
ABSTRACT
Capsule: The undulating display flight of Golden Eagles Aquila chrysaetos has a territorial function on current consensus: we found that displays could occur at any location within a home range of territorial birds and were not more frequent at the territorial limits (as documented by a previous study) or around the nest site.
Aims: To test the null hypothesis that display activity does not spatially differ within the home range of territorial Golden Eagles.
Methods: We used 1488 h of observations of Golden Eagles between April 2003 and October 2004 (excluding November through January inclusive) on the Isle of Lewis, northwest Scotland involving primarily four territories. Territorial boundaries and nest sites were determined by the behaviour of the focal territorial birds. All flight activity was cast into three spatial categories (near-boundary, near-nest and elsewhere) and the relative occurrence of displays was referenced by overall flight activity recorded in these spatial categories.
Results: We failed to reject the null hypothesis. Prior findings of greater display rate at territory limits were not confirmed. Territorial Golden Eagles apparently displayed wherever they happened to be in their home range. There was no difference in display frequency between successful and failed breeding pairs and, within pairs, between successful and failed breeding seasons. Monthly changes in display activity did not differ from monthly changes in the presence of ‘intruding’ sub-adults. A peak in non-territorial bird activity in August and September was not reflected by a peak in display, however, we could not distinguish recently fledged ‘non-threatening’ birds in this period.
Conclusions: Undulating flight displays of Golden Eagles focused neither on territory boundaries nor the nest. Displays may be spontaneous or occur in response to the detection of potential trespassers or neighbours, however, we could not rule out any role of pair-bonding.
Large raptors are uncommon, range widely, and consequently their behaviour is difficult to observe (Newton Citation1979). Recording and interpreting the functions of their behaviour is thereby often fraught through limited data. Moreover, compared to several other birds, large raptors are usually not known for ostentatious display behaviour (Cramp & Simmons Citation1980).
Nevertheless, a distinctive display of Golden Eagles Aquila chrysaetos is an undulating flight in which birds perform a series of deep, alternating dives (stoops) and climbs. Although variations occur, simply described the display is initiated by a steep stoop with wings partially folded, followed by an upward swoop with wings flapping at the apex of the climb (for a full description see: Cramp & Simmons Citation1980, Bergo Citation1987, McGrady Citation1997, Watson Citation2010). These displays were first interpreted as having a nuptial purpose (e.g. mate attraction or ‘pair-bonding’; Brown & Amadon Citation1968). Contradicting this previous interpretation and following new evidence, the undulating flight was later thought to serve primarily as territorial signaling and defence (Harmata Citation1982, Collopy & Edwards Citation1989, Marzluff et al. Citation1997). As part of this thesis advocating a territorial function of undulating displays, Collopy & Edwards (Citation1989) noted that: ‘Most of the observations [of displays] occurred at or near territory boundaries’ during their study of four territorial pairs in Idaho, USA. Marzluff et al. (Citation1997) stated that: ‘Undulating flights were most often given at the edge of territories, rather than near nests’ in later research on more birds in Idaho, additionally noting that displays were least frequent in newly formed pairs.
In our study, Golden Eagle flights were observed on the Isle of Lewis in the Western Isles of Scotland to examine the spatial distribution of undulating flight displays by territory holders and to test the hypothesis that display frequency does not vary in different parts of the home range. An alternative prediction was that of greater display frequency close to territory boundaries (Collopy & Edwards Citation1989, Marzluff et al. Citation1997). We also examined the timing of displays by territory holders in relation to the intrusion of presumed non-territorial individuals and contrasted display activity among pairs of different reproductive status.
Methods
The study site was a 290 km2 area of south Lewis, the northernmost island of the Western Isles of northwest Scotland. The landscape is characteristically low rocky hills (maximum elevation 570 m above sea level) interspersed with lakes (lochs), streams and small rivers, with narrow sea lochs extending into the site at several points. Our study originated to describe the home range use of Golden Eagles in the vicinity of a proposed wind farm as part of the assessment of the proposal’s potential impact.
Golden Eagles in Scotland are non-migratory, and breeders occupy their home ranges year-round (Watson Citation2010). In most parts of Scotland, including the study area, territory holding Golden Eagles do not share extensive foraging areas with their neighbours. We found eight Golden Eagle home ranges within the study area: observations concentrated on four core home ranges that were relatively centrally located. Each home range included many hills and slopes where uplift could be generated by winds that could be used by birds when moving around their home ranges (McLeod et al. Citation2002, Fielding et al. Citation2003, Citation2019). No birds were individually identifiable, although throughout the study all identified territorial pairs were in adult plumage (Bloom & Clark Citation2001). Breeding Golden Eagles are territorial and central place foragers (Watson Citation2010) and so we could identify individuals as occupants of particular home ranges from the context of their behaviours during the time they were in view (e.g. the direction of flights or where they perched). Although the male and female of a pair could often be determined because of their size difference, this determination was not always certain, and this precluded analyses of data by gender.
Observations of eagles were made by a single observer (RR) during timed sessions between April 2003 and October 2004 (excluding November through January inclusive) from 13 vantage points within the study area. Vantage points were chosen collectively to provide the best overview of the proposed wind farm footprint and surrounding 2 km buffers and minimize potential effects or bias due to eagles avoiding observers. The vantage point locations were also chosen to minimize potential disturbance of nesting birds, and so they were relatively distant from nest sites, while still providing full visual coverage of the area around the nest. Each observation session typically lasted 3 h, unless curtailed by poor weather, with two sessions per day. Observations were conducted from each vantage point over two sessions per month. Within that constraint the selection of the vantage point to use at the beginning of any observation day was random, and for practical reasons when two observation sessions occurred in a day the second vantage point was the one nearest to the first. Observations were made by an observer experienced in the methodology (which is typically used at any potential wind farm site in Scotland where an adverse effect on eagles may be a concern), and trained in using visual cues to accurately estimate location and elevation of eagles (Scottish Natural Heritage Citation2014). The topography within the home range is rugged, containing mountains, hillocks and ridges. The observer visually estimated all flight paths of Golden Eagles and locations of undulating display flights to a distance of 2 km from the vantage point on 1:25,000 topographic maps, using the many terrain features to aid in the accuracy of mapping. We recorded the duration of each flight, whether a display or not, from when the eagle was first detected to when it disappeared from view or perched.
A nest was a stick structure where eagles laid eggs in the year of study or that contained fresh nesting material in the absence of egg-laying, and successful pairs were those that raised nestlings to fledging age (Hardey et al. Citation2013, Hayhow et al. Citation2017, Steenhof et al. Citation2017; Gjershaug et al. Citation2018). A nest was deemed to be ‘active’ (‘occupied’: Steenhof et al. Citation2017) in the year of study according to the appropriate temporal presence of fresh nesting material (indicating recent nest-building by observed pairs of birds) as a minimum sign of territory occupancy; and any further activities towards reproduction (e.g. egg laying as the initial further activity). A ‘nest site’ was the geographical location of such a nest in the year of study. Breeding success (young fledged) or failure at each active nest in each year was determined (Hardey et al. Citation2013). Failure in the study (involving eight pairs across two years) typically involved either not laying eggs or at the incubation phase (at young chick stage in one pair in one year) and success involved no more than one fledgling (two pairs in 2004).
Home range boundaries were estimated by taking the outermost geographical limits of flights when the observer was certain that a particular member of a pair (or the pair) was involved i.e. where a flight originated or ended within an area of 500 m around the nest of that pair. In concordance with Collopy & Edwards (Citation1989) these limits were typically where the observer also noted interactions between neighbouring birds. We noted limited overlap in boundaries of home ranges of neighbouring pairs in some areas (see also Marzluff et al. Citation1997, Watson et al. Citation2014), and in such cases we considered the boundary to be the mid-points within the area of overlap. Home range boundaries and nest sites were digitized and entered as layers in a Geographical Information System (GIS).
All eagle flight paths and display flight start and finish locations were digitized and entered as layers in a GIS. We plotted the location of each display flight by territory holding adults at the mid-point between each start and finish location. Display activity was defined as the number of displays per 100 km of flight. We estimated the relative use of different areas by calculating the length of flight per unit area within the GIS. Our basic area units were 200 × 200 m pixel grid squares, chosen as an appropriate spatial scale to incorporate the likely error limits in recording flight path locations (McLeod et al. Citation2002). Observation effort was not incorporated into analyses of eagle use for each grid square or component area of territory because analyses relied on contrasting frequency of display against the frequency of all flight activity and hence any area differences in observation effort were common to both parameters.
To analyse differences in distribution of eagle display activity of territory holders within a home range we calculated the frequency of eagle display activity that occurred within three spatial categories: (1) 500 m of a home range boundary, (2) 500 m of the nest site, and (3) elsewhere within the home range. We also calculated the frequency of display flights in these categories. Our null hypothesis was that display activity should not differ between these spatial categories according to overall flight activity. However, we predicted alternatively that distribution of display flights should be disproportionately greater in the first spatial category (close to the territory boundary) when contrasted with the distribution of all flight activity, based on Collopy & Edwards (Citation1989) and Marzluff et al. (Citation1997). We used Chi-square tests with Yates’ correction to contrast observed and expected frequencies of display activity with respect to all flight activity (km of flight lines). A Kolmogorov–Smirnov test determined if monthly distributions of displays by territorial birds and floater intrusions differed significantly (P < 0.05).
Results
Golden Eagles were observed for 1488 h. Display frequency peaked in March, and declined thereafter (). Frequency of display flights was greater in the breeding season (March to August) than during the rest of the year (χ2 = 21.62, P < 0.001). All four core pairs failed to rear young in 2003 and two pairs fledged one young each in 2004. For the two pairs which fledged young in 2004 there was no difference in display frequency from April to August between 2003 and 2004 (χ2 = 2.41, P = 0.12; χ2 = 0.00, P = 1.00). There was also no difference in display frequency between two successful pairs and the two failed pairs, from March to August 2004 (χ2 = 2.32, P = 0.13). The most obvious annual difference was in one pair which was known to have established their territory in 2003 (where none existed previously), and display activity was higher (χ2 = 4.06, P = 0.04) in 2003 than in 2004: 13.5 and 4.0, respectively (displays per 100 km of flight).
Figure 1. Frequency of Golden Eagle undulating display flights observed by month on Isle of Lewis, Scotland: (a) number of displays per 100 km of flight path, 2004 only; and (b) number of displays per hour of observation, 2003 and 2004 (number of hours observation above columns).
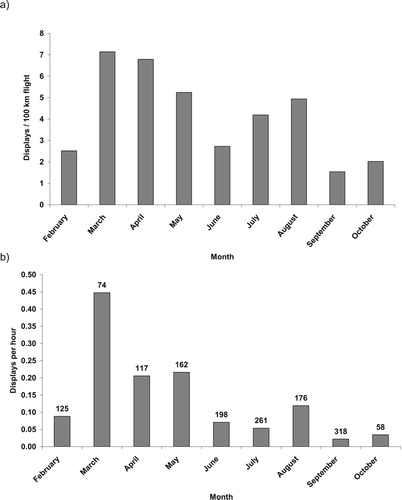
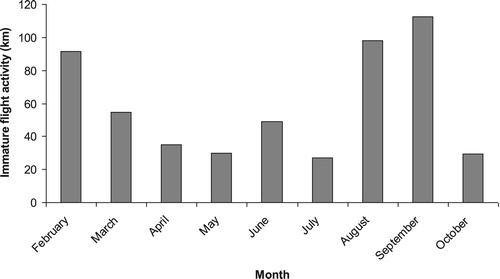
Intrusions of non-territorial eagles (juveniles and sub-adults) within four core territories were relatively high in February, declining in subsequent months before rising again in August and September (). There was no difference among monthly distribution of displays by territorial birds and monthly distribution of flight activity (km of flight lines within home range) of intruding juveniles and sub-adults within territories (Kolmogorov–Smirnov Test, Z = 0.70, P = 0.70).
In four territories where observations were concentrated, there was no evidence that displays were more likely within 500 m of boundaries (χ2 = 0.26, P = 0.61) or within 500 m of nest sites (χ2 = 0.51, P = 0.48) compared with other parts of the territory (). Therefore, undulating displays tended to occur in different parts of the territory at a similar frequency, so that there was no propensity for Golden Eagles to display more often close to an active nest or near territory boundaries. One pair had probably occupied a territory before the study period and later experienced a failure (2003) and a success (2004) in breeding. Perhaps counter-intuitively on activity through breeding status (Haworth et al. Citation2006) in this pair there was greater likelihood of displays (per 100 km of flight) close to the nest in 2003 than in 2004 (15.1 and 1, respectively): (χ2 = 10.69, P < 0.01) with little annual difference at the territory boundaries (6.1 and 5.5, respectively).
Table 1. Distribution of all Golden Eagle flight activity and undulating displays within three spatial categories for the four territories where most observation effort was concentrated.
Discussion
Our results supported the null hypothesis that the frequency of Golden Eagle undulating flight displays did not differ spatially within the home range. We consequently found that displays were not more likely to occur close to nest sites or at the home range limits. On the latter, we did not concur with the results of Collopy & Edwards (Citation1989) and Marzluff et al. (Citation1997).
Although we found no difference between monthly distributions of displays and monthly distributions of floater (non-territorial birds) intrusions, there were some notable differences in several months. Our observed monthly frequency of displays was similar to that documented by Bergo (Citation1987) and Watson (Citation2010) although our recorded frequency in February (the month preceding first prospective egg-laying) was surprisingly low based on expectations from these studies. The data reported by Watson (Citation2010), albeit over a 15-year period, should be relatively comparable to ours because they were collected in western Scotland. In Watson’s (Citation2010) study area, monthly frequency of display was highest in January, a month for which we had no data, but Watson (Citation2010) also noted a broader annual peak extending from January into March. As noted previously, we recorded low display activity in February; but we found a high display activity in the following two months. Factors other than intrusion rate, including age of the territory holders and local densities of breeders and their breeding schedules, may affect frequency of displays.
Many displays are apparently prompted by intruders according to Harmata (Citation1982) and Bergo (Citation1987). Our index of such intrusions showed relatively high flight activity of non-territorial birds in February, which would coincide with the timing of displays observed by Bergo (Citation1987) and Watson (Citation2010) but was not matched by February display activity in our study. Our recorded peak of ‘non-territorial birds’ in August and September was also not reflected by a response through high display activity by territorial eagles. At this time of year many of these ‘intrusions’ were probably newly fledged juveniles, likely not perceived by territory holders as threats (Gjershaug et al. Citation2019) through plumage (Bloom & Clark Citation2001). In contrast, as field observations were at distance, we could not distinguish between newly fledged juveniles or older sub-adult birds, and we did not know if recently fledged birds were from the focal territory or other neighbours.
In essence, we found that displays occurred wherever an eagle happened to be in its territory, likely when stimulated to perform the behaviour. We alternatively expected that undulating displays would be performed more often close to the boundaries of a territory (Collopy & Edwards Citation1989, Marzluff et al. Citation1997). Our study did not concur with this prior finding, but this does not necessarily dispute the central thesis of Collopy & Edwards (Citation1989) on a territorial function. Territorial defence might principally be directed at neighbouring pairs but according to Harmata (Citation1982) and Bergo (Citation1987), intruding sub-adults are often also responsible for eliciting undulating displays. More recent data from satellite tagged Golden Eagles in Scotland show that sub-adults may frequently intrude into occupied territories (Weston Citation2014). Our seasonal records of non-territorial birds as likely intruders (being lowest in the breeding season: ) broadly coincided with the seasonal pattern of intrusion behaviour by satellite tagged sub-adults elsewhere in Scotland (Weston Citation2014). Our study had no data from November through January inclusive. Given the broad coincidence with Weston (Citation2014) in other months, however, intrusions would probably have been high in these undocumented winter months also.
Figure 2. Flight activity (km of flight paths) of non-territorial juvenile and sub-adult Golden Eagles within four core territories.
If eagles also display towards neighbouring pairs when they detect a displaying neighbour (Harmata Citation1982) then this would also not necessarily be a behaviour which would be restricted to the territory boundary; rather it would be a display performed at a location where the displaying neighbour was seen. Therefore, even though displays were not more likely to occur near territory boundaries in our study, our results are still consistent with the hypothesis that undulating flight displays serve primarily as a signal of territory occupancy (Harmata Citation1982, Bergo Citation1987, Collopy & Edwards Citation1989, Marzluff et al. Citation1997). Alternatively, however, our data are inadequate to rule out entirely any potential role of undulating displays in ‘nuptial’ activities such as pair-bonding (Brown & Amadon Citation1968). Our finding that undulating displays were not more frequent at territory boundaries was despite previous findings which equated spatial prevalence at territory boundaries as consistent with a territorial function. Haworth et al. (Citation2006) recorded that failed/non-breeding birds spent more time away from the nest site and towards the territorial limits. Yet, in an established pair which failed at an early stage to reproduce in 2003 but produced a fledgling in 2004, we found no greater propensity for display at the outer limits in 2003. Moreover, our observations of greater display activity in a newly established pair (and territory) in the year of establishment (cf Marzluff et al. Citation1997) could be because the birds were establishing a new territory (greater need for ‘territory defence’) or that the birds were probably a new pair (greater propensity to ‘pair-bond’). Dual functionality of other behaviours and plumage-signals of Golden Eagles have been recently proposed (Walker Citation2017, Gjershaug et al. Citation2019) and undulating displays may also have a dual function.
Acknowledgements
We are grateful for assistance from Mike McGrady, Carol McIntyre, Michael Collopy, Jan Ove Gjershaug, Michael Kochert, Julien Terraube, Ruth Tingay, and an anonymous referee in improving earlier drafts of the manuscript. David McLeod was extremely helpful in data extraction. Paul Haworth instigated, directed and gained funds for this study of Golden Eagles in Lewis, but sadly died before he could appreciate this product of his many efforts.
ORCID
D. Philip Whitfield http://orcid.org/0000-0003-4255-7782
Additional information
Funding
References
- Bergo, G. 1987. Territorial behaviour of golden eagles in western Norway. Brit. Birds 80: 361–376.
- Bloom, P.H. & Clark, W.S. 2001. Molt and sequence of plumages of golden eagles and a technique for in-hand ageing. N. Am. Bird Bander 26: 97–116.
- Brown, L.H. & Amadon, D. 1968. Eagles, Hawks and Falcons of the World. McGraw-Hill, New York.
- Collopy, M.W. & Edwards, T.C. 1989. Territory size, activity budget, and the role of undulating flight in nesting golden eagles. J. Field Ornith. 60: 43–51.
- Cramp, S. & Simmons, K.E.L. 1980. The Birds of the Western Palearctic. Vol. II. Oxford University Press, Oxford.
- Fielding, A., Whitfield, D.P., McLeod, D.R.A., McGrady, M.J. & Haworth, P.F. 2003. Modelling the impact of land use change on Golden Eagles (Aquila chrysaetos). In Thompson, D.B.A., Redpath, S.M., Fielding, A.H., Marquiss, M. & Galbraith, C.A. (eds) Birds of Prey in a Changing Environment, 323–340. The Stationery Office, Edinburgh.
- Fielding, A.H., Haworth, P.F., Anderson, D., Benn, S., Dennis, R., Weston, E. & Whitfield, D.P. 2019. A simple topographical model to predict Golden Eagle Aquila chrysaetos space use during dispersal. Ibis. DOI:10.1111/ibi.12718.
- Gjershaug, J.O., Brøseth, H., Kleven, O., Kålås, J.-A., Mattisson, J. & Tovmo, M. 2018. Monitoring methods for the Golden Eagle Aquila chrysaetos in Norway. Bird Study 65: S43–S51. doi: 10.1080/00063657.2018.1478389
- Gjershaug, J.O., Halley, D. & Stokke, B.G. 2019. Predefinitive plumage in the Golden Eagle (Aquila chrysaetos): a signal of aggression or submission? J. Raptor Res. 53: 431–435. doi: 10.3356/0892-1016-53.4.431
- Hardey, J., Crick, H.Q.P., Wernham, C.V., Riley, H., Etheridge, B. & Thompson, D.B.A. 2013. Raptors. A Field Guide for Surveys and Monitoring, 3rd edn. The Stationery Office, Edinburgh.
- Harmata, A.R. 1982. What is the function of undulating flight display in golden eagles? Raptor Res. 16: 103–109.
- Haworth, P.F., McGrady, M.J., Whitfield, D.P., Fielding, A.H. & McLeod, D.R.A. 2006. Ranging behaviour of golden eagles Aquila chrysaetos in western Scotland according to season and breeding status. Bird Study 53: 265–273. doi: 10.1080/00063650609461442
- Hayhow, D.B., Benn, S., Stevenson, A., Stirling-Aird, P.K. & Eaton, M.A. 2017. Status of Golden Eagle Aquila chrysaetos in Britain in 2015. Bird Study 64: 281–294. doi: 10.1080/00063657.2017.1366972
- Marzluff, J.M., Knick, S.T., Vekasy, M.S., Scheuck, L.S. & Zarriello, T.J. 1997. Spatial use and habitat selection of Golden Eagles in southwest Idaho. Auk 114: 673–687. doi: 10.2307/4089287
- McGrady, M.J. 1997. Aquila chrysaetos Golden Eagle. BWP Update 1: 99–114.
- McLeod, D.R.A., Whitfield, D.P., Fielding, A.H., Haworth, P.F. & McGrady, M.J. 2002. Predicting home range use by Golden Eagles Aquila chrysaetos in western Scotland. Avian Sci. 2: 183–198.
- Newton, I. 1979. Population Ecology of Raptors. Poyser, Berkhamstead.
- Scottish Natural Heritage. 2014. Guidance. Recommended Bird Survey Methods to Inform Impact Assessment of Onshore Wind Farms. May 2014. SNH, Battleby.
- Steenhof, K., Kochert, M.N., McIntyre, C.L. & Brown, J.L. 2017. Coming to terms about describing golden eagle reproduction. J. Raptor Res. 51: 378–390. doi: 10.3356/JRR-16-46.1
- Walker, D. 2017. A Fieldworker’s Guide to the Golden Eagle. Whittles Publishing, Dunbeath.
- Watson, J. 2010. The Golden Eagle, 2nd edn. Poyser, London.
- Watson, J.E., Duff, A.A. & Davies, R.W. 2014. Home range and resource selection by GPS-monitored adult Golden Eagles in the Columbia plateau ecoregion: implications for wind power development. J. Wildl. Manage. 78: 1012–1021. doi: 10.1002/jwmg.745
- Weston, E. 2014. Juvenile dispersal behaviour in the Golden Eagle (Aquila chrysaetos). PhD Thesis, University of Aberdeen.