
ABSTRACT
Capsule: Within breeding season changes in nest orientation of the Rufous Hornero Furnarius rufus may reflect responses to the local climate.
Aims: To explore the effect of climate on the orientation of closed-cup nests of the Rufous Hornero at a subtropical urban site throughout its breeding period.
Methods: The breeding season was divided into two periods: a cold period with low rainfall and a warm, humid period. For each period, the mean nest entrance orientation and distribution were estimated and compared against a random orientation model.
Results: A total of 42 nests were found during the first period of reproduction and 32 during the second period. In both cases, the mean orientation was non-random and significantly different between the two periods: nests were more likely to be orientated towards the northwest in the first period and to the east in the second period.
Conclusions: Climate variation throughout the breeding season was reflected in differences in nest entrance orientation. This could be because this species takes advantage of solar radiation to increase the temperature within the nest during the first cold months of the breeding season and reduces the humidity of the nest, generated by intense rain, as the season progresses.
One important function of birds’ nests is to provide microclimate conditions for adults, nestlings, and eggs (Mainwaring et al. Citation2014). For closed-cup and cavity-nesting birds, the entrance orientation plays an important role in determining the interaction between ambient conditions and optimization of the microclimate within the nest (Long et al. Citation2009, Landler et al. Citation2014, Martin et al. Citation2017, Schaaf et al. Citation2020). Nest placement and orientation are therefore likely to be adaptive or plastic traits and should vary as a function of environmental conditions (Mainwaring et al. Citation2014, Schaaf et al. Citation2020).
By adjusting the structure, position, or orientation of the nest, birds can respond to these climatic variables during their breeding season in different ways, either by orienting their nests against the prevailing winds and solar radiation or by building their nests in sites with dense vegetation cover to protect them from these climatic variables (Hoekman et al. Citation2002, Souza & Santos Citation2007, Schaaf et al. Citation2018). Previous work found that in temperate sites with low diurnal and nocturnal temperatures, birds orient their nests to take advantage of the solar radiation during the day, thereby increasing the temperature within the nests to avoid cooling. While in high solar exposure sites, near the equator, they tend to avoid direct solar radiation to avoid eggs overheating during incubation (Burton Citation2007, Landler et al. Citation2014, Schaaf et al. Citation2018). Furthermore, it was also found that in tropical and subtropical sites with heavy rainfall and humidity, birds orientated their nests to benefit from solar exposure to reduce humidity within their nests (Greeney Citation2009, Schaaf Citation2020). As for vegetation cover, this variable may promote random orientation, as nests are well-protected from wind, rain, and solar radiation that may negatively impact their reproduction (Souza & Santos Citation2007, Schaaf et al. Citation2018).
Despite these advances, few studies have analysed the variation in nest orientation throughout a breeding season, when climate might vary along its duration (Mezquida Citation2004, Long et al. Citation2009). However, birds have been shown to orientate their nest entrances against the prevailing winds and solar radiation in temperate dry sites in central Argentina (Mezquida Citation2004).
This paper explores the orientation of closed-cup nests of the Rufous Hornero Furnarius rufus in a humid subtropical urban site of the Neotropics throughout its breeding season in relation to local climatic factors, specifically wind direction, rainfall, and solar radiation. At the beginning of the breeding season of this species, rainfall is less intense, cold southeastern winds predominate, and temperatures are low, so it is predicted that: (1) nest orientation that takes advantage of increased solar radiation and protects against the prevailing cold winds, in order to increase the temperature within the nest. As the breeding season progresses, rainfall increases, so it is predicted that (2) later in the breeding season, nest orientation should be towards direct solar exposure and warm winds to reduce the humidity of the nesting micro-site (Wiebe Citation2001, Hoekman et al. Citation2002, Greeney Citation2009).
Methods
Study site
The work was carried out in the city of San Salvador de Jujuy, a subtropical site in northwest Argentina (24°11′S 65°17′W, ). This site is part of the Yungas Forest ecoregion, located between 1300 and 1500 m above sea level. The climate is dry and cold during winter and early spring months (July to September) and it has heavy rainfall and high temperatures during the subsequent spring and summer months (mid-October to March) (Grau Citation2005, Malizia et al. Citation2014).
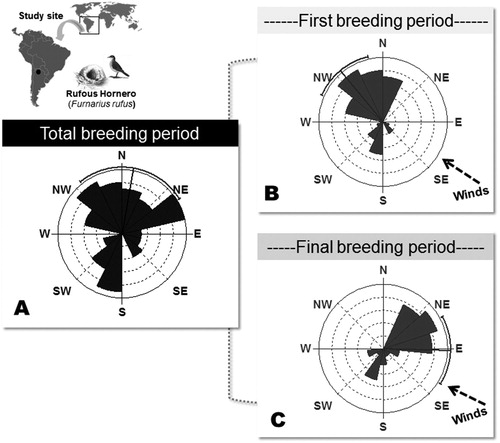
Figure 1. Circular distribution of the entrances of Rufous Hornero nests in a subtropical urban site in the city of San Salvador de Jujuy, Argentina. The mean orientation (±SE) and the total amount of observations (numbers in the bars) are shown: (A) nests found during the whole breeding season (n = 74), (B) nests found during the first reproduction period (n = 32), and (C) nests found during the second reproduction period (n = 42).
Study species
The Rufous Hornero is a species of the Furnariidae family, widely known as ovenbirds, that build closed-cup nests made of mud and vegetal fibres (Zyskowski & Prum Citation1999). These nests are often found in cities and suburban locations, both on trees and on lamp post (Ridgely & Tudor Citation1994). The species is widely distributed in South America, including Bolivia, Brazil, Argentina, Uruguay, and Paraguay, and their nesting season is from September to March (Fraga Citation1980, Peña Citation2005, Shibuya et al. Citation2015). Incubation lasts about 16–17 days and the chicks remain in the nest for 20–23 days (Peña Citation2005). In the location where this work was carried out, it was more frequent to find nests in trees and, to a lesser extent, on lamp posts (Schaaf et al. Citation2018).
Data collection and statistical analysis
From a local weather station (Augusto M. Romain weather station), it was possible to obtain data on rainfall, wind direction and temperature (morning: 09:00; afternoon: 15:00; night: 03:00). The breeding season was split into two periods, in relation to local climate: the first two nesting months of this species (September and October of 2018, a cold period with low rainfall), and the remaining months (November 2018 to February 2019, a wet and warm period). A chi-square test was used (χ2) to identify whether there were significant differences between the total rainfall events among two periods. The differences between period temperatures (day and night) were assessed with the Tukey test (t), using INFOSTAT (Di Rienzo et al. Citation2008). Circular statistics were used to estimate wind direction (±SD) for each reproductive period. A Watson–Williams F test was used to estimate differences in mean orientation values due to wind, using Oriana software (Kovach Citation2004).
Rufous Hornero nests were searched for in trees in the city of San Salvador de Jujuy from September 2018 to February 2019. Active nests, which were most frequent, were systematically searched for along transects during the breeding season (Schaaf et al. Citation2018). The entrance orientation of each nest was recorded with a compass. These tree-located nests were well covered by lateral and upper vegetation (Schaaf et al. Citation2018). All the orientation data were corrected according to the geodetic north (Evans Citation2017).
A nest was considered to be active when parental activity was observed within (parents entering and leaving the nest, chick provisioning). Those nests found during incubation in September and active nests with chicks found at the end of October were included in the first period. Later nests were included in the second period, but nests with incubation during late October were not included in the analysis; that way, it was possible to divide the breeding period to compare it with local climatic variables.
For each period, the nest entrance mean orientation (±SD) was estimated using circular statistics. For total and each breeding period, the Rayleigh test was used to determine whether the nest entrances were randomly distributed or non-randomly distributed (Zar Citation1999). Finally, a Watson–Williams (F) test was performed to estimate differences in mean orientation between breeding periods. All the analyses were performed using Oriana software (Kovach Citation2004).
Results
During the first nesting period (September and October of 2018), the weather was cold with rainfall recorded on 10 non-consecutive days. For the remaining months (November 2018 to February 2019), rainfall was recorded during 51 non-consecutive days. The comparison between mean of wind direction did not show significant differences between periods, during which the winds were predominantly from the southeast. The comparison between mean temperatures showed significant differences in the morning and night time but no significant differences were found in the afternoon temperatures between periods ().
Table 1. Local climatic variables measured during the first nesting period (September and October of 2018, cold with low rainfall), and the second breeding period (November 2018 to February 2019, warm with high rainfall), in the city of San Salvador de Jujuy, Argentina. It shows total rainfall (range), mean ± SD (range) temperatures, and wind direction (±SD). Statistical analysis: chi-square test (χ2), the Tukey test (t), and Watson-Williams test (F).
A total of 74 nests were found: 32 during the first breeding period and 42 during the second period. The mean entrance orientation of all nests exhibited random orientation (9.9° ± 104.7, Rayleigh Test = 2.62, P = 0.07, n = 74). However, nests built in the first period of the breeding season had a significantly non-random mean entrance orientation (northwest: 320.3° ± 73.77, Rayleigh Test = 8.00, P = 0.002, n = 42), as did the nests built during the second period (east: 91.5° ± 73.3, Rayleigh Test = 6.22, P < 0.001, n = 32; ). The orientation of nests differed significantly between these two periods (Watson–Williams F test, F = 34.34, P < 0.001).
Discussion
The results indicate that the Rufous Hornero builds nests with a northwest orientation in the first half of the breeding season, when the weather is cold but dry, and switches to an eastern orientation in the second half of the breeding season, when the weather is warmer but much wetter. Previous studies in other bird species found that non-random nest entrance orientation was lined with the maintenance of the micro-climatic within nests, especially at wet and cold sites (Burton Citation2006, Mainwaring et al. Citation2016, Schaaf et al. Citation2018, Citation2020, Tomiałojć & Neubauer Citation2018). For the Rufous Hornero, nests built at the beginning of the breeding season have to face low temperatures during day and night. Therefore, a northwest orientation would help to accumulate heat during the afternoon to compensate for low night temperatures (Wiebe Citation2001, Burton Citation2007), especially when considering that the nests were located in sites with dense vegetation cover and, therefore, in receipt of less solar radiation (Schaaf et al. Citation2018). Furthermore, these nests were oriented against the prevailing cold southeast winds, hence evidencing that the winds were not providing them with thermoregulation benefits.
As the breeding season progresses, the rainfall and the day and night temperatures increase. Considering that the vegetation cover provides protection against solar radiation and rainfall, the surrounding vegetation can also generate humid microsites around the nest. Exposure to high humidity can negatively affect the development of the eggs during reproduction, as birds require the maintenance of specific temperatures and humidity ranges within nests (Greeney Citation2009, Mainwaring et al. Citation2014, Martin et al. Citation2017). So, by orienting the nest entrances to the northeast quadrant, the Rufous Hornero can take advantage of the morning temperatures, reduce the humidity and avoid the high afternoon temperatures (Wiebe Citation2001, Burton Citation2007). During this period, they also seem to be sensitive to local winds, and orientating their nest entrances against the prevailing winds could provide them with structural advantages (e.g. protection from nest rupture) in nest construction (Long et al. Citation2009).
Finally, the mean difference found between the established breeding seasons highlights the importance of studying how the orientation varies during the entire breeding period (Long et al. Citation2009). When this same species was studied in the same location, nests with dense vegetation cover were found to be orientated randomly (Schaaf et al. Citation2018). Hence, climate variations throughout the breeding period result in variations in nest entrance orientation (Hoekman et al. Citation2002, Long et al. Citation2009) but that pattern is hidden unless breeding periods are also considered. This orientation could be a way for the birds to take advantage of solar radiation to increase the temperature within the nest during the first cold months of their reproduction and to reduce nest humidity towards the end of the breeding season. Future studies that include breeding success and temperature inside the nest shall provide greater evidence of the relationship between climate effects and nest orientation (Rauter et al. Citation2002, Ardia et al. Citation2006, DuRant et al. Citation2013, Shibuya et al. Citation2015).
Acknowledgements
I would like to thank the reviewers for their work and comments that helped to improve this manuscript, Marisel Morales who kindly made the drawings, and Fabio Alabar for provide me the climatic data. AAS is a postdoctoral fellow at CONICET.
References
- Ardia, D.R., Pérez, J.H. & Clotfelter, E.D. 2006. Nestbox orientation affects internal temperature and nest site selection by Tree Swallows. J. Field Ornithol. 77: 339–344. doi: 10.1111/j.1557-9263.2006.00064.x
- Burton, N.H. 2006. Nest orientation and hatching success in the Tree Pipit Anthus trivialis. J. Avian Biol. 37: 312–317. doi: 10.1111/j.2006.0908-8857.03822.x
- Burton, N.H. 2007. Intraspecific latitudinal variation in nest orientation among ground-nesting passerines: a study using published data. Condor 109: 441–446. doi: 10.1093/condor/109.2.441
- Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M. & Robledo, C.W. 2008. Software Infostat, versio´n 2008. Grupo InfoStat, FCA, Universidad Nacional de Co´rdoba, Argentina.
- DuRant, S.E., Hopkins, W.A., Hepp, G.R. & Walters, J.R. 2013. Ecological, evolutionary, and conservation implications of incubation temperature-dependent phenotypes in birds. Biol. Rev. 88: 499–509. doi: 10.1111/brv.12015
- Evans, S.W. 2017. The effect of nest site orientation on the breeding success of Blue Swallows Hirundo atrocaerulea in South Africa. Afr. J. Ecol. 56: 91–100. doi: 10.1111/aje.12421
- Fraga, R.M. 1980. The breeding of Rufous Horneros (Furnarius rufus). Condor 82: 58–68. doi: 10.2307/1366785
- Grau, H.R. 2005. Dinámica de bosques en el gradiente altitudinal de las Yungas Argentinas. In M. Arturi, J. Frangi & J. Goya. (ed) Ecología y Manejo de los Bosques Argentinos, 181–188. Universidad Nacional de La Plata, La Plata.
- Greeney, H.F. 2009. Nest orientation of the spotted barbtail, Premnoplex brunnescens, is strongly correlated with stream flow. J. Ethol. 27: 203–208. doi: 10.1007/s10164-008-0104-1
- Hoekman, S.T., Ball, I.J. & Fondell, T.F. 2002. Grassland birds orient nests relative to nearby vegetation. Wilson Bull. 114: 450–456. doi: 10.1676/0043-5643(2002)114[0450:GBONRT]2.0.CO;2
- Kovach, W. 2004. Oriana v. 2.02 a. Kovach Computing Services, Anglesey, Wales.
- Landler, L., Jusino, M.A., Skelton, J. & Walters, J.R. 2014. Global trends in woodpecker cavity entrance orientation: latitudinal and continental effects suggest regional climate influence. Acta Ornithol. 49: 257–266.
- Long, A.M., Jensen, W.E. & With, K.A. 2009. Orientation of Grasshopper Sparrow and Eastern Meadowlark nests in relation to wind direction. Condor 111: 395–399. doi: 10.1525/cond.2009.080076
- Mainwaring, M.C., Hartley, I.R., Lambrechts, M.M. & Deeming, D.C. 2014. The design and function of birds’ nests. Ecol. Evol. 4: 3909–3928. doi: 10.1002/ece3.1054
- Mainwaring, M.C., Barber, I., Deeming, D.C., Pike, D.A., Roznik, E.A. & Hartley, I.R. 2016. Climate change and nesting behavior in vertebrates: a review of the ecological threats and potential for adaptive responses. Biol. Rev. 92: 1991–2002. doi: 10.1111/brv.12317
- Malizia, L.R., Bergesio, L. & Fierro, P. 2014. Ambiente y sociedad en la comarca de Yala. Centro de Estudios Territoriales Ambientales y Sociales, Facultad de Ciencias Agrarias, Facultad de Humanidades y ciencias Sociales, Universidad Nacional de Jujuy. Editorial de la Universidad Nacional de Jujuy, San Salvador de Jujuy, Argentina. Ediciones del Subtrópico de la Fundación ProYungas, Yerba Buena, Tucumán, Argentina.
- Martin, T.E., Boyce, A.J., Fierro-Calderón, K., Mitchell, A.E., Armstad, C.E., Mouton, J.C. & Bin Soudi, E.E. 2017. Enclosed nests may provide greater thermal than nest predation benefits compared with open nests across latitudes. Funct. Ecol. 31: 1231–1240. doi: 10.1111/1365-2435.12819
- Mezquida, E.T. 2004. Nest site selection and nesting success of five species of passerines in a South American open Prosopis woodland. J. Ornithol. 145: 16–22. doi: 10.1007/s10336-003-0002-9
- Peña, M.R.D.L. 2005. Reproducción de las aves argentinas (con descripción de pichones). LOLA, Buenos Aires. 845 pp.
- Rauter, C.M., Reyer, H.U. & Bollmann, K. 2002. Selection through predation, snowfall and microclimate on nest-site preferences in the Water Pipit Anthus spinoletta. Ibis 144: 433–444. doi: 10.1046/j.1474-919X.2002.00013.x
- Ridgely, R.S. & Tudor, G. 1994. The birds of South America. The suboscine passerines, vol II. University of Texas Press, Austin.
- Schaaf, A.A. 2020. Effects of sun exposure and vegetation cover on Woodpecker nest orientation in subtropical forests of South America. J. Ethol. 38: 117–120. doi: 10.1007/s10164-019-00617-2
- Schaaf, A.A., García, C.G., Puechagut, P.B., Silvetti, L.E., Tallei, E., Ortis, F. & Quaglia, A.I. 2018. Effect of geographical latitude and sun exposure on Rufous hornero (Furnarius rufus) nest orientation. J. Ornithol. 159: 967–974. doi: 10.1007/s10336-018-1569-5
- Schaaf, A.A., Garcia, C.G. & Greeney, H.F. 2020. Nest orientation in closed nests of Passeriformes across a Latitudinal Gradient in the Southern Neotropic. Acta Ornithol. 54: 263–268. doi: 10.3161/00016454AO2019.54.2.012
- Shibuya, F.L., Braga, T.V. & Roper, J.J. 2015. The Rufous Hornero (Furnarius rufus) nest as an incubation chamber. J. Therm. Biol. 47: 7–12. doi: 10.1016/j.jtherbio.2014.10.010
- Souza, F.L. & Santos, C.A. 2007. Climate and nest opening orientation in Furnarius rufus (Furnariidae). Iheringia 97: 293–295. doi: 10.1590/S0073-47212007000300013
- Tomiałojć, L. & Neubauer, G. 2018. Song Thrush Turdus philomelos and Hawfinch Coccothraustes coccothraustes exhibit non-random nest orientation in dense temperate forest. Acta Ornithol. 52: 209–221. doi: 10.3161/00016454AO2017.52.2.008
- Wiebe, K.L. 2001. Microclimate of tree cavity nests: is it important for reproductive success in Northern Flickers? Auk 118: 412–421. doi: 10.1093/auk/118.2.412
- Zar, J.H. 1999. Biostatistical Analysis. Prentice Hall, Upper Saddle River.
- Zyskowski, K. & Prum, R.O. 1999. Phylogenetic analysis of the nest architecture of Neotropical ovenbirds (Furnariidae). Auk. 116: 891–911.