
ABSTRACT
The effect of perching versus hovering on the drinking speed of the Cameroon Sunbird Cyanomitra oritis and the Northern Double-collared Sunbird Cinnyris reichenowi was investigated in a cage experiment. Our results support the hypothesis that the shorter handling times on flowers during hovering compared to perching are largely caused by an increase in drinking speed.
The drinking speed of nectar-feeding birds is an important parameter, which is expected to be affected by both floral and bird traits. The most often reported plant traits involved are the sugar concentration of nectar, nectar volume, and flower tube length and shape. Drinking speed is usually highest at 30–50% concentrations of sucrose and is slower at both lower and higher concentrations (Mitchell & Paton Citation1990). The speed of nectar extraction from a flower is also related to a range of flower traits, including flower tube depth, width, and orientation (Collins Citation2008, Ngcamphalala et al. Citation2018).
Of the bird traits that affect drinking speed, bill length, tongue anatomy, and body size have been studied (Collins & Paton Citation1989). Generally, long-billed species can more effectively exploit long-tubed flowers and obtain richer rewards than short-billed species. Even if short-billed species are able to reach the nectar, they are forced to reduce drinking speed more than long-billed species, because the volume taken per lick decreases with distance to the nectar (Collins Citation2008). The speed is determined by tongue capillary properties, the amount of nectar captured per lick and licking frequency (Paton & Collins Citation1989, Collins Citation2008) and can also be influenced by bird size (Mitchell & Paton Citation1990).
In contrast, the effect of bird behaviour on drinking speed has not been studied widely. Hovering during feeding is well known in American hummingbirds (Feinsinger & Chaplin Citation1975), and although previously not well recognized, is relatively common among Old-World nectarivorous bird species (Wester Citation2014). Hovering birds can acquire nectar from otherwise inaccessible flowers, which typically means those with long pedicels, and a pendant or horizontal corolla orientation (Miller Citation1985, Padyšáková & Janeček Citation2016). Hovering, compared to perching, is also more energetically demanding and may affect net energy gain (Wolf et al. Citation1975) and even hummingbirds, which are adapted to hovering, prefer to feed on a perch whenever possible (Miller Citation1985). Furthermore, body size also affects the energetic cost of hovering, and feeding while hovering may be unprofitable for larger birds (Collins & Paton Citation1989).
To support the ongoing discussion on factors affecting drinking speed and subsequent flower handling times, this study focused on the effect of behaviour (perching versus hovering) on drinking speed. Two sunbird species were chosen that share a similar environment and food sources but differ in body mass, bill length, and in their frequency of hovering (Janeček et al. Citation2011).
Drinking speed was studied in relation to hovering and perching behaviour in 17 individual adult Cameroon Sunbirds Cyanomitra oritis and 17 individual adult Northern Double-collared Sunbirds Cinnyris reichenowi. Cameroon Sunbirds are the larger species (12 g) with a longer bill (28–33 mm), compared to the smaller (9 g) Northern Double-collared Sunbird which has a shorter bill (15–22 mm). Cameroon Sunbirds more often hover while drinking from Impatiens spp. than Northern Double-collared Sunbirds (Janeček et al. Citation2011, Citation2015). Individual sunbirds were caught by mist-net at Irumambai, near the forest line on Mount Cameroon, Cameroon (2050 m above sea level, 4.147° N 9.173° E) in May 2018. Each bird was placed individually inside a cage (1 × 1 × 1.7 m) as soon as it was caught, for a feeding trial, and was released within one hour of capture. Individuals were not tested repeatedly. In the cage, each bird was offered a flower of Impatiens sakeriana, a plant in the family Balsaminaceae which is native to Cameroon and Equatorial Guinea, and which are often visited and pollinated by both sunbird species (Janeček et al. Citation2011). The spur of the flower was replaced with a pipette tip filled with a solution of 30% w/w sucrose, a common nectar concentration in flowers pollinated by birds (Nicolson & Fleming Citation2003). By different arrangements of the flower and branches in the cage, birds were able to feed by perching or only by hovering. Drinking birds were recorded using a GoPro Hero5 camera at a speed of 120 frames per second; however, the behaviour was still blurred at this rate so we calculated the average drinking speed from the volume of nectar consumed and the time spent drinking. Drinking speed was log-transformed to achieve a normal distribution. The foraging depth was defined as the distance between the open end of the pipette tip and the level of the sugar solution. The data were analysed using a generalized linear model using the Statistica software version 13.5 (TIBCO Software Inc. Citation2018).
Drinking speed was significantly different between the two sunbird species as well as between hovering and perched individuals of the same species ( and , ). When hovering, the values were on average 1.2 times (for Cameroon Sunbird) and 1.6 times (for Northern Double-collared Sunbird) higher than when perched (). Because of the possible influence of foraging depths at the beginning of drinking and variability in solution volume consumed, both these variables were used as covariates in the models (). The effect of behaviour did not differ between species, as the interaction between these two variables was not significant. In both species, drinking speed was higher when birds were hovering and lower when they were perched (, ). Cameroon Sunbirds drank faster and more than Northern Double-collared Sunbirds ( and ), but the total volume consumed did not differ between hovering and perching birds of the same species ((C)).
Figure 1. Drinking speed of Cameroon sunbirds and northern double-collared sunbirds while hovering or perching. The whiskers represent the standard deviation.
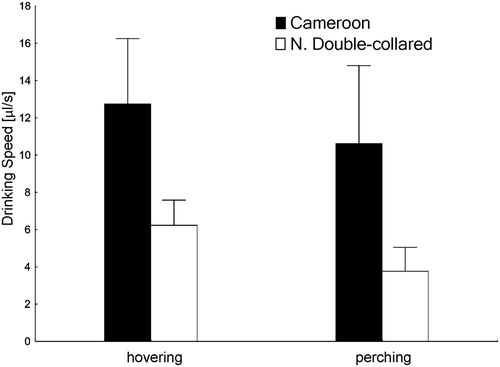
Table 1. Summary of generalized linear models describing the effects on: (A) drinking speed (log-transformed), (B) time and (C) volume.
Table 2. Length of drinking events (time), volume of sugar solution consumed during drinking events (volume) and drinking speed (Speed) in the Cameroon sunbird and the northern double-collared sunbird.
We found both large interspecific and intraspecific variation (1.9–17.9 µL/s) in drinking speed and revealed that bird locomotion during drinking can significantly affect drinking speed. As predicted, hovering birds had a higher drinking speed, and it is assumed that this contributed to the decrease in flower handling time (Padyšáková & Janeček Citation2016; ). Although hovering during foraging is energetically more demanding than perching, it may shorten the time required for moving between flowers and potentially allow more flowers to be visited per hour, while also enabling birds to access food sources where no perches are available (Miller Citation1985, Janeček et al. Citation2011). Whether hovering is beneficial for birds from an energetic point of view, or if it is just an artefact of plant morphology, is still under discussion (Miller Citation1985, Padyšáková & Janeček Citation2016). Data on the impact of different behaviours on drinking speed can help to develop a more detailed energetic model towards answering this question. Moreover, as well as increasing foraging efficiency, hovering could have the additional benefit of reducing predation risk (Schneider Citation1984). Nevertheless, this study was not able to disentangle the causality of a bird drinking faster when hovering: it could have been drinking faster to offset the energetic costs of hovering or hovering might have enabled faster nectar intake.
The behaviour of nectarivorous species has rarely been studied under natural conditions (Wolf et al. Citation1975, Janeček et al. Citation2011, Citation2015). Both species compete for the same nectar sources, and even small differences in drinking speed could change the exploitation of the rich nectar source that flowers of Impatiens sp. offer. However, it would be beneficial to have more studies of nectar drinking behaviour from a wider range of species so that we can better understand the adaptive significance of physical traits and foraging behaviour under natural conditions.
Acknowledgements
The authors thank the municipality of Bokwango for access to its community forest and Jiří Mlíkovský for valuable comments on the manuscript.
Additional information
Funding
References
- Collins, B.G. 2008. Nectar intake and foraging efficiency: responses of honeyeaters and hummingbirds to variations in floral environments. Auk 125: 574–587.
- Collins, B.G. & Paton, D.C. 1989. Consequences of differences in body mass, wing length and leg morphology for nectar-feeding birds. Austr. J. Ecol. 14: 269–289.
- Feinsinger, P. & Chaplin, S.B. 1975. On the relationship between wing disc loading and foraging strategy in hummingbirds. Am. Nat 109: 217–224.
- Janeček, Š, Patáčová, E., Bartoš, M., Padyšáková, E., Spitzer, L. & Tropek, R. 2011. Hovering sunbirds in the Old World: occasional behaviour or evolutionary trend? Oikos 120: 178–183.
- Janeček, Š, Bartoš, M. & Njabo, K.Y. 2015. Convergent evolution of sunbird pollination systems of Impatiens species in tropical Africa and hummingbird systems of the new world. Biol. J. Linn. Soc 115: 127–133.
- Miller, R.S. 1985. Why hummingbirds hover. Auk 102: 722–726.
- Mitchell, R.J. & Paton, D.C. 1990. Effects of nectar volume and concentration on sugar intake rates of Australian honeyeaters Meliphagidae. Oecologia 83: 238–246.
- Ngcamphalala, C.A., Bailey, I.E. & Nicolson, S.W. 2018. Nectar intake and foraging efficiency: the responses of sunbirds to flower morphology. J. Ornit. 159: 1031–1041.
- Nicolson, S.W. & Fleming, P.A. 2003. Nectar as food for birds: the physiological consequences of drinking dilute sugar solutions. Plant Sys. Evol. 238: 139–153.
- Padyšáková, E. & Janeček, Š. 2016. Sunbird hovering behavior is determined by both the forager and resource plant. Biotropica 48: 687–693.
- Paton, D.C. & Collins, B. 1989. Bills and tongues of nectar-feeding birds: a review of morphology, function and performance, with intercontinental comparisons. Aust. J. Ecol. 14: 473–506.
- Schneider, K.J. 1984. Dominance, predation, and optimal foraging in white-throated sparrow flocks. Ecology 65: 1820–1827.
- TIBCO Software Inc. 2018. Statistica data analysis software system, version 13.5 <http://tibco.com>.
- Wester, P. 2014. Feeding on the wing: hovering in nectar-drinking Old World birds – more common than expected. Emu 114: 171–183.
- Wolf, L.L., Hainsworth, F.R. & Gill, F.B. 1975. Foraging efficiencies and time budgets in nectar-feeding birds. Ecology 56: 117–128.