
ABSTRACT
Capsule
Long-term data on changes in avian diversity from urban habitats are essential to address the ongoing declines in European bird populations.
Aims
We aimed to analyse historical changes in the bird assemblage of an urban district of Bonn, Germany, over 50 years.
Methods
We surveyed the breeding bird population in a district of Bonn, Germany, and compared our results in terms of species richness, abundance, habitat type, and biomass with results from a survey conducted exactly 50 years earlier. To compare changes in habitat composition we analysed historical and current aerial photos.
Results
Between surveys, there was a severe decline in species richness and a strong homogenization of the bird assemblage while the overall number of breeding birds slightly increased. Biomass increased but was strongly influenced by the massive increase of the Common Woodpigeon Columba palumbus. Many forest species that rely on older trees increased in abundance while the typical species of settlements and open habitats strongly decreased. However, large-scale habitat changes are not sufficient to explain these patterns.
Conclusions
Urbanization of Central European cities has boosted populations of generalist species while those already of conservation concern will further decline if their demands are not considered in current urban planning approaches. Our work highlights that historical data on breeding bird abundance are of high importance and should be sought and used. In our case such data demonstrated the impact of small-scale changes in habitat and other parameters highlighting that these should receive more attention.
The destruction of natural environments by agricultural changes, urban growth, and urbanization has been recognized as a threat to global biodiversity that currently may largely exceed values known in former human history (McKinney Citation2002, Ceballos et al. Citation2015). Animal species that are able to adapt their ecological requirements through evolutionary change or phenotypic plasticity can be regarded as ‘winners’ while many species cannot cope with environmental changes and ultimately may become extinct locally or at least increasingly rare. While the latter can be described as ‘urban avoiders’, among the ‘winners’ are ‘suburban adaptable species’ and ‘urban exploiters’ sensu Blair (Citation1996). Urbanized areas are often similar to each other in terms of species composition, and also harbour a diminished species assemblage; the result of a process referred to as biotic homogenization (McKinney & Lockwood Citation1999, Clergeau et al. Citation2006, Olden & Rooney Citation2006). Biotic homogenization due to urbanization has been described for many groups of organisms (McKinney & Lockwood Citation1999, Abrahamczyk et al. Citation2020), and particularly for birds (Marzluff et al. Citation2001a, Lepczyk & Warren Citation2012, Murgui & Hedblom Citation2017). In contrast to other taxonomic groups, many bird species inhabiting urbanized areas are well known because they are conspicuous, comparatively easy to study and, arguably, the best-known taxonomic group with a plethora of professional and citizen scientists conducting avian research (Wei et al. Citation2016). Urbanization influences nearly all aspects of avian ecology, including demography, productivity, individual weights, and bioacoustics (Chace & Walsh Citation2006, Chamberlain et al. Citation2009). Urban bird species typically have large distribution ranges, a preference for dense vegetation and are medium-sized, sedentary, omnivorous, and high-vegetation nesters (Croci et al. Citation2008, Conole & Kirkpatrick Citation2011). However, even threatened species can inhabit urbanized town centres (Jokimäki et al. Citation2020). Moreover, bird biomass changes along urban gradients, often with a peak at intermediate levels of urbanization (Blair Citation1996) or, more generally, an increase in biomass despite a reduction in richness (Chace & Walsh Citation2006).
Additionally, urbanization affects the functional diversity of avian communities even on a global scale (Sol et al. Citation2020). In comparison to rural areas, phylogenetic diversity in terms of species uniqueness can also be depauperated in cities (Morelli et al. Citation2016, Ibáñez-Álamo et al. Citation2017). Assemblages of urban birds have strongly changed during the last decades due to the structural change seen in many cities (Nowakowski Citation1996, Rheinwald Citation2005). While a few decades ago Central European cities were still heterogeneous landscapes, composed of a mosaic of true urban housing, gardens, orchards, fallow land, industrial, and forested areas, the strong increase in the urban population led to a decline of less intensively used areas within cities (Kelcey & Rheinwald Citation2005). A consequence of this is that urban bird assemblages often decline in species richness, and especially so in regard of habitat specialists (Bezzel Citation1985, Bonier et al. Citation2007, Devictor et al. Citation2007). Despite this, many studies of urban gradients report a peak in species richness at intermediate levels of urbanization (Jokimäki & Suhonen Citation1993, Blair Citation1996).
Effects of urbanization on avian communities can be analysed in at least two ways. First, from a spatial perspective by studying species along gradients of different land use types from natural habitats to heavily urbanized areas (Erz Citation1966, McDonnell & Pickett Citation1990, Blair Citation1996, Chace & Walsh Citation2006, Pennington & Blair Citation2012). These studies can be conducted in a clear time frame, as theoretically all the data can be collected at the same time. Secondly, and in contrast to the first approach, a temporal perspective can be achieved through analyses of long-term data and historical data. The latter has the advantage of being unbiased by any spatial factors. However, while urbanization has been intensively studied in recent years (Lepczyk & Warren Citation2012), few data on species abundance go back more than a few decades and allow for an analysis of changes over a long period (Marzluff et al. Citation2001b, Suhonen et al. Citation2009, Fidino & Magle Citation2017). In addition, most modern breeding bird monitoring schemes in Europe only date back to the 1970s or 1980s (e.g. Pan-European Common Bird Monitoring Scheme, www.pecbms.info).
Here, we report the findings of a breeding bird survey carried out in 2019 in a suburban district of Bonn, a typical Central European city. A comparable survey took place in 1969 (Wink Citation1971), thus offering the opportunity to compare avian communities from the same area across half a century. In our study we analyse changes in (1) species richness and (2) species composition and relate these to changes in habitat composition. We expected to find a strong decrease in species richness and abundance, as well as a homogenization of the bird assemblage in parallel to a homogenization of habitat types. Moreover, (3) as a species-specific parameter, we check whether the overall biomass of the avian community has changed over time. We expected to find a moderate increase of the total body mass of the community because the degree of urbanization slightly increased between the two survey periods.
Methods
Study site
The city of Bonn is situated in the western part of Germany (), mostly located at 60 m above sea level, and in the Rhine valley, though reaching 171 m above sea level at the Venusberg district and the adjacent Kottenforst. Bonn has an Atlantic climate with mild winters and comparatively warm, humid summers. Due to climate change, mean annual temperature has increased by 1.2°C in Bonn since 1895 and mean annual precipitation has increased by approximately 150 mm per year (www.bonn.de). The increase in temperature is clearly visible when comparing the mean daily temperatures for the two study periods, 1969 and 2019 (), so the weather conditions of both years can be regarded as typical for their decade. These data were taken from www.kachelmannwetter.de.
Figure 1. Map of Europe showing the position of the city of Bonn (Eurostat Citation2020, Bundesamt für Kartographie und Geodäsie Citation2020). The red polygon within the city of Bonn (black polygon) indicates the district of Dottendorf (Stadt Bonn Citation2020, Geobasis NRW Citation2020).
Figure 2. Mean daily temperature and mean daily precipitation per month of the two study periods 1969 and 2019.
Bonn is a city founded over 2000 years ago by the Romans. Some parts of the city were destroyed during World War II and many areas were rebuilt with architecture typical of the 1950s to 1980s. The human population has nearly constantly increased over the last decades and the city is regarded as a regional metropolis. With more than 50% of settlement and transport space in the urban area, Bonn is very densely populated and thus lies above the average density of North Rhine-Westphalian cities (Mutke et al. Citation2019). The ornithology of Bonn has been well documented (Rheinwald et al. Citation1984, Citation1987, Rheinwald & Kneitz Citation2002, Rheinwald Citation2005).
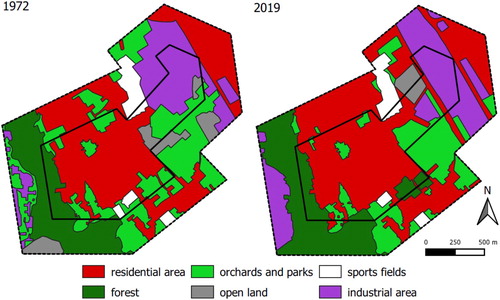
The district of Dottendorf is a suburban area in Bonn, near a mixed deciduous forest (Kottenforst). Until 1904, Dottendorf was an independent village surrounded by orchards, meadows, and vineyards, which was then incorporated into the city of Bonn. Especially after World War II, in the course of Bonn’s development into the capital, many village and rural structures were lost (Wichmann & Wichmann Citation2020). The population figures in Dottendorf have increased only moderately from 5312 (1970) to 5743 inhabitants (2019; Bundesstadt Bonn, Statistikstelle Citation2020a, Citation2020b). Today, Dottendorf is closely linked to the neighbouring districts by residential development and infrastructure, into which the vineyards, meadows, and most of the orchards have been converted; the historic structures are visible on an historical aerial photograph (1967; ) and partly remain today (). For example, village-like structures with larger gardens exist close to two- to five-storeyed houses.
Figure 3. Historical aerial photo with the southwest part of Dottendorf taken in 1964 (Source: Stadtarchiv und Stadthistorische Bibliothek Bonn; Photographer: Aero-Lux Büscher & Co. KG, Frankfurt).

Figure 4. Aerial view of Dottendorf with adjoining district at the right from 2017, showing most of the mapping area except the eastern part; on the left margin the sports field existing already on the historical aerial view is visible (Source: Wikipedia, Wolkenkratzer).

We analysed the change in land use cover using current (from 2019; resolution 10 cm × 10 cm/pixel) and historical (from 1972; resolution 40 cm × 40 cm/pixel) aerial land photos (photos are not shown) provided by the district government of Cologne. We included our study area (103 ha) as well as a 300 m matrix around the study area as we did not want to rule out the possibility that avian diversity structure was affected by surrounding habitats. Structures on the aerial land photos were related to six land use types: residential area, industrial area, sports fields, orchards and parks, open land (meadows and uncultivated land), and forest. The visual interpretation of the aerial land photos was chosen for the classification because of the comparatively small size of the study area. We calculated the total area of these six habitat types for the study area as well as for the matrix in the current and historical aerial land photos, resulting in quantified habitat changes from 1972 to the present.
Bird data
The Dottendorf area (103 ha) was intensively studied by Michael Wink (Wink Citation1971). He conducted a complete survey of the area in April and May 1969 and recorded whether birds showed typical territorial behaviour (singing, territorial defence, nest-building, etc.) and used a territorial mapping approach to delimit territories. The number of territories was defined as equal to the number of breeding pairs. We repeated the study in spring 2019, following a standard protocol (Bibby et al. Citation1995, Südbeck et al. Citation2005). We surveyed the total area of Wink’s study between February and the end of June 2019. In the western sub-area we conducted six breeding bird surveys, and in the eastern sub-area seven surveys. In addition, one evening was designated to search for owls and another one-day survey to search for breeding colonies of Common Swifts Apus apus. Fieldwork was conducted by JL, RS, and SA. While these standard protocols for territorial mapping were not published in 1969, Wink’s study was also very intensive (up to 10 site inspections). Therefore, we are convinced that both data sets are comparable. Four authors jointly evaluated the number of breeding territories based on the field data. Hence, any methodological bias should be small. For the two species with the largest changes, Common Blackbird Turdus merula and Common Woodpigeon Columba palumbus, Kathrin Schidelko, an experienced field ornithologist, repeated the delimitation of territories based on the field maps and came to very similar results (data not shown).
Based on the classification of ecological traits for the bird species of Germany (Wahl et al. Citation2014), we related four habitat categories to all recorded species of breeding birds: several habitat types (generalist), residential area, open land (orchards, ruderal area, meadows, and agricultural fields), and forest. This classification scheme is widely used by the Dachverband Deutscher Avifaunisten (umbrella organisation of German ornithologists) and was selected due to the local scope of our study. We merged the land use types settlements, industrial area, and sports field from our land use analysis into one habitat category and the land use types orchards and gardens and open land into a second, because they share a similar bird assemblage. Data for mean body weight per species were taken from Wahl et al. (Citation2014).
Statistical analysis
To analyse whether bird assemblages and habitat types changed in parallel, we calculated Bray–Curtis dissimilarity indices for bird species presence–absence data and species abundance data, as well as for the land use types of the study area and the 300 m matrix. In a second step we calculated Shannon diversity indices for the entire bird assemblage, as well as separately for open land-, forest-, settlement-, and generalist-species in both the historical and the current data sets. To study homogenization of the bird assemblage, we compared the species richness, bird abundance, and evenness values from the Shannon diversity index for both the historical and current data sets. Third, we analysed the change in breeding pairs per species between 1969 and 2019 for all 62 recorded species. We filtered those 15% of species with the strongest positive and negative changes each. Fourth, we used Fisher’s exact tests to compare the distribution of habitat types between the species with the strongest positive and negative changes and those species that went extinct, with the proportion of habitat types in the entire data set. Finally, we tested for significant differences in total biomass of breeding species (mean body weight per species x number of breeding pairs) between current and historical data using a paired Wilcoxon test. All tests were conducted in R 3.4.3 (R Development Core Team Citation2017).
Results
Between 1969 and 2019 the number of breeding bird species decreased from 57 to 39 in Dottendorf, although the total number of breeding pairs remained relatively constant (654 in 1969, 698 in 2019; online Table S1). During this period, 22 species became extinct and four new species colonized Dottendorf. Most of the species that became extinct in Dottendorf were present in 1969 with fewer than four breeding pairs; the exceptions were Willow Warbler Phylloscopus trochilus (6 pairs), Common Redstart Phoenicurus phoenicurus (8 pairs), European Serin Serinus serinus (15 pairs), and Tree Sparrow Passer montanus (30 pairs).
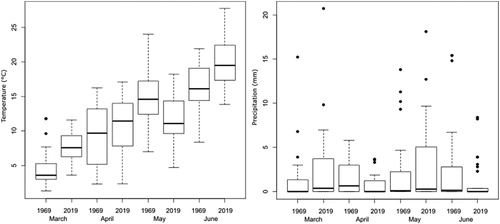
The species showing the strongest decrease in population size were Common Blackbird, House Sparrow Passer domesticus, Tree Sparrow, European Greenfinch Chloris chloris, European Serin, Common House Martin Delichon urbicum, Common Swift, Eurasian Collared Dove Streptopelia decaocto, and Common Redstart (). The decrease of breeding pairs of these species is counterbalanced by an increase in breeding pairs of Common Woodpigeon, Eurasian Blackcap Sylvia atricapilla, Great Tit Parus major, Blue Tit Cyanistes caeruleus, European Robin Erithacus rubecula, Common Firecrest Regulus ignicapilla, Eurasian Wren Troglodytes troglodytes, Song Thrush Turdus philomelos, and Dunnock Prunella modularis ().
Figure 5. Change in breeding pairs between 1969 and 2019 in Dottendorf. Dots to the right of the diagonal line indicate a decrease in breeding pairs while dots to the left of the line indicate species that showed an increase in breeding pairs. Black dots, black stars and black squares indicate the 5%, 10%, and respectively 15% of species that decreased/increased strongest. Strongest decrease: 1 = Common Blackbird Turdus merula, 2 = House Sparrow Passer domesticus, 3 = Tree sparrow Passer montanus, 4 = European Greenfinch Chloris chloris, 5 = European Serin Serinus serinus, 6 = Common House Martin Delichon urbicum, 7 = Common Swift Apus apus, 8 = Eurasian Collared Dove Streptopelia decaocto, 9 = Common Redstart Phoenicurus phoenicurus; Strongest increase: 10 = Common Woodpigeon Columba palumbus, 11 = Eurasian Blackcap Sylvia atricapilla, 12 = Great Tit Parus major, 13 = Blue Tit Cyanistes caeruleus, 14 = European Robin Erithacus rubecula, 15 = Common Firecrest Regulus ignicapilla, 16 = Eurasian Wren Troglodytes troglodytes, 17 = Song Thrush Turdus philomelos, 18 = Dunnock Prunella modularis.
Due to the considerable change in species composition and strong abundance changes of several species of the bird assemblage of Dottendorf between 1969 and 2019, we calculated a dissimilarity value of 30.0% using the presence–absence data and of 45.0% using abundance data (). In addition, the decrease in Shannon diversity indices and species evenness values between the surveys indicated a strong homogenization of the bird assemblage ().
Table 1. Dissimilarity analyses (Bray–Curtis) for land use types and breeding bird assemblage between 1969 and 2019 in Dottendorf.
Table 2. Species number, number of breeding pairs, and Shannon diversity index for the two surveys of breeding birds in Dottendorf.
The total area of semi-natural open land (orchards, ruderal area, meadows, and agricultural fields) decreased by 38.6% in our study site and 41.2% in the matrix, while the area of forest increased by 43.3% in the study site and 17.5% in the matrix (). The area of settlements and industrial production increased by 10.2% in our study site and by 24.6% in the matrix. Due to the much larger proportion of settlements, and the area of industrial production compared to semi-natural open land and forest, the change in land use types was only moderate in our study site (dissimilarity of 12.1%) and in the matrix (dissimilarity of 13.9%). These values therefore only partly reflect the strong changes in the bird assemblage.
Figure 6. Map of the study area Dottendorf (inner polygon) and the 300 m matrix with land use types.
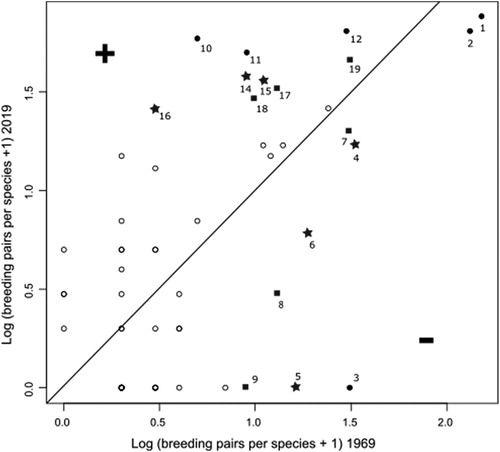
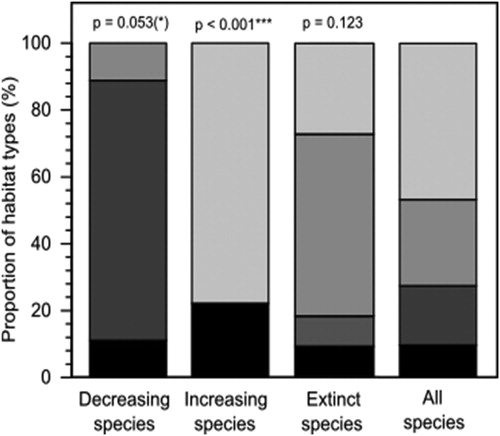
In the full data set, the distribution of species showing the strongest decrease in population is shifted towards species of residential areas and habitat generalists (). In contrast, the species showing the strongest increase in population size are shifted towards forest species (). Species that became extinct were generally open land species, but this result was not statistically significant.
Figure 7. Proportion of habitat types for the nine species (15%) with the strongest decrease in breeding pairs; the nine species (15%) with the strongest increase in breeding pairs; extinct species; and all species; black = several habitat types, dark grey = residential area, mid grey = open land, light grey = forest. P values indicate Fisher’s exact tests between the first three groups against D.
The total biomass of breeding birds in Dottendorf increased by 54.9% within 50 years. However, this increase in biomass was mostly due to the increase in the number of Common Woodpigeons. Excluding the Common Woodpigeon from the analysis revealed a decrease in total biomass of breeding birds by 16.7%. However, comparing the biomass of the historical with the current bird assemblage, we did not find a significant change in biomass, either when including (Wilcoxon tests, V = 814.5, P = 0.597) or when excluding (V = 814.5, P = 0.754) the Common Woodpigeon.
Discussion
Our study on the community of breeding birds in Bonn Dottendorf documented a strong decrease in species richness and evenness compared to the study of Wink (Citation1971). The net loss of 18 species (22 extinct and 4 newly occurring species) reflected a significant change in the structure of avian community. The homogenization pattern evident is often mirrored in large-scale analyses, even on a continental scale, mostly due to habitat loss and climate change (van Turnhout et al. Citation2007, Newbold et al. Citation2018). In Dottendorf, the reduction of semi-natural, open habitats by about 40% is the most likely cause of the extinction of 12 out of the 15 originally existing, open land species. Our results reflect a general trend, since birds in open agricultural landscapes are the most threatened group of common birds in Europe (Donald et al. Citation2001, Citation2006, Heldbjerg et al. Citation2018), a process driven to a great extent by habitat homogenization (Benton et al. Citation2003).
In addition to the now mostly extinct species of open habitats, which largely occurred in low abundance during the earlier study (Wink Citation1971), we found a strong decrease in abundance for formerly common species that are typical of human settlements, such as House Sparrow, Common House Martin, and Common Swift, even though the area of settlements slightly increased in Dottendorf. This population trend agrees with the index of bird populations of settlements in Germany, which is lower than historical levels (Wahl et al. Citation2017). However, on large scales, a slight recovery of bird populations has been observed recently (Gerlach et al. Citation2019). Given that we only compare two points in time and do not have spatial replicates, such conclusion cannot be drawn based on our data.
The decreases of typical species of open land and settlement in Dottendorf are probably caused not only by changes in the area of land use types but also by habitat changes on a smaller scale. Species that nest in buildings, such as House Sparrow or Common Swift, lost breeding space due to modernization of buildings and the construction of new buildings that lack crevices and cavities (Shaw et al. Citation2008, Schaub et al. Citation2016). Further, some species that became extinct in our area depend on special structures, such as sandpits or wasteland, for feeding or breeding habitats, which underlines the importance of even small areas of habitats that add diversity to the landscape. Likely candidates include historically rare species, such as Common Whitethroat Sylvia communis, European Stonechat Saxicola rubicola, or Crested Lark Galerida cristata, as well as historically common and very common species such as Serin or Tree Sparrow (Bezzel Citation2001, Bauer et al. Citation2005a, Field et al. Citation2008, Sudmann et al. Citation2008, Lesiński Citation2009, Grüneberg et al. Citation2013, Gedeon et al. Citation2014). Additionally, urbanization has been related to altered predator–prey relationships (Jokimäki & Huhta Citation2000, Jokimäki et al. Citation2020). A higher predation rate by cats and corvids (Eurasian Magpies, Eurasian Jays, and Carrion Crow increased in abundance in Dottendorf) may have contributed to population declines in species like Common Blackbirds that do not hide their nests very well (Jokimäki et al. Citation2005). However, effects of corvids on the abundance of other species are generally rare (Madden et al. Citation2015) and there are no further hints that underline this speculation. Finally, the surprising declines of Common Blackbirds and Greenfinches might be explained by current outbreaks of diseases. Since 2011 the Usutu virus has been prevalent in western Germany and has killed a large proportion of Common Blackbirds (Bosch et al. Citation2012, Lühken et al. Citation2017), while Trichomonas gallinae, a protozoan, has been responsible for the deaths of numerous Greenfinches since 2009 (Peters et al. Citation2009).
In contrast to the decline in open land and settlement species, most forest species show a strong increase in abundance in Central Europe due to an increase in forest area, tree age, and the development of scrub cover offering more shelter, food, and breeding places (Bundesministerium für Ernährung und Landwirtschaft Citation2014, Citation2017). Partly, this is also reflected at our study site because seven out of the nine species with the strongest increase in population are forest species. But the strong increase in the abundance of forest species is not mirrored by the only marginal increase in forest area in Dottendorf. One plausible explanation for the increasing populations may be the increasing age of trees in gardens and parks leading to a local expansion of the breeding range. Further, species like the Common Woodpigeon or the Blackcap, formerly strict forest species, have increased their habitat tolerance and nowadays breed in settlements to a larger extent (Mattes & Freiling Citation2009, Grüneberg et al. Citation2013, Sakhvon & Kröver Citation2020). Additionally, seven out of the nine species with the strongest increase in abundance are short-distance migrants (Kamp et al. Citation2021). Some of them have recently changed their migration behaviour, migrating shorter distances now and thus arriving earlier in spring (Berthold et al. Citation1992, Tryjanowski et al. Citation2002, Citation2005, Fiedler Citation2003). This may be explained by milder winter temperatures in their new wintering grounds and higher spring temperatures in Central Europe. Since temperatures in Bonn have also increased (www.bonn.de) the development of the bird community in our study site can be regarded as typical for many Central European cities. In summary, the increase of these species – especially if we include the massive rise of the Common Woodpigeon – counterbalanced the decrease of other species groups leading to a slight increase in the total number of breeding pairs in Dottendorf, while the avian assemblage became more homogeneous.
Total biomass of breeding birds increased by 55% in Dottendorf, although this effect was not statistically significant. This might support conclusions that urbanization leads to an increase in biomass (Chace & Walsh Citation2006), possibly due to an increase in habitat quality. However, the increase in biomass was mainly due an increase in the Common Woodpigeon, which commonly breeds in the study site but might make long foraging flights to areas far beyond our study plot (Herkenrath Citation1989). Excluding this species from the analysis led to a decrease in biomass of 16.7%, compared to the bird assemblage of 1969, which gives a more meaningful picture of the situation in Dottendorf, and reflecting patterns observed elsewhere (Bauer et al. Citation2005b). One reason for the slight decrease may be that the entire north-western part of the study site was transformed into an area with modern office buildings that supports only a minimal population of highly adaptable bird species; unfortunately the proportion of this type of land use is increasing.
Conclusions
Our study covers a single study site in Bonn and necessarily compares only two points in time. Hence, we call for caution when transferring our results onto other areas. However, to a great extent, the described patterns are in line with results from other areas or shorter time scales (Bellebaum et al. Citation2009, Bauer et al. Citation2019). Yet current bird monitoring schemes often do not reach far enough back in time to cope with a shifting baseline syndrome (Pauly Citation1995, Weiss & Schulze-Hagen Citation2014). Therefore, we underline the need for similar studies and encourage the search for older surveys in order to document biotic change in urban areas over longer periods of time. In addition, our study points to potential changes in conservation strategies for urban areas. It is important to note that the often-promoted greening of cities (Mell et al. Citation2013) does not necessarily promote avian species richness; so far, local initiatives (Mutke et al. Citation2019) have been unable to halt the loss of urban avian diversity.
Ruderal sites and other open habitats offer both breeding habitats for the open land species and foraging sites for many other species. Therefore, their important role for urban conservation, in addition to urban forests and breeding opportunities on buildings, should not be underestimated. Finally, we stress the importance of species-specific reasons which, in sum, influence the structure of whole communities.
Supplemental Material
Download MS Word (36.5 KB)Acknowledgements
We are grateful to Prof. Dr Michael Wink who gave us valuable information about his bird counts in 1969. Johannes Kamp (Dachverband Deutscher Avifaunisten) kindly provided a table of species-specific traits. Further, we thank Kathrin Schidelko for an independent analysis of the number of Common Woodpigeon and Common Blackbird territories, the district government of Cologne (‘Bezirksregierung Köln’) and the City of Bonn for providing historical aerial photos and statistical data as well as Klaus Göhring for the current aerial photo. We are much obliged to two anonymous reviewers and Ian R. Hartley, whose contributions substantially improved this manuscript.
References
- Abrahamczyk, S., Wohlgemuth, T., Nobis, M., Nyffeler, R. & Kessler, M. 2020. Shifts in food plant abundance for flower-visiting insects between 1900 and 2017 in the canton of Zurich, Switzerland. Ecol. Appl. 30: e02138. DOI: 10.1002/eap.2138.
- Bauer, H.-G., Bezzel, E. & Fiedler, W. (ed) 2005a. Das Kompendium der Vögel Mitteleuropas. Aula-Verlag, Wiebelsheim.
- Bauer, H.-G., Peintinger, M., Heine, G. & Zeidler, U. 2005b. Veränderungen der Brutvogelbestände am Bodensee – Ergebnisse der halbquantitativen Gitterfeldkartierungen 1980, 1990 und 2000. Vogelwelt 126: 141–160.
- Bauer, H.-G., Heine, G., Schmitz, D., Segelbacher, G. & Werner, S. 2019. Starke Bestandsveränderungen der Brutvogelwelt des Bodenseegebietes – Ergebnisse aus vier flächendeckenden Brutvogelkartierungen in drei Jahrzehnten. Vogelwelt 139: 3–29.
- Bellebaum, J., Erfmann, M. & Zegula, T. 2009. Veränderungen der Avifauna in Schwelm 1992–2003 – ein Vergleich zweier Rasterkartierungen. Charadrius 45: 199–212.
- Benton, T.G., Vickery, J.A. & Wilson, J.D. 2003. Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol. Evol. 18: 182–188.
- Berthold, P., Helbig, A.J., Mohr, G. & Querner, U. 1992. Rapid microevolution of migratory behaviour in a wild bird species. Nature 360: 668–670.
- Bezzel, E. 1985. Birdlife in intensively used rural and urban environments. Ornis Fenn. 62: 90–95.
- Bezzel, E. 2001. Bleibt nur der Spatz in der Hand? Vögel in der Planungslandschaft 2000. J. Ornithol. 142: 160–171.
- Bibby, C.J., Burgess, N.D. & Hill, D.A. 1995. Methoden der Feldornithologie. Neumann, Radebeuel.
- Blair, R.B. 1996. Land use and avian species diversity along an urban gradient. Ecol. Appl. 6: 506–519.
- Bonier, F., Martin, P.R. & Wingfield, J.C. 2007. Urban birds have broader environmental tolerance. Biol. Lett. 3: 670–673.
- Bosch, S., Schmidt-Chanasit, J. & Fiedler, W. 2012. Das Usutu-Virus als Ursache von Massensterben bei Amseln Turdus merula und anderen Vogelarten in Europa: Erfahrungen aus fünf Ausbrüchen zwischen 2001 und 2011. Vogelwarte 50: 109–122.
- Bundesamt für Kartographie und Geodäsie (BKG, Hrsg.). 2020. TopPlusOpen 1:250.000 Europa. dl-de/by-2-0 (www.govdata.de/dl-de/by-2-0). https://gdz.bkg.bund.de/index.php/default/open-data/prasentationsgraphik-topplusopen-1-100-000-topplusopen-p100-1091.html. Download: 13.11.2020.
- Bundesministerium für Ernährung und Landwirtschaft. 2014. Der Wald in Deutschland – Ausgewählte Ergebnisse der dritten Bundeswaldinventur. BMEL, 52 pp.
- Bundesministerium für Ernährung und Landwirtschaft. 2017. Waldbericht der Bundesregierung 2017. BMEL, 288 pp.
- Bundesstadt Bonn – Statistikstelle. 2020a. Bevölkerungsstatistik, Stichtag 31.12.2019.
- Bundesstadt Bonn – Statistikstelle. 2020b. Bevölkerungsstatistik, Volkszählung 27. Mai 1970.
- Ceballos, G., Ehrlich, P.R., Barnosky, A.D., García, A., Pringle, R.M. & Palmer, T.M. 2015. Accelerated modern human–induced species losses: entering the sixth mass extinction. Sci. Adv. 1: e1400253.
- Chace, J.F. & Walsh, J.J. 2006. Urban effects on native avifauna: a review. Landsc. Urban Plann. 74: 46–69.
- Chamberlain, D.E., Cannon, A.R., Toms, M.P., Leech, D.I., Hatchwell, B.J. & Gaston, K.J. 2009. Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151: 1–18.
- Clergeau, P., Croci, S., Jokimäki, J., Kaisanlahti-Jokimäki, M.L. & Dinetti, M. 2006. Avifauna homogenisation by urbanisation: analysis at different European latitudes. Biol. Cons. 127: 336–344.
- Conole, L.E. & Kirkpatrick, J.B. 2011. Functional and spatial differentiation of urban bird assemblages at the landscape scale. Landsc. Urban Plann. 100: 11–23.
- Croci, S., Butet, A. & Clergeau, P. 2008. Does urbanization filter birds on the basis of their biological traits. Condor 110: 223–240.
- Devictor, V., Julliard, R., Couvet, D., Lee, A. & Jiguet, F. 2007. Functional homogenization effect of urbanization on bird communities. Conserv. Biol. 21: 741–751.
- Donald, P.F., Green, R.E. & Heath, M.F. 2001. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. Ser. B Biol. Sci. 268: 25–29.
- Donald, P.F., Sanderson, F.J., Burfield, I.J. & van Bommel, F.P.J. 2006. Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric. Ecosyst. Environ. 116: 189–196.
- Erz, W. 1966. Ecological principles in the urbanization of birds. Ostrich 37: 357–363.
- Eurostat (Hrsg.). 2020. Nomenclature des Unités territoriales statistiques (NUTS). https://ec.europa.eu/eurostat/de/web/gisco/geodata/reference-data/administrative-units-statistical-units/nuts. Download: 13.11.2020.
- Fidino, M. & Magle, S.B. 2017. Trends in long-term urban bird research. In Murgui, E. & Hedblom, M. (eds) Ecology and Conservation of Birds in Urban Environments, 161–184. Springer, Cham. DOI: 10.1007/978-3-319-43314-1_9.
- Fiedler, W. 2003. Recent changes in migratory behaviour of birds: a compilation of field observations and ringing data. In Berthold, P., Gwinner, E. & Sonnenschein, E. (eds) Avian Migration, 21–38. Springer, Berlin.
- Field, R.H., Anderson, G.Q.A. & Gruar, D.J. 2008. Land-use correlates of breeding performance and diet in Tree Sparrows Passer montanus. Bird Study 55: 280–289.
- Gedeon, K., Grüneberg, C., Mitschke, A., Sudfeldt, C., Eikhorst, W., Fischer, S., Flade, M., Frick, S., Geiersberger, I., Koop, B., Kramer, M., Krüger, T., Roth, N., Ryslavy, T., Stübing, S., Sudmann, S.R., Steffens, R., Vökler, F. & Witt, K. 2014. Atlas of German Breeding Birds. Bird Monitoring Foundation Germany, Münster.
- Geobasis NRW (Hrsg.) 2020. Digitale Verwaltungsgrenzen. https://www.bezreg-koeln.nrw.de/brk_internet/geobasis/verwaltungskarten/verwaltungsgrenzen/index.html. Download: 13.11.2020.
- Gerlach, B., Dröschmeister, R., Langgemach, T., Borkenhagen, K., Busch, M., Hauswirth, M., Heinicke, T., Kamp, J., Karthäuser, J., König, C., Markones, N., Prior, N., Trautmann, S., Wahl, J. & Sudfeldt, C. 2019. Vögel in Deutschland – Übersichten zur Bestandssituation. DDA, BfN, LAG VSW, Münster.
- Grüneberg, C., Sudmann, S.R., Weiss, J., Jöbges, M., König, H., Laske, V., Schmitz, M. & Skibbe, A. 2013. Die Brutvögel Nordrhein-Westfalens. NWO u. LANUV (Hrsg.), LWL-Museum für Naturkunde, Münster.
- Heldbjerg, H., Sunde, P. & Fox, A.D. 2018. Continuous population declines for specialist farmland birds 1,987–2,014 in Denmark indicates no halt in biodiversity loss in agricultural habitats. Bird Conserv. Int. 28: 278–292.
- Herkenrath, P. 1989. Ein Beitrag zur Nahrungsökologie städtischer Ringeltauben (Columba palumbus). Vogelwelt 110: 2–11.
- Ibáñez-Álamo, J.D., Rubio, E., Benedetti, Y. & Morelli, F. 2017. Global loss of avian evolutionary uniqueness in urban areas. Glob. Change. Biol. 23: 2990–2998.
- Jokimäki, J. & Huhta, E. 2000. Artificial nest predation and abundance of birds along an urban gradient. Condor 102: 838–847.
- Jokimäki, J. & Suhonen, J. 1993. Effects of urbanization on the breeding bird species richness in Finland: a biogeographical comparison. Ornis Fenn. 70: 71–77.
- Jokimäki, J., Kaisanlahti-Jokimäki, M.L., Sorace, A., Fernández-Juricic, E., Rodriguez-Prieto, I. & Jimenez, M.D. 2005. Evaluation of the “safe nesting zone” hypothesis across an urban gradient: a multi-scale study. Ecography 28: 59–70.
- Jokimäki, J., Suhonen, J., Benedetti, Y., Diaz, M., Kaisanlahti-Jokimäki, M.-L., Morelli, F., Pérez-Contreras, T., Rubio, E., Sprau, P., Tryjanowski, P. & Ibáñez-Àlamo, J.D. 2020. Land-sharing vs. land-sparing urban development modulate predator–prey interactions in Europe. Ecol. Appl. 30: e02049. DOI: 10.1002/eap.2049.
- Kamp, J., Frank, C., Trautmann, S., Busch, M., Dröschmeister, R., Flade, M., Gerlach, B., Karthäuser, J., Kunz, F., Mitschke, A., Schwarz, J. & Sudfeldt, C. 2021. Population trends of common breeding birds in Germany 1990–2018. J. Ornithol. 162: 1–15.
- Kelcey, G. & Rheinwald, G. 2005. Birds in European Cities. Ginster-Verlag, St. Katharinen.
- Lepczyk, C.A. & Warren, P.S. (eds) 2012. Urban bird ecology and conservation. In Studies in Avian Biology, Vol. 45, 325 pp. University of California Press, Berkeley.
- Lesiński, G. 2009. Breeding ecology and population decline of the Crested Lark Galerida cristata in Warsaw, Poland. Ornis Hungarica 17: 1–11.
- Lühken, R., Jöst, H., Cadar, D., Thomas, S.M., Bosch, S., Tannich, E., Becker, N., Ziegler, U., Lachmann, L. & Schmidt-Chanasit, J. 2017. Distribution of Usutu virus in Germany and its effect on breeding bird populations. Emerg. Infect. Dis. 23: 1994–2001.
- Madden, C.F., Arroyo, B. & Amar, A. 2015. A review of the impacts of corvids on bird productivity and abundance. Ibis 157: 1–16.
- Marzluff, J.M., Bowman, R. & Donnelly, R. 2001a. Avian Ecology in an Urbanizing World. Kluwer Academic Press, Boston, MA.
- Marzluff, J.M., Bowman, R. & Donnelly, R. 2001b. A historical perspective on urban bird research: trends, terms, and approaches. In Marzluff, J.M., Bowman, R. & Donnelly, R. (eds) Avian Ecology and Conservation in an Urbanizing World, 1–17. Springer, Boston, MA.
- Mattes, H. & Freiling, I. 2009. Der Bestand der Ringeltaube (Columba palumbus) in der Innenstadt von Münster/Westfalen 1989 und 2008. Osnabrücker Naturwissenschaftliche Mitteilungen 35: 223–228.
- McDonnell, M.J. & Pickett, S.T.A. 1990. Ecosystem structure and function along urban-rural gradients: an unexploited opportunity for ecology. Ecology 71: 1232–1237.
- McKinney, M.L. 2002. Urbanization, biodiversity, and conservation. Bioscience 52: 883–890.
- McKinney, M.L. & Lockwood, J.L. 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14: 450–453.
- Mell, I.C., Henneberry, J., Hehl-Lange, S. & Keskin, B. 2013. Promoting urban greening: valuing the development of green infrastructure investments in the urban core of Manchester, UK. Urban For. Urban Green. 12: 296–306.
- Morelli, F., Benedetti, Y., Ibáñez-Álamo, J.D., Jokimäki, J., Mänd, R., Tryjanowski, P. & Møller, A.P. 2016. Evidence of evolutionary homogenization of bird communities in urban environments across Europe. Global Ecol. Biogeogr. 25: 1284–1293.
- Murgui, E. & Hedblom, M. (eds) 2017. Ecology and Conservation of Birds in Urban Environments. Springer, Berlin.
- Mutke, J., Klement, J., Terlau, W., Freund, W. & Weigend, M. 2019. Die Natur der Region Bonn/Rhein-Sieg. Decheniana Beihefte 41: 1–167.
- Newbold, T., Hudson, L.N., Contu, S., Hill, S.L.L., Beck, J., Liu, Y., Meyer, C., Philips, H.R.R., Scharfemann, J.P.W. & Purvis, A. 2018. Widespread winners and narrow-ranged losers: land use homogenizes biodiversity in local assemblages worldwide. PLoS Biol. 16: e2006841.
- Nowakowski, J.J. 1996. Changes in the breeding avifauna of Olsztyn (NE Poland) 1968–1993. Acta. Orn. 31: 39–44.
- Olden, J.D. & Rooney, T.P. 2006. On defining and quantifying biotic homogenization. Glob. Ecol. Biogeogr. 15: 113–120.
- Pauly, D. 1995. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 10: 430.
- Pennington, D.N. & Blair, R.B. 2012. Using gradient analysis to uncover pattern and process in urban bird communities. In Lepczyk, C. A. & Warren, P. S. (eds) Urban Bird Ecology and Conservation. Studies in Avian Biology, Vol. 45, 9–32. University of California Press, Berkeley.
- Peters, M., Kilwinski, J., Reckling, D. & Henning, K. 2009. Gehäufte Todesfälle von wild lebenden Grünfinken an Futterstellen infolge Trichomonas-gallinae-Infektionen-ein aktuelles Problem in Norddeutschland. Kleintierpraxis 54: 433–438.
- R Development Core Team. 2017. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
- Rheinwald, G. 2005. Bonn. In Kelcey, G. & Rheinwald, G. (eds) Birds in European Cities, 41–54. Ginster-Verlag, St. Katharinen.
- Rheinwald, G. & Kneitz, S. 2002. Die Vögel zwischen Sieg, Ahr und Erft. Ginster-Verlag, St. Katharinen.
- Rheinwald, G., Wink, M. & Joachim, H.-E. 1984. Die Vögel in Großraum Bonn. Band 1: Singvögel. Beiträge zur Avifauna des Rheinlandes 22/23, Düsseldorf.
- Rheinwald, G., Wink, M. & Joachim, H.-E. 1987. Die Vögel in Großraum Bonn. Band 2: Nicht-Singvögel. Beiträge zur Avifauna des Rheinlandes 27/28, Düsseldorf.
- Sakhvon, V. & Kröver, L. 2020. Distribution and habitat preferences of the urban Woodpigeon (Columba palumbus) in the north-eastern breeding range in Belarus. Landsc. Urban Plann. 201: 103846. DOI:10.1016/j.landurbplan.2020.103846.
- Schaub, T., Meffert, P.J. & Kerth, G. 2016. Nest-boxes for Common Swifts Apus apus as compensatory measures in the context of building renovation: efficacy and predictors of occupancy. Bird Conserv. Int. 26: 164–176.
- Shaw, L.M., Chamberlain, D. & Evans, M. 2008. The House Sparrow Passer domesticus in urban areas: reviewing a possible link between post-decline distribution and human socioeconomic status. J. Ornithol. 149: 293–299.
- Sol, D., Trisos, C., Múrria, C., Jeliazkov, A., González-Lagos, C., Pigot, A.L., Ricotta, C., Swan, C.M., Tobias, J.A., & Pavoine, S. 2020. The worldwide impact of urbanisation on avian functional diversity. Ecol. Lett. 23: 962–972.
- Stadt Bonn (Hrsg.) 2020. Fläche der Ortsteile. https://opendata.bonn.de/dataset/fl%C3%A4chen-der-ortsteile. Download: 13.11.2020.
- Südbeck, P.H., Andretzke, H., Fischer, S., Gedeon, K., Schikore, T., Schröder, K. & Sudfeldt, C. (eds) 2005. Methodenstandards zur Erfassung der Brutvögel Deutschlands. Vogelwarte Radolfzell, Radolfzell.
- Sudmann, S.R., Grüneberg, C., Hegemann, A., Herhaus, F., Mölle, J., Nottmeyer-Linden, K., Schubert, W. & von Dewitz, W. 2008. Rote Liste der gefährdeten Brutvogelarten Nordrhein-Westfalens. Charadrius 44: 137–230.
- Suhonen, J., Jokimäki, J., Kaisanlahti-Jokimäki, M.L., Hakkarainen, H., Huhta, E., Inki, K. & Suorsa, P. 2009. Urbanization and stability of a bird community in winter. Ecoscience 16: 502–507.
- Tryjanowski, P., Kuźniak, S. & Sparks, T.H. 2002. Earlier arrival of some farmland migrants in western Poland. Ibis 144: 62–68.
- Tryjanowski, P., Kuźniak, S. & Sparks, T.H. 2005. What affects the magnitude of change in first arrival dates of migrant birds? J. Ornithol. 146: 200–205.
- van Turnhout, C.A.M., Foppen, R.P.B., Leuven, R.S., Siepel, H. & Esselink, H. 2007. Scale-dependent homogenization: changes in breeding bird diversity in the Netherlands over a 25-year period. Biol. Conserv. 134: 505–516.
- Wahl, J., König, C., Grüneberg, C. & Trautmann, S. 2014. Abschlussbericht zum Vorhaben “Entwicklung, Charakterisierung und Abstimmung von ökologischen Gruppen von Vogelarten”. Dachverband Deutscher Avifaunisten (DDA). Unpublished report.
- Wahl, J., Dröschmeister, R., König, C., Langgemach, T. & Sudfeldt, C. 2017. Vögel in Deutschland – Erfassung rastender Wasservögel. DDA, BfN, LAG VSW, Münster.
- Wei, J.W., Lee, B.P.Y.-H. & Wen, L.B. 2016. Citizen science and the urban ecology of birds and butterflies – a systematic review. PLoS ONE 11: e0156425.
- Weiss, J. & Schulze-Hagen, K. 2014. Viele Menschen, viele Vögel? Ornithologie und Vogelschutz in Nordrhein-Westfalen. Charadrius 50: 3–22.
- Wichmann, O. & Wichmann, K. 2020. Bonn-Dottendorf im Laufe der Jahre. Förderverein Stadtteilbibliothek Bonn-Dottendorf e.V., KultimO.
- Wink, M. 1971. Qualitative, quantitative und populationsdynamische Untersuchung des Brutvogelbestandes im Bonner Stadtteil Dottendorf. Charadrius 7: 57–63.