Abstract
1. Pullets were given a control diet or a diet supplemented with a non-starch-polysaccharide hydrolysing enzyme preparation (NSP-enzyme) from weeks 6 to 14 of age to induce differences in the viscosity of the small intestinal ingesta. Half of each feeding group (n = 25) was infected with 250 embryonated Ascaridia galli eggs at an age of 6 weeks.
2. At 14 weeks of age, before the pullets were slaughtered, a balance experiment was conducted, to sample ingesta for viscosity measurements. Also, tissue samples of jejunum and ileum were taken for morphometrical and histopathological studies.
3. An infection of pullets with A. galli reduced the viscosity of the jejunal ingesta at high initial levels of viscosity after feeding the NSP-enzyme unsupplemented diet.
4. The faecal A. galli egg output by the pullets expressed as eggs per g excrement (EpG) was significantly reduced in infected pullets given the NSP-enzyme supplemented diet. Also, the number and length of worms was less in these pullets.
5. The tunica muscularis of the jejunum was significantly thickened, this effect being more pronounced at a low intestinal viscosity.
6. NSP-enzyme addition resulted in an increased length of jejunal villus and was paralleled by a decrease in jejunal viscosity.
7. Histopathology of jejunal and ileal sections revealed no pathological alterations.
8. The apparent retention of organic matter was increased after enzyme addition while parasite infection exerted no effect.
Introduction
Gallinaceous poultry kept outdoors or in deep litter are likely to be exposed to endoparasites such as the roundworm Ascaridia galli since the infection cycle runs full circle when the parasite eggs are consumed with excreta or infected earth. A screening for the occurrence of various nematode species in hens kept in different production systems in Bavaria revealed the highest prevalence for Capillaria obsignata (28%), A. galli (18%) and Heterakis gallinarum (16%) in free range hens, while prevalence of infection with these nematodes in floor-kept hens varied between 5 and 7%. Hens kept in cages were virtually free of these nematodes (Zeller, Citation1990). Similarly, the prevalence of C. obsignata, A. galli and H. gallinarum infection in hens kept in Denmark amounted to 54, 64 and 72% for free ranging hens, and to 52, 42 and 19% for floor-kept hens, while only A. galli, with a low prevalence of 5%, was found in cage-kept hens (Permin et al., Citation1999).
A non-specific depression of growth performance might be more likely to indicate A. galli infection than other clinical signs (Gauly et al., Citation2001). Such a growth depression is thought to be the result mainly of a competition between the worms and the host for the nutrients of small intestinal ingesta. However, the magnitude of growth depression ranges from zero, or even growth stimulation, to significant reduction (e.g. Chubb and Wakelin, Citation1963). The reasons for these variations are not yet fully understood and might be influenced by nutrition as discussed in the literature (Chubb and Wakelin, Citation1963; Koski and Scott, Citation2001). Among nutritional factors, the dietary carbohydrate composition was shown to be a potential factor influencing endoparasitoses (Petkevicius, Citation2007).
It is well known that soluble non-starch-polysaccharides (NSP) exert anti-nutritive effects in growing poultry by increasing the viscosity of the small intestinal content and generally impairing digestion and nutrient absorption (Bedford and Schulze, Citation1998; Dänicke et al., Citation1999). It could be hypothesised that intestinal viscosity, as influenced by diet composition, affects the establishment of worms by altering the ecological environment, and/or that worms may themselves influence the intestinal viscosity. Both processes could therefore influence nutrient availability for the host and/or reproductive capacity of the worm in an interacting manner.
We tested this hypothesis by applying a complete 2 × 2 factorial design and examining pullets both A. galli infected and uninfected with low and high intestinal viscosity induced experimentally.
Experimental design and procedures
A total of 100 female one-day-old chicks (Lohmann LSL) were randomly distributed to 4 floor pens with 25 chicks per pen at the Relliehausen experimental station of the University of Göttingen. The temperature and lighting regimen were in accordance with the recommendations of the breeding company. Birds were vaccinated against Newcastle disease at d 1 and 21 of age. Commercial feed and water were provided for ad libitum consumption during the first 5 weeks of age. Pullets in two pens were then fed on a control diet. The remaining pullets were fed the same diet supplemented with a non-starch-polysaccharide hydrolysing enzyme preparation (NSP-enzyme) from weeks 6 to 14 of age to induce differences in the viscosity of the small intestinal ingesta. In order to increase the differences in intestinal viscosity due to NSP-enzyme addition, the basal diet was designed to contain components with higher concentrations of soluble NSP to ensure a relatively high viscosity in the groups receiving the unsupplemented diet ().
Table 1. Experimental design and composition of the experimental diets as fed (g/kg)
Half of each feeding group (25 pullets) was infected with embryonated A. galli eggs at an age of 6 weeks, i.e. one week after introducing the experimental diets. The eggs had previously been collected from the uteri of several females and incubated at 20°C in a 4% potassium bichromate solution for 14 d before being used for infection (Gauly et al., Citation2005). Pullets were orally infected with a suspension of 0.3 ml containing 250 embryonated A. galli eggs using a 6-cm cannula. Spot samples of excreta from each pullet were collected at week 12 (n = 25) and week 14 (n = 9) to count the excreted nematode eggs. Birds were weighed at the beginning of weeks 6, 7, 13 and 15. Based on these data the individual live weight gains were calculated for week 6 (one week before infection), for the period between weeks 7 and 12 (ad libitum feeding period) and for week 14 (balance period). As all pullets of the individual experimental groups were kept in one pen during the rearing period, the feed intake was not recorded.
The balance experiment was performed according to the total collection method as described in detail earlier (Schiemann, Citation1981). According to these guidelines, a minimum of three birds is required when complete diets are tested. However, in order to account for unknown uncertainties related to worm burden, a total of 9 birds were tested per group. Briefly, 9 pullets per group were transferred from the floor pens to individual balance cages and acclimatised during a 4-d pre-period. Then they were adjusted to a daily feed amount of 70 g /pullet in the following 5-d period. This feed quantity was maintained during the following 5-d collection period. Excreta were collected from the plastic trays beneath the cages in the morning and in the afternoon, and were temporarily frozen at −20°C. Excreta were then freeze-dried and ground to pass through a 1-mm screen.
In the course of the balance experiment, the pullets were conditioned to eat their morning meal within a period of less than 10 min. On the last day of the balance experiment, all 36 pullets were killed exactly 3 h after feeding by cervical dislocation followed by cutting the neck vessels. Jejunum (end of duodenum to Meckel's diverticulum), ileum (end of jejunum to approximately 2 cm to the ileo-caecal junction) were quickly dissected and ligated. Two cross-sections of each intestinal segment were further processed for histological examinations.
The contents of the respective segments were stripped out, pooled for three animals to give three samples per treatment, weighed and frozen for further analysis. During collection, ingesta worms were separated, counted and length was measured.
Analyses
Egg counting
A modified McMaster technique (MAFF, 1986) was used for egg counting followed by use of a saturated sodium chloride solution and the MSD counting chamber. The faecal A. galli egg output was expressed as eggs per g excrement (EpG).
Dry matter and crude nutrients
Dry matter, crude ash, crude fat and Kjeldahl N of the diets and freeze dried excreta were analysed according to the methods of the VDLUFA (Naumann and Bassler, Citation1993). Crude protein of the diets was obtained by multiplying the Kjeldahl N by 6.25.
Viscosity
Viscosity of jejunal and ileal ingesta supernatants was determined by using a Brookfield viscometer as described earlier (Dusel et al., Citation1998).
Histology
Tissue samples of jejunum and ileum were fixed in a 10% formaldehyde solution buffered with CaCO3 for at least 24 h and embedded in paraffin. Subsequently, tissues were sectioned at a thickness of 5 µm and stained with haematoxylin and eosin (H&E) for histopathological and histomorphometrical examination. For histopathology, the individual tissue sections (n = 9) were visually evaluated for pathological lesions and graded into categories as follows: 0 = no signs, 1 = low degree and 2 = medium degree. Each particular diagnosis was evaluated by analysing contingency tables based on the comparison between absolute frequencies and the expected frequencies for the particular classes.
Histomorphometric measurements were carried out by light microscopy (Eclipse E200-F-microscope coupled to digital camera DS-5M, imaging software NIS-elements BR; Nikon GmbH, Düsseldorf, Germany). The villus length and thickness of the tunica muscularis were measured for each of the two cross-sections of jejunum and ileum of individual hens by using computer aided image analysis. Values were expressed as mean villus length and mean thickness of the tunica muscularis.
Statistics
Analysis of variance (ANOVA)
All parameters except the histopathological data were analysed according to a complete 2 × 2 factorial design of ANOVA according to the following model:
All statistics were carried out using the Statistica for the Windows™ operating system (StatSoft Citation1995).
Results
Performance
There was a slight trend towards improved live weight gain (P = 0⋅082) due to feeding the NSP-enzyme supplemented diet for one week before the half of the pullets were infected with A. galli (). This effect became significant during the next 5 weeks and occurred at a significantly lower level in nematode-infected pullets (−4%). No treatment effects on live weight gain were detected during the balance period where all pullets were fed the same amount of feed restrictively ().
Table 2. Body weight gain, dry matter content of excreta and apparent nutrient retention of pullets infected or not infected with A. galli (week 6 of age) and fed on diets either unsupplemented or supplemented with a non-starch-polysaccharide cleaving enzyme preparation from weeks 6 to 14 of age (end of week 14 of age, n = 9)
Apparent nutrient retention
The apparent retention of the organic matter was significantly enhanced by 2% NSP-enzyme addition independent of nematode infection (). Although similar relationships were observed for the apparent N-retention, the enzyme related trend for improvement failed to reach significance although the mean increase amounted to 6%. Apparent retention of crude fat and crude ash were not influenced by the treatments and varied between 68⋅5 and 70⋅5%, and between 26⋅7 and 30⋅6%, respectively.
Dry matter of excreta and viscosity of small intestinal ingesta
The supplementation of the diet with the NSP-enzyme resulted in an increase of the dry matter content of the excreta (). However, this effect occurred only in the parasite infected pullets as indicated by the significant interactions between infection and NSP-enzyme addition (P = 0⋅04). Viscosity of the supernatants of jejunal and ileal ingesta was reduced by approximately 60 and 75% due to NSP-enzyme supplementation, respectively (). This viscosity-reducing effect of the NSP-hydrolysing enzyme was on the same order of magnitude in both non-infected and parasite-infected pullets although it generally occurred at a lower level for the latter (−32 and −24% for jejunal and ileal ingesta, respectively).
Table 3. Viscosity of intestinal ingesta (n = 3), empty weight and histomorphometry of small intestine of pullets infected or not infected with Ascaridia galli (week 6 of age) and fed diets either unsupplemented or supplemented with a non-starch-polysaccharide cleaving enzyme preparation from week 6 to 14 of age (end of week 14 of age, n = 9)
Small intestinal morphology and histometry
The empty weight of the ileum was significantly reduced by −11 and −6% in non-infected and infected pullets, respectively, due to NSP-enzyme addition and was generally increased in infected pullets by approximately 14% ().
The villi of the jejunum were significantly longer in pullets fed the NSP-enzyme supplemented diets, where the increase averaged approximately 43% (). In contrast, no treatment effects on villus length were detected in the ileum.
The tunica muscularis of both intestinal segments was significantly thickened due to parasite infection (). NSP-enzyme addition thickened the tunica muscularis of the jejunum significantly with a similar trend in the ileum. However, the enzyme effect was most obvious in the jejunum of infected pullets, as indicated by the significant interactions between infection and NSP-enzyme addition.
Histopathology
Generally, only minor differences were observed in the histopathological diagnoses between groups evaluated for various structures of jejunal and ileal tissues. A higher degree of accumulation of pigment-containing macrophages was detected in the lamina propria mucosae of the jejunum of uninfected pullets fed the unsupplemented diet (; P = 0⋅004). No pathological lesions were seen in the lamina epithelialis, the gut associated lymphoid tissue (GALT), or the tunica muscularis of this segment. In the ileum there was a trend toward increased incidence of crypt abscesses in the lamina epithelialis of pullets fed the NSP-enzyme supplemented diets independent of infection (; P = 0⋅099). Moreover, uninfected pullets fed the unsupplemented diet appeared to have a higher degree of lymphoplasmacytic infiltration of the lamina propria mucosae than the other groups (P = 0⋅052). Other ileal structures remained unaltered.
Table 4. Absolute frequencies of histopathological diagnosis of jejunal tissue (contingency tables) of pullets infected or not infected with Ascaridia galli (week 6 of age) and fedon diets either unsupplemented or supplemented with a non-starch-polysaccharide cleaving enzyme preparation from week 6 to 14 of age (end of week 14 of age, n = 9)
Table 5. Absolute frequencies of histopathological diagnosis of ileal tissue (contingency tables) of pullets infected or not infected with A. galli (week 6 of age) and fed on diets either supplemented or unsupplemented with a non-starch-polysaccharide cleaving enzyme preparation from week 6 to 14 of age (end of week 14 of age, n = 9)
Worm burden and EpG
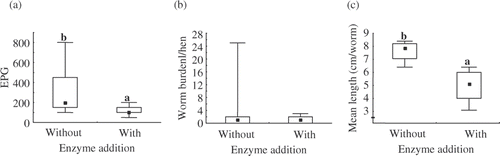
Faecal egg counts expressed as eggs per g excrement (EpG), and its variation, were significantly reduced by supplementing the diet of the infected pullets with the NSP-enzyme. All pullets tested in the balance experiment (n = 9) were EpG-positive at the outset, while only one pullet of each infected group still excreted eggs at the end of the balance trial. When the experiment was terminated at Week 14, adult worms were recovered in 5 out of the 9 pullets in each of the infected groups initially EpG-positive in Week 12. Although the numbers of worms did not significantly differ between groups, it appeared that pullets given the unsupplemented diet harboured more worms, as pullets with the highest and second highest number of worms (25 and 8 worms, respectively) were found in this group. The worm burden of these two pullets corresponded well to the EpG (). The percentage of female worms was not significantly influenced by NSP-enzyme addition and amounted to 61% on average. With enzyme supplementation, the mean worm length was significantly reduced by approximately 38% ().
Figure. Faecal egg output expressed as eggs per gram excrement (EPG) (a), worm burden (b) and mean worm length (c) as influenced by enzyme addition (without, no enzyme added; with, 100 FXU/kg diet); squares represent medians, boxes the 25 and 75 percentile and whiskers the minimum and maximum values; ab, different superscripts mark significant differences (P < 0⋅05)
Discussion
Worm burden and epidemiological aspects
Alternative housing systems with close animal contact to excreta, litter or soil facilitate A. galli infection and result in a higher prevalence of infection (Zeller, Citation1990; Permin et al., Citation1999; Chadfield et al., Citation2001). Rearing pullets, which do not require a high dietary energy concentration, are often fed with diets containing components low in ME concentration and high in fibre content. Bran from various cereal grains not only contains a higher total fibre content–which can be analysed as NSP–but also higher proportions of soluble NSP which might induce an increased viscosity of small intestinal ingesta.
The physiological consequences of high intestinal viscosity with its anti-nutritive effects, the modes of action of enzyme preparations capable of hydrolysing soluble NSP, and of the effects of the subsequent decrease in intestinal viscosity have been extensively studied (Dänicke, Citation1999). The interactions between intestinal viscosity and A. galli infection are not only interesting with regard to host nutrition and viscosity itself, but also from an epidemiological point of view.
Adult worms could be recovered at the end of the balance experiment in 5 out of 9 pullets of each of the infected groups, although all pullets were egg positive before being transferred to the balance cages. This means that worms were eliminated in 4 out of 9 pullets during the balance experiment independent of NSP-enzyme supplementation or intestinal viscosity. Whether the transfer of the pullets from floor pens to individual balance cages and/or the restricted feeding regimen applied in the balance experiment caused stress for the nematodes, and possibly contributed to their elimination, can not be explained by the present results and needs to be examined further. In this context, there are hints of both weak age resistance and a self-cure phenomenon to an A. galli infection (Permin and Ranvig, Citation2001). The latter might be influenced by nutrition or feeding regimen (ad libitum vs. restrictive). Moreover, a genetic component is discussed in the expulsion capacity of adult worms (Permin and Ranvig, Citation2001; Gauly et al., Citation2002).
Worm eggs were found in the excreta of only one of the 5 worm-harbouring pullets of each infected group. This might be explained by the low total reproductive capacity of the worm burden which amounted, in some cases, only to one or two female worms in individual pullets.
With regard to treatment effects it is evident that the worm number and the mean worm length were higher in pullets with higher intestinal viscosity due to feeding the NSP-unsupplemented diet. Together with the significantly higher mean EpG, as measured before the pullets were transferred to the balance cages, it might be assumed that high feeding diets inducing a high intestinal viscosity would favour the establishment of the nematodes and a more effective spreading of oocysts to the environment. That these effects were indeed closely related to intestinal viscosity can be deduced by the corresponding decrease in number and length of the worms, and the lower EpG, due to supplementation of the diet with an exogenous enzyme preparation well known to decrease intestinal viscosity via a partial hydrolysis of soluble NSP.
From an epidemiological point of view, not only the higher EpG under the conditions of a higher intestinal viscosity is of interest, but also the lower dry matter content and the higher viscosity of excreta, which is in general agreement with the literature (Bedford and Schulze, Citation1998). Both higher excreta viscosity and moisture content might significantly contribute to the spreading of eggs, since the feathers of the birds concerned are contaminated with excreta (Dänicke et al., Citation2007). Wet litter facilitates external parasite development (Davies and Joyner, Citation1955) and increases the tenacity of eggs in the environment (Roesicke and Greuel, Citation1992).
Growth performance, viscosity, nutrient retention, gut morphology and pathology
The artificial infection of pullets with embryonated eggs of A. galli resulted in a significant depression of growth performance when compared to the uninfected control group. The NSP-enzyme addition to the diet counteracted this detrimental effect to some extent, but failed to reach the live weight-gain improvement level of the uninfected group given the NSP-enzyme supplemented diet. These enzyme effects were closely related to the enzyme-induced decrease in intestinal viscosity, and the concomitant increase in the apparent retention of organic matter with a similar trend to nitrogen retention. These increases in the apparent nutrient retention are probably based on an improved nutrient digestibility and absorption and find their morphological equivalent in the increased length of jejunal villi, which favour absorption by an increase of the absorptive surface as suggested by Langhout et al. (Citation1999).
However, when discussing the effects of intestinal viscosity in relation to nutrient retention, it needs to be stressed that enzyme effects on intestinal viscosity generally occurred at a significantly lower level for infected birds. Thus, although the parasite infection itself also decreased viscosity, this additional reduction was insufficient to reach the performance levels of their uninfected counterparts. This would suggest that other adverse parasite effects prevented the positive effects on live weight gain which would be expected from the lower intestinal viscosity. Moreover, the parasite-related extra effect on decreased viscosity seemed to be independent of nutrient retention and jejunal villus length. Interestingly, the relative weight of the emptied ileum was decreased by NSP-enzyme addition, but at a significantly higher level when birds were infected. Moreover the thickness of the tunica muscularis of both small intestinal segments was increased in infected pullets. It appeared that the thickness was further increased when pullets were fed the diet with added NSP-enzyme, as indicated by the significant interactions between infection and NSP-enzyme supplementation. Neglecting these interactions for a moment, it appeared obvious that the parasite-related increase of the ileal weight was mainly caused by the increased thickness of the tunica muscularis, while the effect of the NSP-enzyme addition on the reduction of the weight of the small intestine was not reflected by the thickness of the tunica muscularis. Additional treatment effects on the length of small intestinal segments which were not recorded might be responsible for the unexplained variation observed. Histomorphological viscosity-related alterations included an increased number of goblet cells per 100 enterocytes (Langhout et al., Citation1999) while physiologically, an increased thickness of the unstirred water layer adhering to the mucosa and an enhanced protein synthesis of mucosal cells could be demonstrated (Dänicke et al., Citation2000). These processes may have increased the empty weight of small intestinal tissues by the more-or-less water soluble layer sticking to the mucosa.
On the other hand, the parasite effects on the increased empty weights of small intestines can be explained by the increased thickness of the tunica muscularis, which might be interpreted as a reactive hypertrophy to the presence of the parasite. This reaction occurred independently of the presence of worms in the intestine as determined at the end of the experiment. Thus, the hypertrophy of the tunica muscularis might indicate chronic effects of this endoparasitosis. The histopathological examinations did not reveal lesions due to parasite invasion, which could be expected during the histotropic period of A. galli. However, it needs to be considered that the histotropic phase typically occurs in the period between about one and 3 weeks post infectionem (Tugwell and Ackert, Citation1952), but pullets were slaughtered 8 weeks after infection. It has been shown that the duration of the histotropic phase strongly depends on the infection dose and might be influenced by the interactions between host immunity and parasite-arrested development (Herd and McNaught, Citation1975). Thus tissue damage probably occurred some weeks before jejunal and ileal cross-sections were prepared for histological examinations. Diphteric or fibrinoid inflammation of the small intestine, or tissue focussing of inflammation cells like macrophages or neutrophilic granulocytes as reported for turkeys (Hemsley, Citation1971) infected with Ascaridia spp., could not be observed in the present experiment. This might additionally support the view that acute tissue lesions due to parasite infection were not present. Whether or not an earlier histotropic period and/or the presence of adult worms in the intestinal lumen stimulated the increase in tunica muscularis thickness can not be explained by the present experiment and this needs to be investigated in future studies.
In conclusion, when discussing the results from the present experiment, it should be kept in mind that the transfer of the infected birds from floor pens to the individual balance cages, along with the small number of replications (n = 9 per group in the balance experiment owing to experimental limitations), might have interacted with the course of infection or its recognition.
Besides this limitation, A. galli-infected growing pullets kept outdoors or on deep litter might contribute to a more effective spreading of parasite eggs if they are fed cereal grains and/or bran with higher concentrations of soluble NSP, inducing an enhanced intestinal viscosity. Besides a general deterioration of individual (excreta sticking to cloacae and to the feathers) and environmental hygiene (wet and sticky litter), the live weight gain and apparent retention of organic matter might be adversely affected under such conditions.
The efficacy of NSP-enzymes added to the diet in counteracting such effects and in influencing the epidemiology of A. galli prevalence needs to be examined in more detail.
References
- Bedford , MR and Schulze , H . 1998 . Exogenous enzymes for pigs and poultry . Nutrition Research Reviews , 11 : 91 – 114 .
- Chadfield , M , Permin , A , Nansen , P and Bisgaard , M . 2001 . Investigation of the parasitic nematode Ascaridia galli (Shrank 1788) as a potential vector for Salmonella enterica dissemination in poultry . Parasitology Research , 87 : 317 – 325 .
- Chubb , LG and Wakelin , D . 1963 . Nutrition and Helminthiasis in Chickens . Proceedings of the Nutrition Society , 22 : 20 – 28 .
- Dänicke , S . 1999 . Zum Einfluss von Nicht-Stärke-Polysacchariden (NSP) und NSP-spaltenden Enzymen auf die Passagezeit der Ingesta sowie den Energie- und Proteinumsatz von wachsenden Schweinen und Broilern . Übersichten zur Tierernährung , 27 : 221 – 273 .
- Dänicke , S , Dusel , G , Jeroch , H and Kluge , H . 1999 . Factors affecting efficiency of NSP-degrading enzymes in rations for pigs and poultry . Agribiological Research , 52 : 1 – 24 .
- Dänicke , S , Böttcher , W , Simon , O , Jeroch , H and Thielebein , J . 2000 . Replacement of soybean oil with tallow in rye-based diets without xylanase increases protein synthesis in small intestine of broilers . Journal of Nutrition , 130 : 827 – 834 .
- Dänicke , S , Valenta , H and Matthes , S . 2007 . On the interactions between Fusarium toxin-contaminated wheat and nonstarch polysaccharide hydrolyzing enzymes in diets of broilers on performance, intestinal viscosity, and carryover of deoxynivalenol . Poultry Science , 86 : 291 – 298 .
- Davies , SF and Joyner , LP . 1955 . Observations on the parasitology of deep litter in poultry houses . Veterinary Record , 67 : 193 – 199 .
- Dusel , G , Kluge , H , Jeroch , H and Simon , O . 1998 . Xylanase supplementation of wheat-based rations for broilers: influence of wheat characteristics . Journal of Applied Poultry Research , 7 : 119 – 131 .
- Gauly , M , Bauer , C , Mertens , C and Erhardt , G . 2001 . Effect and repeatability of Ascaridia galli egg output in cockerels following a single low dose infection . Veterinary Parasitology , 96 : 301 – 307 .
- Gauly , M , Bauer , C , Preisinger , R and Erhardt , G . 2002 . Genetic differences of Ascaridia galli egg output in laying hens following a single dose infection . Veterinary Parasitology , 103 : 99 – 107 .
- Gauly , M , Homann , T and Erhardt , G . 2005 . Age-related differences of Ascaridia galli egg output and worm burden in chickens following a single dose infection . Veterinary Parasitology , 128 : 141 – 148 .
- Hemsley , RV . 1971 . Fourth stage Ascaridia spp. larvae associated with high mortality in turkeys . Canadian Veterinary Journal , 12 : 147 – 149 .
- Herd , RP and McNaught , DJ . 1975 . Arrested development and the histotropic phase of Ascaridia galli in the chicken . International Journal of Parasitology , 5 : 401 – 406 .
- Koski , KG and Scott , ME . 2001 . Gastrointestinal nematodes, nutrition and immunity: breaking the negative spiral . Annual Reviews of Nutrition , 21 : 297 – 321 .
- Langhout , DJ , Schutte , JB , van Leeuwen , P , Wiebenga , J and Tamminga , S . 1999 . Effect of dietary high- and low-methylated citrus pectin on activity of the ileal microflora and morphology of the small intestinal wall of broiler chicks . British Poultry Science , 40 : 340 – 347 .
- Naumann , C and Bassler , R . 1993 . Die Chemische Untersuchung von Futtermitteln , Darmstadt : VDLUFA-Verlag .
- Permin , A and Ranvig , H . 2001 . Genetic resistance to Ascaridia galli infections in chickens . Veterinary Parasitology , 102 : 101 – 111 .
- Permin , A , Bisgaard , M , Frandsen , F , Pearman , M , Kold , J and Nansen , P . 1999 . Prevalence of gastrointestinal helminths in different poultry production systems . British Poultry Science , 40 : 439 – 443 .
- Petkevicius , S . 2007 . The interaction between intestinal helminth infection and host nutrition. Review . Veterinarija ir Zootechnika. T. , 37 : 53 – 60 .
- Roesicke , E and Greuel , E . 1992 . The survival ability of salmonella, coccidia oocysts and ascarid eggs in laying hen feces from different housing systems . Deutsche tierärztliche Wochenschrift , 99 : 492 – 494 .
- Schiemann , R . 1981 . Methodische Richtlinien zur Durchführung von Verdauungsversuchen für die Futterwertschätzung . Archives of Animal Nutrition , 31 : 1 – 19 .
- StatSoft . 1995 . 1984–1995: Statistica for the Windows™ Operating System, Version 7.1 , Tulsa, OK : StatSoft Inc. .
- Tugwell , RL and Ackert , JE . 1952 . On the tissue phase of the life cycle of the fowl nematode Ascaridia galli (Schrank) . Journal of Parasitology , 38 : 277 – 288 .
- Zeller, B. (1990) Vergleichende Untersuchungenen über den Endoparasitenbefall der Haushühner (Gallus gallus var. domesticus l.) beim Wirtschafts- und Rassegeflügel. Diss. Universität München Fb Veterinärmedizin