
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Capsule:
Compared to their rural counterparts, urban Northern Goshawks Accipiter gentilis potentially maximize their energy delivery to the nest by exploiting heavier prey species close to their maximum carrying capacity.
Aims:
We aimed to assess prey use of a raptor species with distinct reversed sexual size dimorphism from a perspective of physical limitations during foraging and the transportation of prey.
Methods:
We estimated the theoretical maximum load carrying capacity (MLCC) of fully developed Northern Goshawks based on their flight muscle mass. Additionally, we collected data on the breeding season diet of Northern Goshawks in urban and rural habitats in Germany. By linking MLCC estimates to the diet we explained the relevance of prey size from a viewpoint of load carrying capacity.
Results:
Estimates for the mean (± sd) additional portable loads were 684 g (± 237) for males and 971 g (± 235) for females, accounting for 96% and 84% of their body mass, respectively. Overall prey weight averaged higher for urban areas compared to rural ones, while the majority of prey items were between 200 and 500 g and below the estimated MLCC of both sexes, with the exception of single heavy species. Results suggest that prey use of Northern Goshawks during the breeding season is barely limited by prey transportability, since both sexes are physically capable of carrying the majority of prey species to their nest.
Conclusions:
Urban Northern Goshawks can exploit heavier, available prey species compared to their rural counterparts, allowing them to hunt larger prey closer to their MLCC. Ultimately, by maximizing their energy delivery to the nest, this presumably constitutes one key factor why Goshawks successfully colonized European cities. This study is the first to link estimates of physical limitations in prey transportation for a free-living raptor species to its prey composition in the light of colonizing urban environments.
Diet composition and prey choice in avian predators is a central and diversely addressed topic in the literature because the complex interplay between associated factors, such as prey abundance, prey vulnerability and transport costs, makes universal theories controversial in their applicability (Pyke Citation1984, Sih & Christensen Citation2001, Stephens & Krebs Citation1986). In general, theories predict that the choice of prey should be primarily based on its profitability, hence maximizing the energy provided by the prey per unit time, while taking any costs associated with the foraging process into account (Sih & Christensen Citation2001, Stephens & Krebs Citation1986). Such costs may come into play during locating, capturing, handling, and eventually carrying prey to a different location (Sih & Christensen Citation2001, Sinervo Citation1997). However, quantifying related costs remains challenging because isolating single factors is hardly possible.
The selection of prey is of vital importance to airborne central place foragers when maximizing the energy content delivered to the nest in order to cover increasing food demands of the mate and/or offspring (Fagerström et al. Citation1983, Kacelnik Citation1984, Orians & Pearson Citation1979). For the majority of raptor species, a large share of the prey items taken is transported to the nest during the chick-rearing period (Kenward Citation2006, Newton Citation1979, Walls & Kenward Citation2020). For these species, prey size is an important factor as the profitability of prey mainly depends on the interplay between the energy required for lifting and carrying, the distance to the nest and, hence, the associated costs of transportation (Sonerud Citation1992). Small prey items with energetic values below a certain threshold might not be transported at all (load-size effect: Sonerud Citation1992, Stephens & Krebs Citation1986) but will be directly consumed at the capture site (Rutz Citation2003, Sonerud Citation1992). Thus, biases in diet analysis due to selective transportation towards larger prey species might occur for single-prey loaders (Korpimäki et al. Citation1994, Sonerud Citation1992).
In general, selecting larger prey is beneficial when maximizing the rate of energy delivery to the nest, particularly as the travel distance from the nest increases (Orians & Pearson Citation1979, Stephens & Krebs Citation1986). However, physical limitations in carrying prey to the nest might restrict prey choice during the breeding season. Because reversed sexual size dimorphism is very common among raptors, prey size restrictions are expected to vary significantly between the sexes (Kenward Citation2006, Krüger Citation2005, Newton Citation1979, Opdam Citation1980). Additionally, limitations in prey size might be even more important for the smaller male, which captures most of the prey during incubation and early nestling period, while the female is predominantly sitting on the nest and protecting the brood (Kenward Citation2006, Newton Citation1979, Walls & Kenward Citation2020).
The maximum carrying capacity (the maximum mass with which an animal can still develop vertical lift) ultimately acts as an upper limit for prey choice in central place foragers, as the potential load additional to the predator’s own body mass is limited, and prey above a certain threshold will be too heavy to be transported to the nest (Stephen & Krebs Citation1986). Raptors in Europe feeding on large prey relative to their own body mass, have developed specific strategies to maximize the benefits of selecting these larger prey, especially when having to transport them over longer distances. Vultures, for example, do not carry the whole prey carcass or parts of it; instead they swallow only the most energy-rich parts of the carcass and carry these in their crop and gizzard to their young (von Blotzheim et al. Citation1989). In this way they avoid transporting less valuable but heavy parts, such as bones, skin, etc. Other raptors reduce the lifting of prey required as much as possible by building their nests below their hunting grounds. Golden Eagles living in Alpine regions of Europe, for example, hunt above the tree zone in open ranges and use gliding flight to carry the prey, with minimum energy expenditure, to the nest located beneath the hunting ground (Haller Citation1996, Pedrini & Sergio Citation2002).
To what extent a flying individual is able to transport additional load predominantly depends on the lift production generated by the wings and flight muscles (Marden Citation1987, Pennycuick et al. Citation1989). Lift production and take-off ability across insect and bird species have been shown to be well predicted by the mass of flight muscles, suggesting that high flight muscle mass allows steeper take-off angles and higher climbing rates (Hartman Citation1961, Marden Citation1987). In general, a high flight muscle ratio (flight muscle mass/ total body mass) was described as the single best factor explaining lift production, contradicting predictions that it should decrease with increasing body mass, or wing or disk loading (Marden Citation1987, Pennycuick Citation1968). High flight muscle ratios should particularly occur in animals that frequently need to lift loads with maximum power output, e.g. when transporting captured prey (Marden Citation1987). However, to our knowledge, no studies analysing prey choice and diet composition of raptor species have assessed the interplay between physiology and prey choice, or integrated physical limitations of prey transportation with respect to the maximum carrying capacity.
The Northern Goshawk Accipiter gentilis (hereafter Goshawk) is an avian top predator that is formerly described to inhabit wooded, undisturbed habitats in rural environments (Fischer Citation1995, Kenward Citation2006, Rutz et al. Citation2006). However, despite its high level of sensitivity to human disturbance, Goshawks have increasingly colonized urban environments in several European cities over recent decades, notably in Germany (Altenkamp Citation2002, Merling de Chapa et al. Citation2020, Rutz et al. Citation2006, Würfels Citation1999). While green metropoles such as Berlin, Cologne, and Hamburg offer sufficient breeding grounds in the form of parks and cemeteries, the year-round availability of profitable and abundant prey is most likely one of the key factors promoting the increasing colonization of urban environments by the species (Rutz et al. Citation2006). Thus, the diet of breeding Goshawks in European cities has been the topic of an increasing number of studies, especially in those German cities just mentioned (Altenkamp Citation2002, Altenkamp & Herold, Citation2001, Merling de Chapa et al. Citation2020, Rutz Citation2004, 2003, Würfels Citation1999). Throughout these studies several prey species, notably Feral Pigeon Columba livia, European Starling Sturnus vulgaris, Blackbird Turdus merula, and various corvids, repeatedly showed the highest proportions in diet, suggesting that Goshawks mainly prey on abundant species that are yet profitable and vulnerable to attacks (Rutz et al. Citation2006, Tornberg Citation1997). Rebollo et al. (Citation2017) further showed that prey preference of Goshawk populations in southern Europe was best explained by prey size, with the highest selectivity indices for prey of 100–400 g in body mass. The results suggest that small – to medium-sized prey species are most profitable for Goshawks as they provide the most energy per unit time, taking costs (locating, capturing, handling, transporting) according to the optimal diet theory into account (Sih & Christensen Citation2001).
In this paper we assess prey use of the Goshawk, a raptor species with distinct reversed sexual size dimorphism, from a perspective of physical limitations during foraging and transportation of prey. By estimating the theoretical maximum carrying capacity of fully developed Goshawks, based on their flight muscle mass, we seek to explain the relevance of prey size as a criterion for prey choice. Additionally, we linked such estimates to up-to-date data on the breeding season diet of both urban and rural Goshawk populations in Germany, in order to determine differences in prey composition between urban and rural populations from load carrying capacity perspective.
Methods
Flight muscle dissection
We determined the total flight muscle mass (TFMM) of eight fully developed Goshawks (four male, four female) found dead in Berlin between 2017 and 2020. For all eight birds, trauma was identified as the cause of death, with specific trauma due to collision for six individuals and unspecific trauma for the other two individuals. All birds therefore experienced a sudden death and were in good physical condition, based on measurements of fat tissues (subcutaneous, peritoneal cavity, coronary sulcus), shape of the chest muscles (pectoralis major), and the body mass. Carcasses were recovered within one day after death and were stored at −20 °C until examination. Birds were measured and weighed prior to muscle dissection. Age was determined according to the plumage colouration and sex was determined based on tarsus length and body weight (Kenward Citation2006). We did not differentiate between juvenile (< 1 year, n = 4) and adult (> 1 year, n = 4) individuals since all birds were fully developed. We dissected all flight muscles of the flight locomotor system according to Hartman (Citation1961) for one side of the body from medial to distal and weighed each muscle individually (online Table S1). TFMM was then determined as twice the cumulative mass of all muscles of the flight locomotor system ().
Figure 1. Flight muscles of the left-hand body side of a male Goshawk from a ventral (left) and a dorsal (right) view after skinning, prior to dissection.

Maximum carrying capacity
For each Goshawk we estimated the maximum carrying capacity (MCC) as the maximum possible load being lifted by an individual using an extended linear regression model (Equation 1), predicting the load-lifting ability of different bird, insect and bat species as an isometric function of flight muscle mass (Marden Citation1990). By integrating the findings of Pennycuick et al. (Citation1989) on additional load carrying in adult Harris’s Hawks Parabuteo unicinctus, Marden (Citation1990) showed that the estimated MCC of Harris’s Hawks (body mass approximately 920 g) is very consistent with previous measurements of the load-lifting ability of much smaller flying taxa (Marden Citation1987), hence expanding the strong isometric relationship between flight muscle mass and MCC across a broad range of taxa up to raptor species of the size of Goshawk. Although the weight of female Goshawks () slightly exceeded the range in body mass of the linear regression model, the consistency of predicted carrying capacity across flying species of various sizes gives us reason enough to tolerate extrapolation by such a small amount. We calculated the maximum portable load, hereafter the maximum load carrying capacity (MLCC) as the difference between an individual’s body mass and its MCC. This additional load represents the maximum prey mass at which Goshawks should still be able to create vertical lift during take-off.
(1)
(1)
Table 1. Different body measurements and parameters calculated for individual male and female Goshawks. TFMM = Total flight muscle mass; MCC = Maximum carrying capacity; MLCC = Maximum load carrying capacity.
Diet and prey mass analysis
We analysed data on the breeding season diet of Goshawks published by Merling de Chapa et al. (Citation2020); following which, prey items were collected in selected territories of three urban and three rural study sites during the breeding season (March until July 2016). Territories were located in the three German cities: Berlin, Hamburg and Cologne, as well as three rural ‘control’ sites: Brandenburg near Barnim, Schleswig-Holstein near Schleswig, and North Rhine-Westphalia near Kleve (Merling de Chapa et al. Citation2020). Prey remains were collected around nest sites following Rutz (Citation2003, Citation2004). Exact methods for collecting and identifying prey items are described in detail by Merling de Chapa et al. (Citation2020).
Breeding season diet was analysed with respect to prey size and mass. We assigned an average body mass for each bird prey species according to del Hoyo et al. (Citation1992), using the mean of listed body mass values for males and females. We further classified prey items of birds into nestlings, fledglings, and adult birds based on the growth stage of their feathers (Newton & Marquiss Citation1982), and assigned a mass according to their age. A mass of two-thirds of the adult weight was given to fledglings (Newton & Marquiss Citation1982, Opdam Citation1975, Rutz Citation2004), while for nestlings an estimated weight of one-third of the adult bird was assigned (Grønnesby & Nygård Citation2000, Rutz Citation2004). Although Goshawks pluck their prey by thoroughly removing feathers prior to transportation (Fischer Citation1995, Kenward Citation2006), we did not correct for a related reduction in prey mass, as feathers only account for a small proportion of a bird’s body mass.
For the majority of mammalian prey items, we could not determine the individual’s age accurately, so prey weights were adopted from Rutz (Citation2004). We adopted weight classes from Toyne (Citation2008) with minor adjustments.
Statistical analysis
Differences in mean prey mass between urban and rural habitats were tested using a non-parametric Mann–Whitney U test, as underlying data did not meet assumptions for parametric statistics. The significance level α was set at P < 0.05 for all statistical tests. The summarizing group values are presented as mean ± standard deviation (sd), if not stated otherwise. Data processing and statistical analyses were performed in R, version 3.5.1 (R Core Team Citation2018).
Results
Flight muscle mass and maximum load carrying capacity
The TFMM of Goshawks averaged 211.5 g (± 35.7) for males and 318.2 g (± 54.9) for females, accounting for 30% and 28% of the sexes’ body mass, respectively (). The MCC averaged 1399 g (± 239) for males and 2115 g (± 368) for females. A mean MLCC, representing the maximum additional load, of 684 g (± 237) was determined for male and 971 g (± 235) for female Goshawks, which on average covered 96% and 84% of their body mass, respectively. This additional load represents the maximum prey mass at which Goshawks should still be able to create vertical lift during take-off. The masses of dissected muscles for each individual Goshawk are provided in online Table S1.
Breeding season diet
Some 898 prey items were collected within 93 territories of the three urban (554 items) and three rural (344 items) study areas (Merling de Chapa et al. Citation2020). Breeding season diet predominantly consisted of bird species () of different sizes and taxa. In urban environments, medium-sized prey (240–320 g) such as the Feral Pigeon and larger prey (400–520 g), including Common Wood Pigeon Columba palumbus and Carrion Crow Corvus corone, comprised the largest share of the diet, together accounting for 70% and 80% in terms of counts and biomass, respectively (). Columbidae species made up significantly more of the diet in urban areas (65.4%, n = 355) compared to rural areas (35.7%, n = 122; Merling de Chapa et al. Citation2020). Further, prey between 400 and 520 g made up 15% more of the diet of urban Goshawks by means of biomass compared to absolute counts. In rural areas, prey was distributed more equally across weight classes, yet prey heavier than 240 g accounted for 74% of the diet with respect to biomass, but only 44% with respect to counts (). The heaviest prey species, including Mallard Anas platyrhynchos and Northern Raven Corvus corax, were almost exclusively recorded in rural areas, accounting for less than 5% in urban areas, both by means of biomass and counts.
Figure 2. Proportion of prey by weight classes by means of counts and biomass for the two different environments.
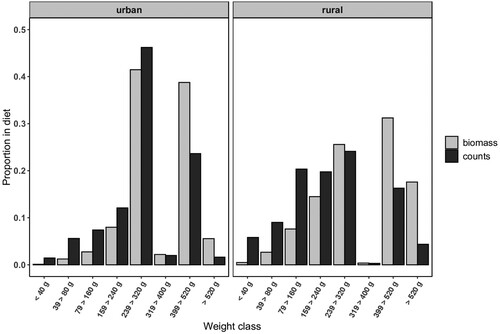
Table 2. Features of breeding season diet for Goshawks at urban and rural study sites.
In cities, juvenile prey (nestlings and fledglings combined) made up 10.9% of the diet by means of counts but only 6.6% by biomass. The proportion of juveniles was almost twice as high in rural areas by both counts (18.2%) and biomass (13.2%) compared to urban areas. Mammalian prey such as Rabbit Oryctolagus cuniculus, Brown Rat Rattus norvegicus, and different vole species made up between 2% and 4.3% of the diet of urban and rural Goshawks.
Mean prey mass significantly differed between habitat types (Mann–Whitney U test, W = 120760, df = 1, P < 0.001), with urban areas averaging higher prey mass (299 g ± 161 sd) than rural ones (255 g ± 221 sd). Among rural areas, however, the average prey mass at the Kleve study site (281 g ± 195 sd) was comparatively higher than at the other two sites, as medium-sized prey (200–300 g) made up a large share (42%, n = 87) of the diet (). In contrast, for the other two rural study sites, the majority of prey items weighed less than 200 g, accounting for approximately two-thirds of the diet (Brandenburg: 66%, n = 45; Schleswig: 63%, n = 43).
Figure 3. Density distribution of prey weights for all six study sites. Dashed (male goshawk, red area) and dotted (female goshawk, blue area) indicate mean maximum additional load, with coloured lines representing standard errors. Pictograms of exemplary prey species are positioned as a reference and according to their body weight.
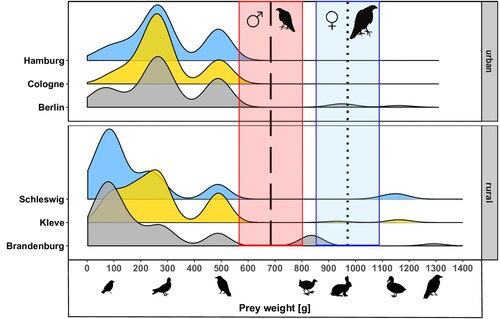
Compared to the MLCC of Goshawks, the majority of prey items in urban and rural areas lay beneath the maximum transportable prey mass of male and female Goshawks (). In cities, 98.4% (n = 545) and 99.5% (n = 551) of prey items weighed less than the MLCC of males and females, respectively. In the rural counterparts, prey weights of 95.6% (n = 329) and 97.7% (n = 336) of all items were below the respective MLCC of male and female Goshawks. Theoretically, portable prey weights for male Goshawks covered prey up to the size of Common Wood Pigeon and Carrion Crow, whereas larger prey, such as Eurasian Coot Fulica atra, Rabbit, and Mallard, lay above the estimated weight limit for males. The MLCC of female Goshawks covered a larger weight range, up to heavier prey such as Rabbit but not extending to Mallard duck and Northern Ravens.
Discussion
We determined the load carrying capacity of fully developed Goshawks based on their flight muscle mass, as a proxy towards limitations in prey use during the breeding season. Estimates for the MLCC for both sexes yielded transportable loads close to their respective average body mass. Despite the relatively small sample size of dissected individuals (n = 8; 4 males, 4 females), within-sex variation in TFMM/ body mass ratio was small and in line with flight muscle ratios reported for different song bird species and Harris’s Hawk (Hartman Citation1961, Marden Citation1987), suggesting that theoretical estimations were sufficiently reliable. Comparing the total flight muscle ratios of Goshawks with other raptor species remains difficult, however, as to our knowledge only Marden (Citation1990) reported comparable ratios of a flight muscle ratio of 22% for Harris’s Hawks, which were lower than the ratios of 28% (females) and 30% (males) reported in this study. Although the sample size of eight individuals was comparatively low, the body mass of all individuals fell within the range of mass reported for European Goshawks (male: 580-870 g, female: 880-1320 g, Kenward Citation2006) and was, in fact, very close to average weights reported for males (720 g) and females (1130 g).
The composition of breeding season diet of urban populations was consistent with findings of earlier studies, particularly with respect to the large proportion of pigeon species taken across urban populations, for which a detailed comparison of prey composition to the literature has already been reported by Merling de Chapa et al. (Citation2020). On average, urban Goshawks foraged on heavier prey compared to rural Goshawks. The mean prey weight of 299 g for urban populations was higher than in comparable studies, e.g. a mean weight of 247 g reported by Rutz (Citation2004) for the city of Hamburg. Only one rural site had higher mean prey weight than any urban site. We assume that the higher prey weight was mainly attributed to a higher availability of Feral Pigeons in the region. The higher average prey weight of urban Goshawks was also not the result of foraging on very heavy prey species, as these were almost exclusively foraged on in rural areas.
With respect to body mass, the majority of prey items of the different species lay below the MLCC of both male and female Goshawks, regardless of the habitat type. Rabbit and Eurasian Coot lay between the MLCC of both sexes; both species were too heavy to be transported by males but were within the MLCC of female Goshawks. Only few particularly heavy prey species exceeded the MLCC of both sexes, including Mallard and Northern Raven, which were almost exclusively foraged on by rural Goshawks. These findings have the potential to contradict our assumption that prey too heavy to be transported should be excluded from the breeding season diet. Potential explanations for the delivery of heavier prey could be based on the following factors: Firstly, aging of mammalian species such as Rabbit was not always possible, as only fur and single bones could be found, hence younger individuals could have been significantly lighter (Rutz Citation2004). Moreover, although plucking primarily improves the aerodynamics of a prey item, it might also reduce the weight, particularly for large bird species, which we could not account for when reflecting the process of prey transportation (Rutz Citation2003). Lastly, prey items too heavy to be transported to the nest by the adults could still be used for their own consumption at the place of capture in situations when opportunistic hunting of locally available prey allowed for it (Stephen & Krebs Citation1986). Information on prey items captured and eaten outside our sampling area would have been missed. However, although prey species heavier than our predicted MLCC have been reported regularly by studies on Goshawk breeding season diet, they accounted for extremely small proportions of the diet by means of quantity. We therefore consider them as highly opportunistically hunted prey species, that do contradict general patterns of prey use during the breeding season.
Overall, the results suggest that prey use of Goshawks during the breeding season is only marginally limited from a perspective of prey transportability, since both sexes are physically capable of carrying a broad range of prey species, representing the typical avifauna in both urban and rural environments, to their nest. Although we highlight that comprehensively assessing prey choice of raptors is complex and the foraging process shaped by several factors which we did not account for (particularly prey availability, but also encounter rate, prey handling, etc.), prey transportation still remains one of the major bottlenecks when foraging in order to feed the young (Sih & Christensen Citation2001, Sinervo Citation1997).
Because maximizing the energy content delivered to the central place is particularly important throughout the breeding period, prey selection should generally be adjusted accordingly (Fagerström et al. Citation1983, Kacelnik Citation1984, Orians & Pearson Citation1979), which is in line with the general theories of optimal foraging (Stephens & Krebs Citation1986). As a result, larger prey should be preferred if available, as it is generally associated with relatively smaller capture and transportation costs compared to smaller prey when taking energy output and expenditure into account (Andersson & Norberg, Citation1981, Korpimäki et al. Citation1994, Rebollo et al. Citation2017). While, in this context, the estimated MLCC constitutes the absolute maximum transportable load, the cost–benefit ratio might realistically be highest for prey species slightly below the MLCC. We therefore conclude from our findings that in urban environments Goshawks can exploit heavier, available prey species compared to their rural counterparts, which allows them to hunt larger prey closer to their MLCC in the first place. This is underlined by the higher percentage of pigeons hunted by urban Goshawks compared to rural individuals (Merling de Chapa et al. Citation2020). By primarily hunting prey which are closer to the MLCC, urban Goshawks can maximize their energy delivery to the nest when rearing their young. This particularly applies to the male, as it is almost solely responsible for prey intake until the female can temporarily leave the nest. In rural areas, on the contrary, Goshawks appear to mainly rely on smaller song bird species, much lighter than their potential MLCC, with very heavy prey constituting the exception.
Although we did not link data on breeding season diet with prey availability, the results are strongly in line with preference of Goshawks for a particular prey size range reported by Rebollo et al. (Citation2017) and preferential, opportunistic hunting of the most abundant and accessible prey of a given size (Kenward Citation2006, Merling de Chapa et al. Citation2020, Rutz et al. Citation2006). Although Goshawks show strong reversed sexual size dimorphism (Kenward Citation2006, Newton Citation1979), the MLCC of males was still much higher than the body mass of the majority of prey species detected in this study, as well as prey mass preferred by Goshawks in southwest Europe (Rebollo et al. Citation2017). Even the smaller male, which captures most of the prey during the incubation and early nestling periods, appears to be not majorly limited in potential prey species (Kenward Citation2006, Newton Citation1979, Walls & Kenward Citation2020). Reversed sexual size dimorphism is partially explained by the small-male hypothesis, following which males have evolved to be smaller for more efficient foraging and to reduce intersexual competition (Krüger Citation2005, Newton Citation1979). In general, smaller males have been associated with higher reproductive success (Pérez-Camacho et al. Citation2015). As assigning collected prey items specifically to one sex was not possible in our study, we could not determine whether the smaller male foraged on smaller prey, on average, than the female. We hypothesize, however, that in urban environments both members of the breeding pair can potentially exploit a broad range of prey species, while only locally available heavy prey, such as Rabbit and Mallard, are predominantly hunted by females, as they are too heavy for males to be transported to the nest.
Goshawk territory size is described to differ due to food availability (Kenward Citation2006). Particularly in cities, where transportation distances are presumably shorter due to locally available prey sources in the vicinity of the breeding location, preference for large prey is not necessarily a function of increasing distance to maximize energy output (Orians & Pearson Citation1979), but is also beneficial at short distances. In this context, territory sizes range between 500–5000 ha (Rutz Citation2006, Würfels Citation1994, Brüll Citation1953, Ziesemer Citation1983), but smaller territories have been reported for urban environments (Kenward Citation2006). Studies from Cologne and Hamburg describe home ranges from 500–1100 ha (Rutz Citation2006, Würfels Citation1994) whereby home ranges from non-urban breeders are described to range between 1500–5000 ha (Brüll Citation1953, Ziesemer Citation1983).
Despite being close to the MLCC, and abundant in urban environments, Carrion Crows comprised a large, yet smaller than expected proportion of the Goshawk diet. We assume that the social and aggressive defence behaviour generally displayed by corvids towards raptors makes them particularly difficult to hunt, resulting in higher risks or energy expenditure during foraging (Pérez-Camacho et al. Citation2015, Rebollo et al. Citation2017).
Ultimately, urban Goshawks appear to exploit heavier, available prey species compared to their rural counterparts, allowing them to hunt larger prey closer to their MLCC and therefore maximizing their energy delivery to the nest. Potentially, this constitutes one of the key factors that may have enabled the successful colonization of several European cities (Rutz et al. Citation2006), notably the German cities of Berlin, Hamburg, and Cologne (Altenkamp Citation2002, Rutz Citation2008, Würfels Citation1999). Being categorized as urban exploiters (Merling de Chapa et al. Citation2020), Goshawks appear to thrive off locally available and highly profitable prey, such as Feral Pigeon, with respect to their physiology.
To our knowledge, this study is the first to assess the physical capability of prey transportation for a free-living raptor species in a context of understanding prey use by linking emerging physical limitations to up-to-date data on breeding season diet. We highlight that several other factors, along with the foraging process, generally influence the choice of prey for aerial raptors and that future studies should focus on assessing these in order to illuminate foraging behaviour in its full complexity.
Supplemental Material
Download MS Word (17.8 KB)Acknowledgements
We specifically would like to thank Michael Lakermann, Gerard Müskens, Ronald Zollinger and Hans Wirth for collecting prey items. We also thank Katrin Koch (NABU Berlin e.V.) for reporting and passing on carcasses to the IZW.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Altenkamp, R. 2002. Bestandsentwicklung, Reproduktion und Brutbiologie einer urbanen Population des Habichts Accipiter gentilis (Linné 1758). Diploma Thesis, University of Berlin, Berlin.
- Altenkamp, R. & Herold, S. 2001. Habicht (Accipiter gentilis). In Arbeitsgemeinschaft Berlin-Brandenburgischer Ornithologen ABBO: Die Vogelwelt von Brandenburg Und Berlin, 175–179. Natur & Text, Rangsdorf.
- Andersson, M. & Norberg, R.A. 1981. Evolution of reversed sexual size dimorphism and role partitioning among predatory birds, with a size scaling of flight performance. Biol. J. Linn. Soc. 15: 105–130.
- Brüll, H. 1953. Der Greifvogel im Niederwildrevier. Merkblätter des Niederwildausschusses des DJV, No. 9. F.C. Meyer-Verlag, München.
- del Hoyo, J., Elliott, A. & Sagatal, J. 1992. Handbook of the Birds of the World. Lynx Edicions, Barcelona.
- Fagerström, T., Moreno, J. & Carlson, A. 1983. Load size and energy delivery in birds feeding nestlings: Constraints on and alternative strategies to energy-maximization. Oecologia 56: 93–98.
- Fischer, W. 1995. Die Habichte, 3rd edn. Neue Brehm-Bücherei, Magdeburg.
- Grønnesby, S. & Nygård, T. 2000. Using time-lapse video monitoring to study prey selection by breeding Goshawks Accipiter gentilis in Central Norway. Ornis Fenn. 77: 117–129.
- Haller, H. 1996. Langfristige Untersuchungen zur Populationsökologie von Aquila chrysaetos im Zentrum der Alpen. Ornithol. Beob, 9: 1–167.
- Hartman, F.A. 1961. Locomotor mechanisms of birds. Smilhson Misc Colitis 143: 1–91.
- Kacelnik, A. 1984. Central place foraging in starlings (Sturnus vulgaris). I. Patch residence time. J. Anim. Ecol. 53: 283–299.
- Kenward, R.E. 2006. The Goshawk. T & AD Poyser and Buteo Books, London.
- Korpimäki, E., Valkama, J. & Tolonen, P. 1994. Functional responses and load-size effect in central place foragers: data from the kestrel and some general comments. Oikos 69: 504–510.
- Krüger, O. 2005. The evolution of reversed sexual size dimorphism in hawks, falcons and owls: a comparative study. Evol. Ecol. 19: 467–486.
- Marden, J.H. 1990. Maximum load-lifting and induced power output of harris’ hawks are general functions of flight muscle mass. J. Exp. Biol. 149: 511–514.
- Marden, J.H. 1987. Maximum lift production during takeoff in flying animals. J. Exp. Biol. 130: 235–258.
- Merling de Chapa, M., Courtiol, A., Engler, M., Giese, L., Rutz, C., Lakermann, M., Müskens, G., van der Horst, Y., Zollinger, R., Wirth, H., Kenntner, N., Krüger, O., Chakarov, N., Müller, A.-K., Looft, V., Grünkorn, T., Hallau, A., Altenkamp, R. & Krone, O. 2020. Phantom of the forest or successful citizen? Analysing how Northern Goshawks (Accipiter gentilis) cope with the urban environment. R. Soc. Open Sci. 7: 201356. https://doi.org/10.1098/rsos.201356
- Newton, I. 1979. Population Ecology of Raptors. Poyser, Berkhamsted.
- Newton, I. & Marquiss, M. 1982. Food, predation and breeding season in Sparrowhawks (Accipiter nisus). J. Zool 197: 221–240.
- Opdam, P. 1980. Feeding Ecology and Niche Differentiation in Goshawk Accipiter gentilis L. and Sparrowhawk Accipiter nisus L. Radboud Universiteit Nijmegen (former Katholieke Universiteit Nijmegen), Nijmegen.
- Opdam, P. 1975. Intra- and interspecific differentiation with respect to the feeding ecology in two sympatric species of the genus Accipiter. Ardea 63: 30–54.
- Orians, G.H., Pearson, N.E., Horn, D.J., Mitchell, R.D. & Stairs, G.R. 1979. On the theory of central place foraging. In Horn, D.J., Mitchell, R.D. & Stairs, G.R. (ed) Analysis of Ecological Systems, 154–177. Ohio State Press, Columbus.
- Pedrini, P. & Sergio, F. 2002. Regional conservation priorities for a large predator: golden eagles (Aquila chrysaetos) in the Alpine range. Biol. Conserv. 103: 163–172.
- Pennycuick, C.J. 1968. A wind-tunnel study of gliding flight in the pigeon Columba livia. J. Exp. Biol. 49: 509–526.
- Pennycuick, C.J., Fuller, M.R. & McAllister, L. 1989. Climbing performance of Harris’ hawks (Parabuteo unicinctus) with added load: implications for muscle mechanics and for radiotracking. J. Exp. Biol. 142: 17–29.
- Pérez-Camacho, L., García-Salgado, G., Rebollo, S., Martínez-Hesterkamp, S. & Fernández-Pereira, J.M. 2015. Higher reproductive success of small males and greater recruitment of large females may explain strong reversed sexual dimorphism (RSD) in the northern goshawk. Oecologia 177: 379–387.
- Pyke, G.H. 1984. Optimal foraging theory: a critical review. Annu. Rev. Ecol. Syst 15: 523–575.
- R Core Team. 2018. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- Rebollo, S., García-Salgado, G., Pérez-Camacho, L., Martínez-Hesterkamp, S., Navarro, A. & Fernández-Pereira, J.-M. 2017. Prey preferences and recent changes in diet of a breeding population of the Northern Goshawk Accipiter gentilis in Southwestern Europe. Bird Study 64: 464–475.
- Rutz, C. 2008. The establishment of an urban bird population. J. Anim. Ecol. 77: 1008–1019.
- Rutz, C. 2006. Home range size, habitat use, activity patterns and hunting behaviour of urban-breeding Northern Goshawks Accipiter gentilis. Ardea 94: 185–202.
- Rutz, C. 2004. Breeding Season Diet of Northern Goshawks Accipiter gentilis in the city of Hamburg. Corax, Germany, 311–322.
- Rutz, C. 2003. Assessing the breeding season diet of goshawks Accipiter gentilis: biases of plucking analysis quantified by means of continuous radio-monitoring. J. Zool. 259: 209–217.
- Rutz, C., Bijlsma, R.G., Marquiss, M. & Kenward, R.E. 2006. Population limitation in the Northern Goshawk in Europe: a review with case studies. Stud Avian Biol. 31: 158–197.
- Sih, A. & Christensen, B. 2001. Optimal diet theory: when does it work, and when and why does it fail? Anim. Behav. 61: 379–390.
- Sinervo, B. 1997. Optimal foraging theory: constraints and cognitive processes. In Behavioural Ecology, 105–130. University of California, Santa Cruz.
- Sonerud, G.A. 1992. Functional responses of birds of prey: biases due to the load-size effect in central place foragers. Oikos 63: 223–232.
- Stephens, D.W. & Krebs, J.R. 1986. Foraging Theory, Monographs in Behavior and Ecology. Princeton University Press, Princeton, NJ.
- Tornberg, R. 1997. Prey selection of the Goshawk Accipiter gentilis during the breeding season: The role ofprey profitability and vulnerability. Ornis Fenn. 74: 15–28.
- Toyne, E.P. 2008. Breeding season diet of the Goshawk Accipiter gentilis in Wales. Ibis 140: 569–579.
- von Blotzheim, U.N., Bauer, K.M. & Bezzel, E. 1989. Handbuch der Vögel Mitteleuropas. AULA-Verlag GmbH, Wiesbaden.
- Walls, S. & Kenward, R. 2020. The Common Buzzard, Poyser Monographs. T & AD Poyser Ltd (A & C Black), London.
- Würfels, M. 1994. Entwicklung einer städtischen Population des Habichts (Accipiter gentilis) und die Rolle der Elster (Pica pica) im Nahrungsspektrum des Habichts – Ergebnisse vierjähriger Beobachtungen im Stadtgebiet von Köln. Charadrius 30: 82–93.
- Würfels, M. 1999. Ergebnisse weiterer Beobachtungen zur Populationsentwicklung des Habichts (Accipiter gentilis) im Stadtgebiet von Köln 1993–1998 und zur Rolle der Elster (Pica pica) im Nahrungsspektrum des Habichts. Charadius 35: 20–32.
- Ziesemer F. 1983. Untersuchungen zum Einfluß des Habichts (Accipiter gentilis) auf Populationen seiner Beutetiere. Beiträge zur Wildbiologie, Heft 2. PhD Thesis, University of Kiel, Kiel.