. Citation2008, Sayer et al. Citation2012) and increased land drainage (Newton Citation2004). Despite the widespread introduction of agri-environmental schemes (AES) by the European Union (EU) as a key component of the Common Agricultural Policy (CAP) in part to address declines in farmland biodiversity (Natural England Citation2009, Davey et al. Citation2010), populations of many farmland bird species have continued to decline. However, schemes that target key habitat provision, such as the Cirl Bunting Emberiza cirlus reintroduction project, have been shown to be locally effective in increasing bird populations (Aebischer et al. Citation2015, Jeffs et al. Citation2016). One AES measure that has had a poor uptake in the UK is pond management (Natural England Citation2009). Over the last 30–40 years, due to a reduction in traditional farmland pond management practices combining occasional scrub and sediment removal, many ponds have become highly terrestrialised. As a consequence numbers of early and mid-succession open-canopy farmland ponds have dwindled (Sayer et al. Citation2012, Sayer Citation2014, Thornhill Citation2017). Recent research has revealed that restoration and subsequent management of UK farmland ponds by sediment and scrub removal to re-establish and maintain open-canopy conditions promotes significant increases in the abundance and diversity of aquatic macrophytes and invertebrates (Sayer et al. Citation2012, Sayer et al. Citation2013). Furthermore, Davies et al. (Citation2016) and Lewis-Phillips et al. (Citation2019) demonstrated that bird abundance and overall species richness were significantly higher at managed open-canopy agricultural ponds than at their unmanaged overgrown counterparts in mid-summer and year-around surveys, respectively.
Farmland birds are highly mobile organisms (Siriwardena et al. Citation2006) that prioritise different resources at different times of the year (Vickery et al. Citation2009) and switch between breeding and winter feeding sites within the wider landscape (Aebischer et al. Citation2015). In order to fully understand the potential contribution of AES measures for birds, it is therefore important to evaluate the localised effects of these measures during different times of the year (Siriwardena et al. Citation2006, Redhead et al. Citation2018), and especially to separate patterns for breeding and non-breeding periods as associated with major bird diet shifts (Wilson et al. Citation1999).
To fill this research gap, this study investigates the local effect of farmland pond management for bird communities during the breeding, post-breeding and winter seasons. We specifically test the hypotheses that: (a) managed open-canopy ponds harbour a significantly higher bird species richness and abundance than unmanaged overgrown ponds over all seasons, reflecting a higher availability of resources all year around; (b) bird communities vary seasonally in response to environmental variations in pond and pond margin environments and (c) conservation priority bird species are recorded at higher abundances over all seasons at managed open-canopy ponds in comparison to unmanaged overgrown ponds.
Methods
Study site
We studied 16 agricultural ponds situated in North Norfolk, eastern England (Lewis-Phillips et al. Citation2019) (online supplementary Table S1, Figure S1). Ponds fell into two distinct categories: ‘managed open-canopy’ ponds and ‘unmanaged overgrown’ ponds. The open-canopy ponds had either been restored by major scrub and sediment removal (n = 8) or managed by more moderate scrub removal (n = 8) within the last five years. These ponds had low shading (<10%) and were all dominated by aquatic macrophytes. By contrast, the unmanaged overgrown ponds had not been managed for at least 20–40 years, and as a result had >85% shaded water, and a general absence of aquatic macrophytes. The unmanaged overgrown ponds were predominantly overgrown by Willow Salix spp. and European Alder Alnus glutinosa trees and also contained large quantities of dead wood (Sayer et al. Citation2012, Sayer et al. Citation2013). All study ponds were situated within agricultural arable fields and featured an herbaceous margin >7 m wide. The ponds were shallow (mean depth <1.5 m), had a mean (±sd) water surface area of 303 ± 31 m2, and a total mean (±sd) footprint, including pond margins, of 2694 ± 464 m2. A minimum distance of 200 m between pond sites was maintained to reduce spatial autocorrelation (Ralph et al. Citation1995).
Bird surveys
All bird species observed around the agricultural pond sites were included within this study. For analysis we differentiated between three major ecological seasons for birds: April to June as ‘breeding season’ (BTO Citation2011, Redhead et al. Citation2018), December to February as ‘winter season’ (Redhead et al. Citation2018), with the inclusion of a ‘post-breeding season’ from July to September.
Bird surveys were undertaken between May 2016 and April 2017 (Lewis-Phillips et al. Citation2019). One series of morning (05:00–10:30) and afternoon (12:00–17:30) surveys were undertaken on a monthly basis. Each bird survey series consisted of three ‘main’ and three ‘snapshot’ surveys. ‘Main’ surveys consisted of five-minute length point-count surveys, with a two-minute interval between each survey (Voříšek et al. Citation2008). The ‘Main’ surveys recorded all bird species with the exception of the aerial insectivores, Barn Swallow Hirundo rustica, Common Swift Apus apus and Common House Martin Delichon urbicum. The ‘snapshot’ surveys were aimed specifically at the aforementioned aerial insectivore species and took place exactly one minute after each ‘main’ survey was completed. To avoid the potential influence of poor weather on bird activity and behaviour, monitoring was not conducted in heavy rain or wind. Study site visit order was randomised on each occasion. To reduce bias due to variation in the detectability of different species, alongside potential detectability differences between the open-canopy and overgrown farmland ponds, bird individuals were recorded using either telescope/binoculars or by sound from a set location that maximised visibility of the pond open surface area and margin (Bibby et al. Citation1992). Individuals were identified to species level along with their initial location within the pond. Bird conservation status, categorised as ‘green’ (species of least conservation concern), ‘amber’ (species that have experienced moderate declines) and ‘red’ (species that have experienced severe population declines) (for further details see RSPB Citation2018) were also recorded. Birds initially flushed on arrival were included within the initial ‘main’ survey (Voříšek et al. Citation2008). Birds observed within the pond or surrounding herbaceous margin were recorded, but flying individuals were only counted if seen within a height of 10 m above the pond surface.
Pond habitat analysis
To determine the environmental setting of each site, we measured pond habitats during the bird survey year (2017) to an accuracy of one metre, including pond connectivity (distance to nearest semi-natural terrestrial habitat), total pond area, area of non-shaded water, area of shaded water, pond area covered by aquatic macrophytes, area of herbaceous margin, bramble area within the pond margin (defined as dense understory vegetation <3 m tall, mainly dominated by bramble Rubus spp., but sometimes also including Hawthorn Crataegus monogyna and Blackthorn Prunus spinosa), and area of trees within the pond margin (defined as tree species >3 m tall). Aerial images of each study pond were recorded using a DJI Mavik Pro unmanned aerial vehicle in combination with a scale marker. Habitat areas were then calculated using the scale tool in Photoshop Creative Cloud 2017 and subsequently updated for the three selected seasons. Pond connectivity was calculated as the distance from the centre of the pond and the nearest semi-natural terrestrial habitat feature (typically a hedgerow or woodland patch), using online Ordnance Survey maps and verified via field surveys. At unmanaged overgrown sites, the area of shaded water was also verified through field survey.
Data analysis
Bird community analysis
Following Lewis-Phillips et al. (Citation2019), bird abundance was measured as the overall number of bird recordings at each study site, combined across species and survey types. Species richness, Simpson’s exponential and Shannon’s diversity indices were used to calculate seasonal α-diversity and γ-diversity (Crist et al. Citation2003, Jost Citation2006). Flocks and families of birds were recorded as a single visit to prevent potential statistical bias generated by artificially increasing sample sizes (e.g. a single-species flock of 10 birds arriving together was counted as one visit, as was a single bird arriving alone). The ‘vegan’ package (Oksanen et al. Citation2013) in R software 3.5.2. (R Core Team Citation2017) was used to calculate the diversity indices. All further analyses were also completed in R software 3.5.2. (R Core Team Citation2017).
Our analysis mainly focuses on assemblage-level bird indices, based on the assumption that the benefits of pond management would be displayed through net increases in abundance or species richness. We acknowledge that individual species show species-specific response patterns to certain environmental variables, such as bramble area and, therefore, we provide a further analysis of Common Linnet Linaria cannabina (as an obligate granivorous specialist) and aerial insectivore abundance, alongside abundances of conservation priority red and amber listed species. Mann–Whitney tests were used to compare bird abundance and species richness alongside the abundance of Linnet, aerial insectivores and conservation priority species, between pond management types for each season.
Environmental predictors of seasonal bird communities
Pearson’s correlations between the environmental predictors were calculated for each season and individual variables from pairs with correlation coefficients >0.7 were removed (Booth et al. Citation1994, Dormann et al. Citation2013). Subsequent analyses were then conducted based on the remaining variables (Bates & Maechler Citation2010).
For each season, bird abundance and species richness were analysed using generalised linear mixed models (GLMMs) of assemblage-level indices. We modelled effects using a Poisson distribution applying a log-link function. Study pond and ‘pond nested within date’ were included as random effects, using the lme4 package (Bates & Maechler Citation2010) in R-software package 3.5.2. (R Core Team Citation2017), controlling potential temporal autocorrelation connected with repeat survey visits to study sites, in combination with multiple observations per date-pond combination. The difference in Akaike’s information criterion corrected for small sample sizes (ΔAICc < 2) was used to select models (Burnham & Anderson Citation2002, Grueber et al. Citation2011) and model averaging was conducted with full average results presented (Bolker et al. Citation2009). All variables were standardised prior to analysis.
Results
Seasonal patterns in bird assemblages
Bird abundance
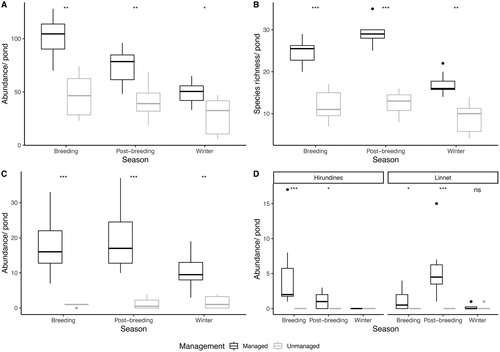
Significantly higher bird abundance was recorded at managed open-canopy pond sites over the breeding, post-breeding and winter seasons, in comparison to unmanaged overgrown ponds (). At both pond management types, bird abundance was highest over the breeding season and lowest over the winter season.
Table 1. Bird species richness and abundance for seasonal bird α-diversity by pond management type and seasonal γ-diversity of birds over all ponds.
GLMMs identified pond management as consistently exerting the strongest positive influence over bird abundance, with significant differences identified over the breeding and winter seasons ( and , online supplementary Table S2). Pond connectivity to semi-natural terrestrial landscape features such as hedgerows was also found to be a significant predictor of bird abundance, exercising a positive influence throughout the year (). Bramble area was a significant predictor of bird abundance over the breeding and post-breeding seasons, with higher bird abundance associated with larger areas of bramble. Bird abundance was significantly positively associated with overall area of the pond margin over the breeding season, and with the area of tree cover around the pond margin over the winter season.
Figure 1. Effect size plots for bird abundance over (a) breeding (b), post-breeding and (c) winter seasons. Taken from GLMM analysis, with se and upper and lower confidence intervals. Significance values as: P < 0.001 ***, P < 0.001 **, P < 0.05 *.
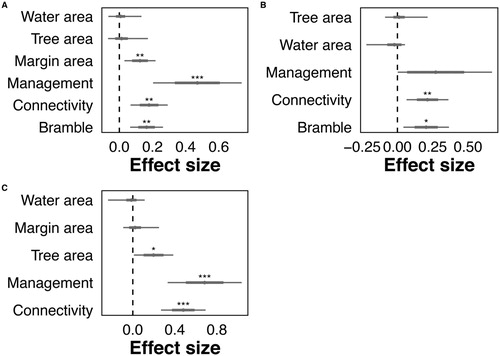
Table 2. Summary of the GLMMs explaining (a) breeding abundance and (b) species richness, in relation to pond features.
Table 3. Full average parameter estimates from the GLMMs for bird abundance, by season.
Bird species richness
Significantly higher bird species richness was observed at managed open-canopy pond sites throughout the breeding, post-breeding and winter seasons, in comparison to unmanaged overgrown ponds (). Regardless of pond management, bird species richness was highest at all study sites over the post-breeding season and lowest over the winter season.
Pond management and connectivity with terrestrial semi-natural landscape features were significant predictors exerting a positive influence on bird species richness throughout the year. Over the post-breeding season, bird species richness was also positively associated with a higher tree area around the margin ( and , online supplementary Table S2, ).
Figure 2. Effect size plots for bird species richness (a) over breeding (b), post-breeding and (b) winter seasons. Taken from GLMM analysis, with se and upper and lower confidence intervals. Significance value codes: P < 0.001 ***, P < 0.001 **, P < 0.05 *.
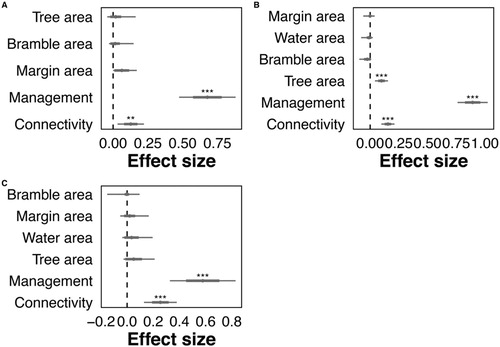
Table 4. Full average parameter estimates from the GLMMs for bird species richness, by season.
Seasonal shifts in bird use of ponds
Seasonal fluctuations in bird communities were more prominent at open-canopy pond sites in comparison to overgrown sites (online Table S3). Abundance of conservation priority red and amber listed bird species was also higher at managed-open canopy ponds over all seasons (). At managed open-canopy ponds, the abundance of obligate aerial was significantly higher over the breeding and post-breeding seasons. In contrast, no aerial insectivore visits were recorded at the unmanaged overgrown ponds over the same period. Linnet abundance was significantly higher at managed open-canopy ponds over the breeding and post-breeding season, whereas observations of this species were either very low or absent over all seasons at the overgrown ponds ().
Figure 3. (a) Bird abundance by pond management and season, (b) bird species richness by pond management and season, (c) conservation priority (red and amber) bird abundance by pond management and season and (d) abundance of aerial insectivores by pond management and season. Boxplots show median, upper and lower quartiles, with data falling outside Q1–Q3 range plotted as outliers. Statistical significance of Mann–Whitney tests denoted by P < 0.001 ***, P < 0.01 **, P < 0.05 * and ns = not significant.
Over the breeding and post-breeding seasons, managed open-canopy ponds were visited by a variety of warblers including Common Whitethroat Sylvia communis, Lesser Whitethroat Sylvia curruca, Willow Warbler Phylloscopus trochilus, Common Chiffchaff Phylloscopus collybita and Blackcap Sylvia atricapilla. With the exception of Chiffchaff, that was regularly recorded at both pond management type, other warbler species were either observed in lower numbers or entirely absent from unmanaged overgrown pond sites throughout all seasons. Other conservation priority species with specialist requirements, such as Yellowhammer Emberiza citrinella and Spotted Flycatcher Muscicapa striata were also only recorded at managed open-canopy ponds over the breeding season. In comparison, generalist bird species, such as European Robin Erithacus rubecula, Common Blackbird Turdus merula, Common Wood Pigeon Columba palumbus, Eurasian Blue Tit Cyanistes caeruleus and Eurasian Wren Troglodytes troglodytes were frequently recorded at both pond management types over all seasons. With the exception of Eurasian Blue Tit and Common Wood Pigeon, these generalist species were consistently recorded at higher frequencies at managed open-canopy ponds.
Regardless of pond management type, farmland ponds were most utilised by local bird communities over the breeding and post-breeding seasons with visits lower over winter. Nevertheless, in winter Redwing Turdus iliacus, Fieldfare Turdus pilaris, Eurasian Teal Anas crecca, Mallard Anas platyrhynchos and Common Snipe Gallinago gallinago were recorded at managed-open canopy ponds, but were largely absent from unmanaged overgrown ponds. By contrast, Eurasian Woodcock Scolopax rusticola and Brambling Fringilla montifringilla were recorded only at unmanaged overgrown pond sites in winter.
Discussion
Previous research has demonstrated the importance of farmland pond management for local bird communities (Davies et al. Citation2016, Lewis-Phillips et al. Citation2019) but seasonal patterns were not identified. To evaluate the effectiveness of farmland pond management as an AES measure, it is important to understand not only if ponds positively influence local bird communities but also whether bird community responses vary seasonally (Siriwardena et al. Citation2006, Siriwardena Citation2010). As such, the finer temporal resolution of this study offers previously unidentified insights into how local bird communities utilise managed open-canopy and unmanaged overgrown ponds in different seasons. Alongside the all-season preference for managed open-canopy ponds by more abundant and diverse bird communities, including conservation priority species, more prominent seasonal shifts in bird communities at managed open-canopy ponds likely reflect seasonal shifts in resource and habitat availability. The regular and exclusive use of managed open-canopy ponds over the breeding and post-breeding seasons by a variety of migrant warbler species (e.g. Lesser Whitethroat and Common Chiffchaff), alongside many other bird species with specialist requirements (e.g. Yellowhammer), strongly suggests an increased importance of invertebrate and seed resources at these ponds, alongside better provision of favourable (cover next to food sources) nesting sites within bramble patches and increased availability of nesting materials. Similarly, open-canopy sites are preferentially visited over winter by migrant species such as Redwing and Fieldfare, possibly due to increased availability of fruits associated with early successional bramble habitat. Further, species such as Eurasian Teal may be potentially attracted to managed open-canopy sites over winter due to accessibility to areas of open water. The less prominent seasonal shifts in bird communities at unmanaged open-canopy ponds, with visits throughout the year largely dominated by generalist non-conservation concern species, suggest that these pond sites offer a less resource-rich environment in all seasons.
Environmental controls on birds
The observed key role that pond management plays in explaining both higher bird abundances and species richness across all seasons can be linked to major management linked environmental and habitat changes that occur both within the pond and in the pond margin. The removal of pond sediment and opening of the canopy allows light to reach the water and increases the open, non-shaded water area. Consequently, macrophyte coverage and diversity increases (Sayer et al. Citation2012), providing important physical structure within the water, creating microgradients of nutrients, oxygen and pH (Declerck et al. Citation2011) and, as emergence trap results have demonstrated, a significantly increased abundance of emergent invertebrates particularly over the breeding season, in comparison to unmanaged overgrown sites (Lewis-Phillips et al. in review). Emerging aquatic invertebrates provide the foundation for numerous aquatic-terrestrial interactions, with birds directly benefitting from the simultaneous arrival of a high-quality and high-quantity food resource (Baxter et al. Citation2005, Bradbury & Kirby Citation2006, Schummer et al. Citation2012, Popova et al. Citation2017).
An abundant, pond-derived invertebrate food source may be of particular importance over the bird breeding season as the high quantities of unsaturated omega-3 fatty acids contained within aquatic invertebrates (but which are low or absent in terrestrial invertebrates), are known to be highly beneficial to chick development (Twining et al. Citation2016, Twining et al. Citation2018). Increased emergent invertebrate availability at managed, open-canopy ponds may therefore act as an incentive for some birds to visit over the breeding and post-breeding seasons in order to obtain a biochemically important, but restricted (to open-canopy aquatic habitats), food source in intensively farmed landscapes. For example, that aerial insectivores were frequently recorded at managed open-canopy ponds over the breeding season, but entirely absent from unmanaged overgrown ponds, strongly suggests that individuals were attracted to increased invertebrate foraging opportunities in the former during this season. This idea is further supported by breeding season visits to open-canopy ponds by a variety of warbler species alongside Spotted Flycatcher and Yellowhammer that were absent at the overgrown pond sites.
Restoration and management work that reduces scrub cover next to ponds tends to increase the area of wetted pond margin, particularly in the first few years after management/restoration before scrub starts to re-invade. Consequent access to wet mud at pond edges is likely a key factor determining invertebrate food availability to birds (Green et al. Citation2000), for example benefiting species that forage in soft substrates such as Common Snipe (Bradbury & Kirby Citation2006), recorded at managed open-canopy pond sites over the winter season, and Song Thrush Turdus philomelos, observed at open-canopy sites over the breeding and winter seasons. Furthermore, increases in the open water area and wet perimeter provide birds with a source of nesting material (e.g. wet mud and a variety of vegetation) alongside drinking and washing opportunities. As such, Barn Swallow and House Martin were regularly recorded drinking from open-canopy pond sites over the breeding and post-breeding seasons, while Great Tit Parus major, Yellowhammer and Wren were all observed collecting nesting materials (e.g. grasses) during the breeding season from muddy pond edges.
Larger pond margin areas were found to be significantly positively correlated with bird abundance over the breeding season, likely linked to the provision of higher levels of complex habitat thus affording more resources for bird nesting and foraging. Pond management results in habitat changes within pond margins, especially promoting the development of more diverse habitats along the moisture gradient from the pond edge to the field margin, including patches of disturbed ground, bramble, low growing scrub and seed-rich plants such as Chickweed Stellaria media. Rank wetland vegetation, frequently found around managed open-canopy ponds, is important over the breeding season for species such as Common Reed Bunting Emberiza schoeniclus that benefit from this habitat through nest concealment alongside greater invertebrate prey availability, in comparison to surrounding agricultural land (Brickle & Peach Citation2004, Redhead et al. Citation2018). Further, the positive seasonal link identified between the area of dense bramble understory area and bird abundance during the breeding and post-breeding seasons is likely related to the highly suitable nesting habitat provided by this vegetation type for a wider variety of bird species, including for species regularly recorded at managed open-canopy ponds such as Linnet (Moorcroft et al. Citation2006), Yellowhammer (Peakall Citation1960), Common Chiffchaff and Blackcap (Rodrigues & Crick Citation1997). Over the post-breeding season, species with second broods, such as Yellowhammer (Peakall Citation1960) and Chiffchaff, could continue to utilise bramble as nesting habitat, while juvenile birds may use it as cover and protection against predators. The absence of any significant relationship between bramble and bird communities over the winter season further indicates that seasonal bird usage of bramble is potentially related to nesting.
Positive associations between tree area within the margin and bird abundance over the winter season and bird species richness over the post-breeding season may be related to specific resources provided by trees. Several bird species including Common Chiffchaff and Common Chaffinch Fringilla coelebs were observed foraging in and around trees at both pond management types over the breeding and post-breeding seasons, while Fieldfare and Redwing were observed foraging for fruits around managed open-canopy ponds over the winter season. Trees generally afford shelter, food resources such as seeds and fruit (Peck Citation1989), nesting habitat and shade, and they may also increase connectivity, assisting the movement of individuals through the landscape to access pond resources. Nonetheless, despite an overall higher proportion of tree cover, bird communities recorded at unmanaged overgrown ponds were less diverse and had lower abundances, suggesting that many birds associated with farmland ponds favour tree habitats close to non-shaded open water.
The importance of increased connectivity with landscape features is well documented for a range of animal and plant species (Joyce et al. Citation1999, Bennett et al. Citation1994, Parish et al. Citation1994, Wehling & Diekmann Citation2009, Sullivan et al. Citation2017). Connectivity may be of particular importance within intensively farmed landscapes where hedgerows and other semi-natural features likely facilitate the movement of farmland bird individuals between remaining patches of good quality habitat (Hinsley & Bellamy Citation2000). The importance of connectivity specifically for birds is again well documented. Vanhinsbergh et al. (Citation2002) showed that bird species richness is significantly higher in new farm woods when they are connected by hedgerows, while Bellamy & Hinsley (Citation2005) demonstrated that farmland bird individuals are three times more likely to move through hedgerows than across open fields with the presence of a range of breeding bird species linked to improved hedgerow connectivity in woodland patches (Hinsley et al. Citation1995). In support of this previous work, our research demonstrates that increased connectivity with semi-natural landscape features such as hedgerows and woodland patches significantly influences bird communities at farmland ponds over all seasons, with ponds situated closer to hedgerows or small woodland areas hosting a higher bird abundance and species richness than more isolated ponds.
Conservation implications
Combined with the demonstrated benefits that managed open-canopy ponds offer to aquatic plants, invertebrates and amphibians (Sayer et al. Citation2012, Sayer et al. Citation2013) and building on our previous work (Davies et al. Citation2016, Lewis-Phillips et al. Citation2019), this study highlights the seasonal value of pond management and in turn in-pond and surrounding pond margin composition for localised bird communities. We show that, relative to unmanaged overgrown ponds, managed open-canopy farmland ponds are of particular importance to bird communities over the breeding and post-breeding season, as indicated by seasonal peaks in bird abundance and species diversity. In contrast, both managed open canopy and unmanaged overgrown ponds were distinctly less utilised in winter, although the former ponds still hosted a wider range of bird species, including winter migrants.
Our results have important direct implications for the design of farmland pond management in AES, showing that an approach to pond restoration and management that promotes a mosaic of open water, groups of trees, bramble, low scrub and tall grasses to develop in combination with muddy patches around the pond water edge will optimise the benefits of open ponds for local bird communities over the breeding and post-breeding seasons. Given the positive year-round influence of pond connectivity with nearby semi-natural terrestrial features such as hedgerows for birds across all seasons, we further recommend that farmland ponds situated at field edges should be prioritised for restoration and subsequent management measures and that hedgerow planting to connect isolated farmland ponds to other semi-natural features should be encouraged. Echoing previous work (Sayer et al. Citation2013, Davies et al. Citation2016, Lewis-Phillips et al. Citation2019), we strongly conclude that farmland ponds have been undervalued in farmland conservation for a range of taxa and advocate that this situation should be re-evaluated so that pond management is more widely adopted and promoted within future AES.
Acknowledgements
We would like to thank the London Doctoral Training Partnership and the Wildfowl & Wetlands Trust for their continued support throughout this project. We also thank Derek Sayer, Jimmy Gallon, Thomas and Amelia Courthauld, Peter Seaman and Paul Marsh for site access. We dedicate this paper to Richard Waddingham who inspired this study.
ORCID
Jonathan Lewis-Phillips http://orcid.org/0000-0002-3070-8531
Additional information
Funding
References
- Aebischer, N.J., Bailey, C.M., Gibbons, D.W., Morris, A.J., Peach, W.J. & Stoate, C. 2015. Twenty years of local farmland bird conservation: the effects of management on avian abundance at two UK demonstration sites. Bird Study 63: 1–21.
- Bates, D. & Maechler, M. 2010. lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-37. https://cran.r-project.org/package=lme4.
- Baxter, C.V., Fausch, K.D. & Saunders, W.C. 2005. Tangled webs: Reciprocal flows of invertebrate prey link streams and riparian zones. Freshwater Biol. 50: 201–220. doi: 10.1111/j.1365-2427.2004.01328.x
- Bellamy, P.E. & Hinsley, S.A. 2005. The role of hedgerows in linking woodland bird populations. In McColin D, Jackson J (eds) Planning, People and Practice: The Landscape Ecology of Sustainable Landscapes. Proceedings of the thirteenth IALE (UK) conference.
- Bennett, A.F., Henein, K. & Merriam, G. 1994. Corridor use and the elements of corridor quality: Chipmunks and fencerows in a farmland mosaic. Biol. Conserv. 68: 155–165. doi: 10.1016/0006-3207(94)90347-6
- Bibby, C.J., Burgress, N.D., Hill, D. & Mustoe, S. 1992. Bird Census Techniques. Academic Press, Cambridge.
- Bolker, B.M., Brooks, M.E., Clark, C.J. & Geange, S.W. 2009. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 24: 127–135. doi: 10.1016/j.tree.2008.10.008
- Booth, G.D., Niccolucci, M.J., Schuster, E.G., & Ogden, U. 1994. Identifying Proxy Sets in Multiple Linear Regression: An Aid to Better Coefficient Interpretation. U.S. Department of Agriculture, Forest Service, Intermountain Research Station (Research paper INT).
- Bradbury, R.B. & Kirby, W.B. 2006. Farmland birds and resource protection in the UK: Cross-cutting solutions for multi-functional farming? Biol. Conserv. 129: 530–542. doi: 10.1016/j.biocon.2005.11.020
- Brickle, N.W. & Peach, W.J. 2004. The breeding ecology of Reed Buntings. Ibis 146: 69–77. doi: 10.1111/j.1474-919X.2004.00349.x
- Bright, J., Morris, T. & Winspear, R. 2008. A review of indirect effects of pesticides on birds and mitigating land-management practices. Research Report, RSPB, UK.
- BTO. 2011. Breeding Bird Survey. Research Report, BTO, Thetford.
- Burnham, D.R. & Anderson, K. 2002. Model Selection and Multimodel Inference. Springer, New York.
- Butler, S.J., Boccaccio, L., Gregory, R.D., Vorisek, P. & Norris, K. 2010. Quantifying the impact of land-use change to European farmland bird populations. Agric. Ecosyst. Environ. 137: 348–357. doi: 10.1016/j.agee.2010.03.005
- Crist, T.O., Veech, J.A., Gering, J.C. & Summerville, K.S. 2003. Partitioning species diversity across landscapes and regions: A hierarchical analysis of α, β, and γ diversity. Am. Nat 162: 734–743. doi: 10.1086/378901
- Davey, C., Vickery, J., Boatman, N., Chamberlain, D., Parry, H.R. & Siriwardena, G.M. 2010. Assessing the impact of Environmental Stewardship on lowland farmland birds in England. Ibis 152: 459–474. doi: 10.1111/j.1474-919X.2009.01001.x
- Davies, S.R., Sayer, C.D., Greaves, H., Siriwardena, G.M. & Axmacher, J.C. 2016. A new role for pond management in farmland bird conservation. Agric. Ecosyst. Environ. 233: 179–191. doi: 10.1016/j.agee.2016.09.005
- Declerck, S.A.J., Bakker, E.S., van Lith, B., Kersbergen, A. & van Donk, E. 2011. Effects of nutrient additions and macrophyte composition on invertebrate community assembly and diversity in experimental ponds. Basic Appl. Ecol. 12: 466–475. doi: 10.1016/j.baae.2011.05.001
- Defra. 2018. Wild bird populations in the UK, 1970–2015. Research Report, Defra, UK.
- Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carre, G., Garcia Marquez, J.R., Gruber, B., Lafourcade, P.J., Leitao, P.J., Munkemuller, T., McClean, C., Osborn, P.E., Reineking, B., Schroder, B., Skidmore, A.K., Zurell, D. & Lautenbach, S. 2013. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 36: 027–046. doi: 10.1111/j.1600-0587.2012.07348.x
- Evans, A., Vickery, J. & Shrubb, M. 2004. Importance of overwintered stubble for farmland bird recovery : A reply to Potts. Without this prescription, populations of seed-eating passerines are unlikely to recover. Bird Study 51: 94–96. doi: 10.1080/00063650409461339
- Green, R.E., Tyler, G.A. & Bowden, C.G.R. 2000. Habitat selection, ranging behaviour and diet of the stone curlew (Burhinus oedicnemus) in southern England. J. Zool. 250: 161–183. doi: 10.1111/j.1469-7998.2000.tb01067.x
- Grueber, C.E., Nakagawa, S., Laws, R.J. & Jamieson, I.G. 2011. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 24: 699–711. doi: 10.1111/j.1420-9101.2010.02210.x
- Hallmann, C.A., Foppen, R.P.B. & van Turnhout, C.A.M. 2014. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 511: 341–343. doi: 10.1038/nature13531
- Hinsley, S. & Bellamy, P.E. 2000. The influence of hedge structure, management and landscape context on the value of hedgerows to birds: A review. J. Environ. Manage. 60: 33–49. doi: 10.1006/jema.2000.0360
- Hinsley, S.A., Bellamy, P.E., Newton, I. & Sparks, T.H. 1995. Habitat and landscape factors influencing the presence of individual breeding bird species in woodland fragments habitat and landscape factors influencing the presence of individual breeding bird species in woodland fragments. J. Avian Biol. 26: 94–104. doi: 10.2307/3677057
- Inger, R., Gregory, R., Duffy, J.P., Stott, I., Vorısek, P. & Gaston, K.J. 2015. Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol. Lett. 18: 28–36. doi: 10.1111/ele.12387
- Jeffs, C., Davies, M., Carter, I., Gregson, J., Sainsbury, A. & Lister, J. 2016. Reintroducing the Cirl Bunting to Cornwall. Br. Birds 109: 374–388.
- Jost, L. 2006. Entropy and diversity. Oikos 113: 363–375. doi: 10.1111/j.2006.0030-1299.14714.x
- Joyce, K.A., Holland, J.M. & Doncaster, C.P. 1999. Influences of hedgerow intersections and gaps on the movement of carabid beetles. Bull. Entomol. Res. 89: 523–531. doi: 10.1017/S000748539900067X
- Lewis-Phillips, J., Brooks, S., Sayer, C.D., McCrea, R., Siriwardena, G.M. & Axmacher, J.C. 2019. Pond management enhances the local abundance and species richness of farmland bird communities. Agric. Ecosyst. Environ 273: 130–140. doi: 10.1016/j.agee.2018.12.015
- Moorcroft, D., Wilson, J. & Bradbury, R. 2006. Diet of nestling Linnets Carduelis cannabina on lowland farmland before and after agricultural intensification. Bird Study 53: 156–162. doi: 10.1080/00063650609461428
- Natural England. 2009. Agri-environment schemes in England 2009. Research Report, Defra, UK.
- Newton, I. 2004. The recent declines of farmland bird populations in Britain: An appraisal of causal factors and conservation actions. Ibis 146: 579–600. doi: 10.1111/j.1474-919X.2004.00375.x
- Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P., O'Hara, R.B., Simpson, G., Solymos, P., Stevens, M.H.H. & Wagner, H. 2013. Vegan: Community Ecology Package. R Package Version. 2.0-10. https://cran.r-project.org/package=vegan.
- Parish, T., Lakhani, K.H. & Sparks, T.H. 1994. Modelling the relationship between bird population variables and hedgerow and other field margin attributes. I. Species richness of summer, winter and breeding birds. J. Appl. Ecol. 31: 764–775. doi: 10.2307/2404166
- Peakall, D.B. 1960. Nest records of the yellowhammer. Bird Study 7: 94–102. doi: 10.1080/00063656009475963
- Peck, K.M. 1989. Tree species preferences shown by foraging birds in forest plantation in northern England. Biol. Conserv. 48: 41–57. doi: 10.1016/0006-3207(89)90058-X
- Popova, O.N., Haritonov, A.Y., Sushchik, N.N., Makhutova, O.N., Kalachova, G.S., Kolmakova, A.A. & Gladyshev, M.I. 2017. Export of aquatic productivity, including highly unsaturated fatty acids, to terrestrial ecosystems via Odonata. Sci. Total Environ. 581–582: 40–48. doi: 10.1016/j.scitotenv.2017.01.017
- Ralph, C.J., Sauer, J.R., & Droege, S. (eds) 1995. Monitoring bird populations by point counts. Gen. Tech. Rep. PSW-GTR-149. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station. doi: 10.2737/PSW-GTR-149
- R Core Team. 2017. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.r-project.org.
- Redhead, J.W., Hinsley, S.A., Beckmann, B.C., Broughton, R.K. & Pywell, R.F. 2018. Effects of agri-environmental habitat provision on winter and breeding season abundance of farmland birds. Agric. Ecosyst. Environ. 251: 114–123. doi: 10.1016/j.agee.2017.09.027
- Robinson, R.A. & Sutherland, W.J. 2002. Post-war changes in arable farming and biodiversity in Great Britain. J. Appl. Ecol. 39: 157–176. doi: 10.1046/j.1365-2664.2002.00695.x
- Rodrigues, M. & Crick, H.Q.P. 1997. The breeding biology of the Chiffchaff Phylloscopus collybita in Britain: A comparison of an intensive study with records of the BTO Nest Record Scheme. Bird Study 44: 374–383. doi: 10.1080/00063659709461073
- RSPB. 2018. UK conservation status explained. Available at: https://www.rspb.org.uk/birds-and-wildlife/wildlife-guides/uk-conservation-status-explained/.
- Sayer, C.D. 2014. Conservation of aquatic landscapes: Ponds, lakes, and rivers as integrated systems. Wires Water 1: 573–585. doi: 10.1002/wat2.1045
- Sayer, C., Andrews, K., Shilland, E., Edmonds, N., Edmonds-Brown, R., Patmore, I., Emson, D. & Axmacher, J.C. 2012. The role of pond management for biodiversity conservation in an agricultural landscape. Aquat. Conserv. 22: 626–638. doi: 10.1002/aqc.2254
- Sayer, C.D., Shilland, E., Greaves, H., Dawson, B., Patmore, I.R., Emson, E., Alderton, E., Robinson, P., Andrews, K., Axmacher, J.A. & Wiik, E. 2013. Managing British ponds—conservation lessons from a Norfolk farm. Br. Wildl. 25: 21–28.
- Schummer, M.L., Palframan, J., McNaughton, E., Barney, T. & Petrie, S. 2012. Comparisons of bird, aquatic macroinvertebrate, and plant communities among dredged ponds and natural wetland habitats at Long Point, Lake Erie, Ontario. Wetlands 32: 945–953. doi: 10.1007/s13157-012-0328-2
- Siriwardena, G.M. 2010. The importance of spatial and temporal scale for agri-environment scheme delivery. Ibis 152: 515–529. doi: 10.1111/j.1474-919X.2010.01034.x
- Siriwardena, G.M., Calbrade, N.A., Vickery, J.A. & Sutherland, W.J. 2006. The effect of the spatial distribution of winter seed food resources on their use by farmland birds. J. Appl. Ecol. 43: 628–639. doi: 10.1111/j.1365-2664.2006.01170.x
- Sullivan, M.J.P., Pearce-Higgins, J.W., Newson, S.E., Scholefield, P., Brereton, T. & Oliver, T.H. 2017. A national-scale model of linear features improves predictions of farmland biodiversity. J. Appl. Ecol. 54: 1776–1784. doi: 10.1111/1365-2664.12912
- Thornhill, I. 2017. The application of graph theory and percolation analysis for assessing change in the spatial configuration of pond networks. Urban. Ecosyst. 21: 213–225.
- Tucker, G. & Heath, M. 1994. Birds in Europe: Their Conservation Status. Birdlife International, Cambridge.
- Twining, C.W., Brenna, J.T., Lawrence, P., Shipley, J.R., Tollefson, T.N. & Winkler, D.W. 2016. Omega-3 long-chain polyunsaturated fatty acids support aerial insectivore performance more than food quantity. PNAS 113: 10920–10925. doi: 10.1073/pnas.1603998113
- Twining, C.W., Shipley, J.R. & Winkler, D.W. 2018. Aquatic insects rich in omega-3 fatty acids drive breeding success in a widespread bird. Ecol. Lett. 21: 1812–1820. doi: 10.1111/ele.13156
- Vanhinsbergh, D., Gough, S., Fuller, R.J. & Brierley, E.D.R. 2002. Summer and winter bird communities in recently established farm woodlands in lowland England. Agric. Ecosyst. Environ. 92: 123–136. doi: 10.1016/S0167-8809(01)00301-2
- Vickery, J.A., Feber, R.E. & Fuller, R.J. 2009. Arable field margins managed for biodiversity conservation: A review of food resource provision for farmland birds. Agric. Ecosyst. Environ. 133: 1–13. doi: 10.1016/j.agee.2009.05.012
- Voříšek, P., Klvaňová, A., Wotton, S. & Gregory, R.D. 2008. A Best Practice Guide for Wild Bird Monitoring Schemes. RSPB, London.
- Wehling, S. & Diekmann, M. 2009. Importance of hedgerows as habitat corridors for forest plants in agricultural landscapes. Biol. Conserv. 142: 2522–2530. doi: 10.1016/j.biocon.2009.05.023
- Wilson, J.D., Morris, A.J., Arroyo, B.E. & Clark, S.C. 1999. A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in Northern Europe in relation to agricultural change. Agric. Ecosyst. Environ. 75: 13–30. doi: 10.1016/S0167-8809(99)00064-X