
ABSTRACT
Thermistors embedded on and within the nest wall were used to measure the changes in temperature between different parts of the architecture of the wall of Song Thrush nests as heat from a light bulb was conducted across the wall. The wood pulp forming the nest cup provided the greatest resistance to heat loss across the nest wall. Despite differences in nest mass and architecture, thermal conductance for Song Thrush nest walls were not statistically different from values previously reported for Common Blackbird Turdus merula nests. Song Thrush nests constructed over many years in the same geographical location exhibited an increase in the thickness of the wood pulp cup and a reduction in thermal conductance, i.e. the most recent nests were better insulated. However, nest mass or thermal insulation did not affect reproductive success between years.
The nest has a pivotal role in avian reproduction because it is where the clutch of eggs is laid and kept safe during contact incubation by the bird. In many species, the nest also has a key role during chick rearing (Deeming Citation2023). Contact incubation involves heat transfer between the bird and the eggs, so the energetics of incubation have interested physiologists for many years. However, the significance of energetics remains confusing because some species appear to expend very little extra energy during contact incubation, whereas in others incubation poses a considerable energetic burden (Tinbergen & Williams Citation2002, Nord & Williams Citation2015). Despite this ambiguity, the nest has long been considered to play a key role in minimizing the energetic expenditure of incubating birds, and so has been regularly studied over the past few decades (see reviews by Deeming & Mainwaring Citation2015, Deeming Citation2023). Some studies aimed to determine the thermal conductance of the nest wall (Heenan & Seymour Citation2011), whereas others have determined insulatory values using temperature loggers (Deeming et al. Citation2020). Whatever the technique, insulation seems to exhibit relatively similar effects among species constructing nests of differing sizes or construction techniques (Heenan & Seymour Citation2011, Deeming et al. Citation2020).
Insulatory values (°C·min−1) for thrush (Turdidae) nests were similar for the four species regularly breeding in the UK (Deeming et al. Citation2020), despite differences in nest mass. For instance, the nest of the Song Thrush Turdus philomelos is only 50–60% the mass of the nests of the Common Blackbird Turdus merula, Mistle Thrush Turdus viscivorus and Ring Ouzel Turdus torquatus (Biddle et al. Citation2018a). However, other studies showed that different parts of the wall structure affected insulation values of the nest. For Common Blackbird nests, removing the grass lining the cup and the outer wall of woody stems and moss increased cooling rates of heated temperature loggers inside the nest, which reduced the insulatory value of the nest to only 20% of the whole nest (Biddle et al. Citation2019). However, removal of the outer wall of the Song Thrush nest to leave the wood pulp layer, which constitutes the cup in this species, only reduced the insulatory value to 60% of the complete nest (Biddle et al. Citation2019). These differences were attributed to nest architecture; in the Common Blackbird removal of the grass lining in the cup exposed the temperature logger to the internal mud cup that had a different thermal conductance to grass. By contrast, for the Song Thrush, partial deconstruction of the nest did not affect the thermal conductance of the wood pulp nest cup. Externally, the Song Thrush nest is superficially like that of the Common Blackbird because they both have an outer wall consisting of comparable amounts of moss and woody stems (Biddle et al. Citation2018a). However, the internal mud cup and grass lining of the cup found in the Common Blackbird nests are replaced in the Song Thrush by a wood pulp cup that has a smooth, unlined internal surface (Biddle et al. Citation2018a, Biddle et al. Citation2018b). This meant that the temperature logger within the nest cup was in contact with wood pulp before and after partial deconstruction, and the impact on insulatory value was reduced.
Recently, Deeming & Jarvis (Citation2022) demonstrated that the internal architecture of the Common Blackbird’s nest affected the conduction of heat across the different elements of the nest wall in situ. Although the mud cup had the highest cooling gradient, values for thermal conductance were significantly affected by the thickness of the layer of grass lining the cup. Hence, if nest architecture affected insulation in this species what would be the effect of a different nest architecture? As far as is known, all European and North American thrushes use mud within the nest and line the cup with grass or roots (Harrison Citation1975, Citation1978), which suggests that they would have thermal characteristics like that of the Common Blackbird. However, the unique nest architecture and materials of the Song Thrush, and previous data from deconstructed nests (Biddle et al. Citation2019), would suggest that heat conduction across the nest wall would be quite different in this species.
The first aim of this study was to explore the factors that affect the thermal conductance of Song Thrush nests using the same methodology used for the Common Blackbird nests (Deeming & Jarvis Citation2022). It was predicted that the wall, which is of similar thickness in the two species (Biddle et al. Citation2018a), would have comparable values for thermal conductance, but that the different structural elements of the Song Thrush’s nest would affect how heat is conducted through the structure. For instance, conductance would correlate with cup thickness. Moreover, the Song Thrush nests studied were all collected in a small geographical area over a period of six years. This meant that the effect of time could be explored for a group of nests that were built in a similar kind of microclimate. It was predicted that year of construction would not affect thermal conductance. Finally, the effects of variation in thermal conductance on reproductive parameters, i.e. clutch size, hatchability, and fledging success, were explored to test the idea that better-insulated nests improved reproductive success.
Methods
Twenty-one Song Thrush nests were collected in west Devon (south-west England) by Mr. Rob Hubble over a period of six years. All but one of the nests were collected within a 2 km radius of Shaugh Prior, near Plymouth (centred on 50.4486, −4.0578 decimal latitude and longitude). The exception was a nest collected in 2019 at Buckland Monachorum (50.4949, −4.1239) some 7 km to the north-west of Shaugh Prior, but this was considered sufficiently close to be included in the analysis. Nests were available from 2017 (N = 3), 2018 (N = 4), 2019 (N = 5), 2021 (N = 5) and 2022 (N = 4). English COVID-19 restrictions prevented nest monitoring and collection, so nests were unavailable in 2020. Otherwise, nests were visited as part of the British Trust for Ornithology Nest Record Scheme and so the outcome of the breeding attempt was recorded for each nest. Data were available for date for the first egg, clutch size, and the number of hatched and fledged chicks. Nests were carefully collected after completion of the breeding season, were air-dried, and posted to Lincoln (eastern England) where they were stored dry in plastic bags within cardboard boxes, at a room temperature (∼18–20°C) prior to testing (as per Biddle et al. Citation2018a, Deeming & Jarvis Citation2022).
Prior to testing, each nest was first weighed (±0.1 g) on an electronic balance. The average total thickness of the wall was measured in mm using electronic callipers at 10 points around the top perimeter of the nest (Deeming & Jarvis Citation2022). The thickness of the outer nest wall was determined by pushing a wooden cocktail stick into the outer nest wall from the outside until it met the outside of the wood pulp cup. In each case, 10 measurements were taken around the nest and averaged. The thickness of the wood pulp cup wall was determined by subtracting the mean outer wall thickness from the mean wall thickness.
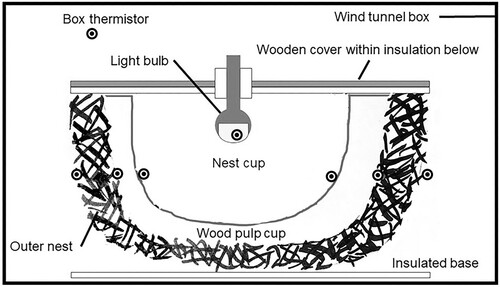
Nest insulation was measured using apparatus and a methodology as described by Deeming & Jarvis (Citation2022), which provides heat for the eggs so mimicking the presence of an incubating bird. Full details are described by Deeming & Jarvis (Citation2022) but are described briefly here (). A nest was placed within a purpose-built wind tunnel (100 × 50 × 50 cm, L x W x H) that minimized the effects of air movement on heat loss (Deeming & Humphreys Citation2019). Thermistors (RS Components, Corby, UK) were pushed into the nest wall, taped to wooden cocktail sticks that facilitated their positioning and fixing at key locations. These were located: (1) on the inside of the wood pulp cup, (2) external to the wood pulp cup and internal to the outer of moss and twigs, and (3) on the external surface of the outer nest layer. Two sets of the three thermistors were placed at either end of the long axis of each nest. Other thermistors monitored air temperature in the wind tunnel and the temperature of the bulb of a 3 W LED bulb of a flexible desk lamp. The plugs of each thermistor were attached to the outside of the wind tunnel for easy access. During testing the lamp was inserted through a hole filled with plastic insulation material in a wooden board that covered the nest surface. The underside of the board was lined with polystyrene that sealed the cup from the ambient air and insulated the top of the nest. Once the nest was prepared, and the bulb and the lid of the wind tunnel were in place, each thermistor was sequentially plugged into a digital thermometer (RS Components) and the temperature recorded in °C to one decimal place. Temperature readings were recorded every five minutes until 60 min had elapsed.
Figure 1. Diagrammatic illustration of the experimental set up showing the positioning of thermistors (black/white circles) within the nest wall (shown in section). Wires from each thermistor to the electronic thermometer are not shown. Modified from Deeming and Jarvis (Citation2022).
Data for analysis were taken for the full 60 min. For each nest, the two values for temperature recorded at equivalent thermistor positions within the wall were averaged. The difference in temperature between adjacent thermistors allowed calculation of the temperature difference created by the wood pulp cup and the outer wall of twigs, leaves and moss. Thermal conductance (mW·°C−1; Heenan & Seymour Citation2011) of the whole nest wall was calculated by dividing the heat flux from the bulb (3000 mW) by the mean temperature difference (°C) between the inside and outside surfaces of the nest.
Statistical analysis involved linear models (lm) in R (R Core Development Team Citation2021) that explored the relationships between variables, e.g. thermal conductance and nest mass. A two-sample t-test in R was used in comparison with published data for thermal conductance of Common Blackbird nests. Relationships between the temperature differences across each nest layer, and the width of the layer, were tested using Spearman rank correlation tests in R. Other analyses that compared the effects of the wall layers on variables involved either standard linear models or linear mixed-effects regression (lmer) models using the lmerTest (Kuznetsova et al. Citation2017) and lme4 (Bates et al. Citation2015) packages in R. Initial analysis tested the effect of the nest part on mean thicknesses, and for the mean temperature differentials, associated with each part. Further analysis determined the effects on the temperature differences recorded across the layers of wall thickness as a covariate and the part of the nest (i.e. wood pulp cup and outer wall), as a categorical factor, and an interaction term between thickness and nest part. Where the interaction term was not significant, the model was simplified. These lmer models included a nest ID number as a random factor to account for repeated measures from the nests.
Results
Thermal properties
Air temperature within the wind tunnel was 22–23°C and did not appreciably change during the experiments. The light bulb temperature often reached ∼50–55°C but on average the temperature on the inside surface of the wood pulp cup was 31.5°C (). There was a decrease of 6.9°C (SD = 1.8) for the temperature recorded on the external side of the wood pulp compared with a further decrease of 3.3°C (SD = 1.2) for the temperature recorded on the external surface of the outer nest. On average 67.8% (SD = 11.9) of the temperature difference across the wall is attributable to the wood pulp cup. These temperature differences were significantly different (lmer: F1,40 = 60.79, P < 0.0001). The thicknesses of the two structural layers were very different, with averages of 2.84 mm (SD = 1.85) and 29.67 mm (SD = 5.65) for the wood pulp cup and the outer nest wall, respectively (; lmer: F1,20 = 416.79, P < 0.0001).
Figure 2. Mean (±SD) values for temperature at different positions within the wall of Song Thrush nests. Slopes indicate the rate of change in temperature per cm of the respective wall structural components, i.e. the inner wood pulp cup and the outer nest wall. Position of the points on the x-axis indicates the mean (±SD) values for the thickness of the two different parts of the nest wall for the sample of 21 nests.
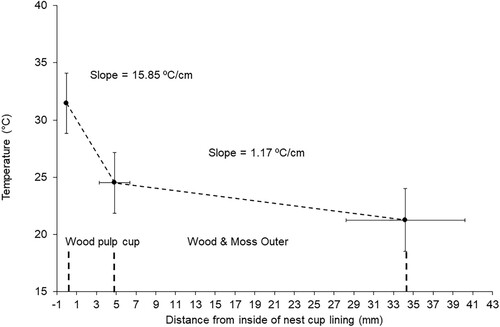
The linear mixed-effects model showed that there was no significant interaction on temperature difference between the layer thickness and the type of layer (F1,38 = 0.67, P = 0.418). In the simplified model, thickness as a covariate did not affect temperature difference (F1,39 = 0.44, P = 0.513) but nest wall layer as a fixed factor did significantly affect temperature difference (F1,39 = 9.49, P = 0.004). The temperature gradients (°C·cm−1) were also significantly different for the two parts of the nest wall (; lmer: F1,40 = 126.99, P < 0.0001). However, there were no significant relationships between temperature differences and the thickness of the wood pulp cup wall or the thickness of the outer nest (rho19 = 0.243, P = 0.289, and rho19 = 0.128, P = 0.580, respectively). There was also no significant relationship between the total thickness of the nest wall and the temperature difference recorded across the nest wall (rho11 = 0.185, P = 0.243).
The average thermal conductance of the nest wall was 303.3 (SD = 54.0) mW·°C−1 for nests that had mean mass of 79.8 (SD = 13.0) g. This value was not significantly different (two sample t-test: t = −0.38, DF = 16.68, P = 0.707) from the mean thermal conductance of 13 Common Blackbird nests of 292.0 mW·°C−1 (SD = 96.8) reported by Deeming & Jarvis (Citation2022). Thermal conductance was not significantly affected by either nest mass (F1,19 = 0.06, P = 0.813), wall thickness (F1,19 = 2.27, P = 0.149), outer wall thickness (F1,19 = 1.43, P = 0.246), or cup thickness (F1,19 = 2.05, P = 0.168).
Effect of time
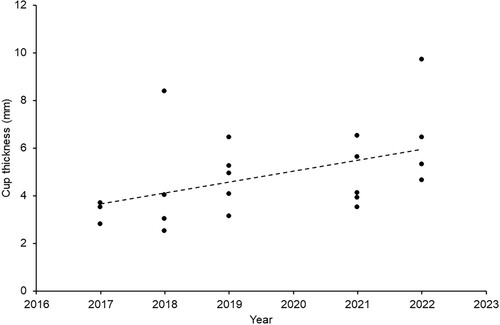
Linear modelling showed no significant effect of year on nest mass (lm: F1,19 = 2.73, P = 0.115), total wall thickness (lm: F1,19 = 0.01, P = 0.920), or outer wall thickness (lm: F1,19 = 0.27, P = 0.607). However, there was a significant effect of year on wood pulp cup thickness (lm: F1,19 = 4.67, P = 0.044, R² = 0.197), which doubled in thickness between 2017 and 2022 ().
Figure 3. Relationship between the thickness of the wood pulp cup and the year nests were collected. Trendline is generated by Excel to illustrate the pattern.
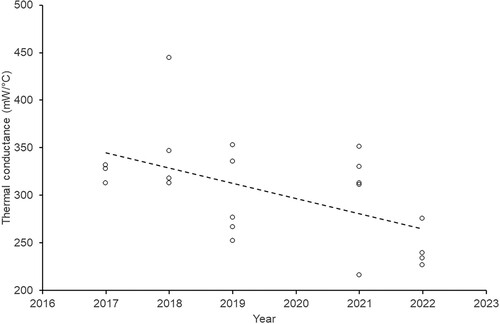
Thermal conductance had a significant negative relationship with year (; lm: F1,19 = 7.65, P = 0.012, R² = 0.287) but exhibited no significant effect with nest mass (lm: F1,19 = 0.06, P = 0.813), total wall thickness (lm: F1,19 = 2.27, P = 0.149), outer wall thickness (lm: F1,19 = 1.43, P = 0.246), or cup thickness (lm: F1,19 = 2.05, P = 0.168). The temperature gradient across the outer nest wall had no effect on conductance (lm: F1,19 = 0.48, P = 0.496) but had a significant negative relationship with the temperature gradient across the wood pulp cup (lm: F1,19 = 13.26, P = 0.002).
Figure 4. Relationship between the thermal conductance of the nest wall and the year nests were collected. Trendline is generated by Excel to illustrate the pattern.
Reproductive success
First egg day averaged ordinal day 116.0 (SD = 24.3) with a range of 71–154. Neither year nor thermal conductance significantly affected timing of breeding (lm: both P > 0.05). Clutch size averaged 3.7 eggs (SD = 0.8) but predation of 12 nests at the egg stage meant that only an average of 1.7 eggs (SD = 1.9) hatched. A further two nests were predated at the chick stage so an average of only 1.1 chicks (SD = 1.7) fledged. As for clutch initiation, neither year, date of clutch initiation, nor thermal conductance significantly affected breeding success (lm: all P > 0.05).
Discussion
Song Thrush nest architecture may differ from that of the Common Blackbird, but the thermal characteristics of the nest wall were similar in the two species. However, the use of wood pulp in the cup by the Song Thrush significantly affected the pattern of conduction of heat across the wall. Over time wood pulp cup thickness increased and thermal conductance decreased but there were no effects on reproductive success.
Previous studies of the insulation of Song Thrush and Common Blackbird nests have shown that insulatory values determined using temperature loggers were around 10% lower for Song Thrushes (Deeming et al. Citation2020). Using infrared thermography there were no temperature differences across intact and deconstructed walls for these two species (Biddle et al. Citation2018b). Biddle et al. (Citation2019) also found no species differences for insulatory values of intact nests determined using temperature loggers, but removal of outer nest materials and cup lining where present decreased considerably the insulation of Common Blackbird nests. Removal of the outer wall of Song Thrush nests reduced insulatory values with the wood pulp cup contributing 60% of the insulation provided by the whole wall. That the present study found that 68% of the temperature differences across the nest wall were attributable to the wood pulp cup suggests that this element of the nest wall is critical in determining thermal insulation in this species. Insulatory values are reflected in bird mass (Deeming et al. Citation2020), so similarities in thermal conductance between these two thrush species probably reflect their relatively similar sizes, but the slightly smaller Song Thrush requires slightly better insulation in the nest wall.
It was interesting that most physical characteristics of the Song Thrush nests were not affected by the year of construction, except for an increase in the thickness of the wood pulp cup and a decrease in the thermal conductance of the nest wall. It was not possible to ascertain what factors, e.g. climatic changes, were driving these changes over the six years of construction. However, Pied Flycatcher Ficedula hypoleuca nests constructed in any one year varied between widely spaced geographical locations (Briggs & Deeming Citation2016), and Common Redstart Phoenicurus phoenicurus nests varied between different habitats in the same vicinity (Briggs & Deeming Citation2021). Few studies have explored the effect of different nesting seasons on nest construction or function. Although there was no effect of year on size or composition of Eurasian Nuthatch Sitta europaea nests constructed in Lancashire over 10 years, thermal properties of the nest walls were not explored (Briggs & Deeming Citation2022). The present study is the first to demonstrate that nests built by a single species in the same geographical location can exhibit variation in composition and thermal characteristics over a long period of time. It is difficult to say whether the increase in the thickness of the wood pulp cup was a response by the birds to perceived need for better thermal insulation perhaps during colder weather. However, this study provides more evidence to demonstrate that nest construction is a plastic behaviour (i.e. not fixed, or stereotypical) that can respond to the circumstances encountered by the birds during nest building (see Deeming Citation2023). Further research is needed to collect nests from the same location for a wider range of species and over many years to see whether this result is unique.
Given that the nests of all other thrushes studied to date rely on mud in nest building (Biddle et al. Citation2018a), why have Song Thrushes adopted a different material for building the cup, and why is the cup not lined with grass? Song Thrushes seem to have evolved a method of nest construction that can produce levels of insulation comparable to that of other thrushes (see Deeming et al. Citation2020), but in a structure that weighs less than half the mass of the other nests (Biddle et al. Citation2018a). Give that nest construction will involve expenditure of time and energy (Mainwaring & Hartley Citation2013) and could be physiologically stressful (Moreno et al. Citation2008), Song Thrushes have evolved a method of nest construction that may improve their fitness by reducing the total effort of nest building. Wood pulp is possibly lighter to carry than mineral-based mud and not having to line the cup with grass or other plant material would also mean a considerable saving in time and energy. However, this energetic saving may have come at a cost because Song Thrushes rely on damp wood pulp collected from rotten wood. Absence of such material would limit the places where they can nest. Indeed, Tomialojc (Citation1992) suggested that the wider geographical distribution of the Song Thrush in Europe was limited by the availability of nest materials.
Whether nest construction has direct effects on reproductive fitness is unclear. Nest construction was seen as a stressful behaviour in Pied Flycatchers that negatively impacted female fitness, but was unrelated to breeding outcomes (Moreno et al. Citation2008). Experimentally supplementing Pied Flycatcher nests allowed females to have a higher provisioning rate leading to faster chick growth rates (Moreno et al. Citation2010). That thermal conductance did not affect timing, or the outcome, of breeding in the Song Thrush nests described here reflects previous studies where measures of thermal insulation or variation in nest composition have not correlated with reproductive outcomes (Schöll & Hille Citation2015, Briggs et al. Citation2019, Deeming et al. Citation2019, Briggs & Deeming Citation2021). This lack of an effect of nest construction could be interpreted as indicating that nest characteristics are of no consequence to the outcome of breeding. Alternatively, there may not be enough data to make solid conclusions.
However, variation in nest size and composition between birds and across years was observed here and seems to be typical of all species studied to date (Biddle et al. Citation2018a, Dickinson et al. Citation2022). The lack of effect may reflect birds building a nest that is pre-adapted to the prevailing environmental conditions at the time. These conditions change from year to year and so does nest composition and size. This implies that positive or negative effects of nest characteristics on reproductive output are not to be expected. By contrast, if nest architecture and composition were unchanging between years and individuals, adverse effects may be predicted. Hence, in some instances thermal characteristics of the wall may be ideal in some environments but there will be adverse effects in most years because the nest will not match the local environment that year. Further research is needed to explore the idea that variation in nest architecture, or thermal insulation, does not affect reproductive fitness in birds.
Acknowledgements
Many thanks go to Mr Rob Hubble for collecting and supplying the nests over such a long time. We are grateful to two anonymous reviewers for helping to improve a previous iteration of the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Bates, D., Mächler, M., Bolker, B. & Walker, S. 2015. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67: 1–48.
- Biddle, L.E., Broughton, R.E., Goodman, A.M. & Deeming, D.C. 2018a. Composition of bird nests is a species-specific characteristic. Avian Biol. Res. 11: 132–153.
- Biddle, L.E., Goodman, A.M. & Deeming, D.C. 2018b. Infrared thermography provides insight into the thermal properties of bird nests. J. Therm. Biol. 76: 95–100.
- Biddle, L.E., Goodman, A.M. & Deeming, D.C. 2019. Wall structure affects insulation of nests built by thrushes. Acta. Ornithol. 54: 255–262.
- Briggs, K.B., Biddle, L.E. & Deeming, D.C. 2019. Geographical location affects size and materials used in the construction of Pied Flycatcher (Ficedula hypoleuca) nests. Avian Res. 10: 17.
- Briggs, K.B. & Deeming, D.C. 2016. Use of materials in nest construction by Pied Flycatchers Ficedula hypoleuca reflects localised habitat and geographical location. Bird Study 63: 516–524.
- Briggs, K.B. & Deeming, D.C. 2021. Localised habitat affects size and materials used in the construction of Common Redstart (Phoenicurus phoenicurus) nests. Bird Study 68: 9–20.
- Briggs, K.B. & Deeming, D.C. 2022. Effects of year and box size on construction of Eurasian Nuthatch Sitta europaea nests. Ardea 110: 61–74.
- Deeming, D.C. 2023. A review of the roles materials play in determining functional properties of bird nests. Acta Ornithol. 58: 1–28.
- Deeming, D.C., Dickinson, A.M., Broughton, R.E., Locke, E., Gray, L.A., Bennett, S., Gilchrist, R., Muniz, S., Goodman, A.M. & Biddle, L.E. 2020. Factors affecting thermal insulation of songbird nests as measured using temperature loggers. Physiol. Biochem. Zool. 93: 488–504.
- Deeming, D.C. & Humphreys, E. 2019. Insulation of Common Chaffinch Fringilla coelebs nests is largely driven by animal-derived materials in the cup lining. Acta. Ornithol. 55: 253–260.
- Deeming, D.C. & Jarvis, R. 2022. Internal architecture differentially affects thermal insulation in the walls of Common Blackbird Turdus merula nests. Acta Ornithol. 57: 203–210.
- Deeming, D.C. & Mainwaring, M.C. 2015. Functional properties of nests. In Deeming, D.C. & Reynolds, S.J. (eds) Nests, Eggs, and Incubation: new Ideas About Avian Reproduction, 29–49. Oxford University Press, Oxford.
- Deeming, D.C., Morton, F.E.M. & Laverack, K.L. 2019. Nestbox size affects mass and proportions of materials used in Blue Tit Cyanistes caeruleus nests. Bird Study 66: 130–135.
- Dickinson, A.M., Locke, E., Gray, L.A., Bennett, S.L., Biddle, L.E., Goodman, A.M. & Deeming, D.C. 2022. Composition of nests constructed by species in the Motacillidae, Sylviidae and Prunellidae. Avian Biol. Res. 15: 21–33.
- Harrison, C. 1975. A Field Guide to the Nests, Eggs and Nestlings of British and European Birds. Collins, London, UK.
- Harrison, C. 1978. A Field Guide to the Nests, Eggs and Nestlings of North American Birds. Collins, London, UK.
- Heenan, C.B. & Seymour, R.S. 2011. Structural support, not insulation, is the primary driver for avian cup-shaped nest design. Proc. R. Soc. B 278: 2924–2929.
- Kuznetsova, A., Brockhoff, P.B. & Christensen, R.H.B. 2017. Lmertest package: tests in linear mixed effects models. J. Stat. Softw. 82: 1–26.
- Mainwaring, M.C. & Hartley, I.R. 2013. The energetic costs of nest building in birds. Avian Biol. Res. 6: 12–17.
- Moreno, J., Lobato, E., González-Braojos, S. & Ruiz-De Castañeda, R. 2010. Nest construction costs affect nestling growth: a field experiment in a cavity-nesting passerine. Acta Ornithol. 45: 139–145.
- Moreno, J., Martínez, J., Corral, C., Lobato, E., Merino, S., Morales, J., Martínez-de la Puente, J. & Tomás, G. 2008. Nest construction rate and stress in female Pied Flycatchers Ficedula hypoleuca. Acta Ornithol. 43: 57–64.
- Nord, A. & Williams, J.B. 2015. The energetic costs of incubation. In Deeming, D.C. & Reynolds, S.J. (eds) Nests, Eggs and Incubation: new Ideas About Avian Reproduction, 152–170. Oxford University Press, Oxford.
- R Core Development Team. 2021. R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
- Schöll, E.M. & Hille, S.M. 2015. Do Great Tits Parus major nesting at high altitudes build better insulated nests? Ardeola 61: 323–333.
- Tinbergen, J.L. & Williams, J.B. 2002. Energetics of incubation. In Deeming, D.C. (ed) Avian Incubation: Behaviour, Environment, and Evolution, 299–313. Oxford University Press, Oxford.
- Tomialojc, L. 1992. Colonisation of dry habitats by the Song Thrush Turdus philomelos: is the type of nest material an important nest constraint? Bull. Brit. Ornithol. Club 112: 27–34.