
ABSTRACT
1. Feather pecking (FP) is said to be a redirection of food-related foraging pecks to feathers. The effects of three foraging enrichments on the pecking behaviours of layer pullets in pens and as hens in cages (Gallus gallus domesticus) were studied.
2. Treatments included no added enrichment (Control), addition of a mix of wheat and sorghum grains (Grain), a mix of wheat and sorghum grains with lucerne and oaten chaff (Chaff), and lucerne hay (Hay). In pens, Hay was provided in racks, while Grain and Chaff were scattered on the floor. In cages, all treatments were provided in racks. It was predicted that enrichments that encouraged more pecking activity would be most successful at reducing FP.
3. Overall pecking activity was similar between all treatments. The Chaff groups tended to FP less than Grain groups, and Hay groups had the least FP. In cages, both Chaff and Hay groups had significantly less FP than either the Grain or Control groups. FP did not reflect stimulus peck rates. Although Chaff groups pecked at their rack significantly less than either Grain or Hay groups, their FP was on a par with the birds in the Hay groups. Feather quality followed these trends.
4. The fact that these results were inconsistent with the original hypothesis led to a proposal for a modified hypothesis. Rather than the appetitive component of food searching motivation being redirected to feathers, it is the appetitive component of exploratory motivation, and the ability to reach the goal to update information about the environment is what feeds back to the motivation which leads to FP. This modified hypothesis is consistent with the results of this and many other studies and with modern concepts of motivation, foraging, exploration and food selection.
Introduction
Feather pecking (FP) in domestic fowls (Gallus gallus domesticus), consisting of damaging (non-aggressive) pecks to feathers of conspecifics, does not occur in small groups under ‘natural’ conditions where birds have access to typical flora and fauna (Chow and Hogan Citation2005). However, it is encountered in all intensive housing systems (Appleby et al. Citation1992; Nicol et al. Citation2013). Consequent feather loss is a commercial problem because of increased feed costs (Herremans et al. Citation1989; Peguri and Coon Citation1993), reduced egg production (El-Lethey et al. Citation2000) and may lead to cannibalism resulting in increased mortality (Cloutier et al. Citation2000; Blokhuis et al. Citation2007). FP is now generally accepted as a pathological behaviour in birds kept in ‘unnatural’ conditions (Chow and Hogan Citation2005).
The usual method of control (beak trimming) consists of removing a portion of the beak. This removes sensitive sensory receptors which may lead to chronic pain (Hughes and Gentle Citation1995). The EU has already banned laying cages (Rodenburg et al. Citation2013) and beak trimming is about to be banned in a number of European countries (Jung and Knierim Citation2019). Yet FP is more difficult to control in large group housing systems and can lead to the more injurious behaviour of cannibalism (Rodenburg et al. Citation2013). Therefore, it would be advantageous to find some way to control FP without the need to beak trim.
Many risk factors are known to influence the incidence of FP, including genotype, hormones, nutritional factors, food form, light, group size, stocking density, flooring, details of housing furnishing and fear and stress (Hughes and Duncan Citation1972; Nicol et al. Citation2013; Rodenburg et al. Citation2013; Schreiter et al. Citation2019). However, most literature stresses the importance of providing foraging material (Van de Weerd and Elson Citation2006; Rodenburg et al. Citation2013; Schreiter et al. Citation2019).
Hughes and Duncan (Citation1972) stated that the risk factors may all have a common path by acting on the tendency to peck. It seems that these are probably connected with an underlying motivation associated with foraging behaviour. Such risk factors could interact in complex ways with an underlying motivation associated with foraging.
Hughes and Duncan (Citation1972), Nicol et al. (Citation2001), Lambton et al. (Citation2010), and Jung and Knierim (Citation2019) showed reduced FP in birds with access to foraging material. Blokhuis and Arkes (Citation1984), Blokhuis (Citation1989), Huber-Eicher and Wechsler (Citation1997) and Huber-Eicher and Wechsler (Citation1998) showed a negative relationship between the amount of foraging activity and rate of FP. It is therefore understood that FP is a redirection of pecks away from foraging to feathers, in the absence of suitable foraging material.
Rudkin (Citation1990) studied the development of FP in four groups of White Leghorns with untrimmed beaks held in cages from hatching to 27 weeks of age. Two groups each had access to a small pen with grassy and sandy areas. Early results showed a negative relationship between activity rates and FP rates in individuals. However, after onset of lay, one group of penned hens developed FP. In this case, there was a positive relationship between total and FP rates, as individuals that pecked at a higher rate showed more FP. Moreover, after all groups were released into an open range with access to tree litter and grassy areas on a 2 ha property total peck rates reduced, while FP declined and ceased after two weeks.
Other studies also had equivocal results. For instance, although Blokhuis (Citation1986) found chicks with litter did less FP than those without after feeding. Before feeding, both showed the same rate of FP. Aerni et al. (Citation2000) found hens with litter only showed less FP if they were fed pellets rather than mash. Chow and Hogan (Citation2005) found that chicks in a stimulus rich pen showed as much FP as those in the control pen, but the enriched chicks had less FP than those that had no exposure when both were in the control pen.
Nørgaard-Nielsen (Citation1989) found that providing straw in baskets successfully reduced FP whether they were pecked at a high rate (when food was in the form of pellets) or at a low rate (when food was in the form of mash). Blokhuis and Van Der Haar (Citation1992) found that adults raised with access to straw litter pecked more than those that were raised with grain added to the litter or the controls, yet their feathering was poorer than grain raised birds. Bessei et al. (Citation2018) found that foraging is not a heritable autonomous behaviour and saw no evidence that FP substituted for foraging, when measured by time spent walking and litter pecking. Dixon et al. (Citation2010) found that peck rates by hens with three different types of enrichment were not reflected by FP rates. They suggested that enrichments are not necessarily effective because they give the animals ‘more to do’. Schreiter et al. (Citation2019) noted that studies have not found consistent results for the benefits of adding various enrichment materials compared to littered housing systems.
Hansen (Citation1994) found a positive correlation between the frequency of feather pecking and the frequency of litter pecking in aviaries and floor pecking in cages. Lindberg and Nicol (Citation1994) found an increased risk of FP by hens with operant feeders compared to those that had food ad libitum. Newberry et al. (Citation2007) found that individual birds that foraged more and were more active when young, FP more both as juveniles and as adults. They concluded that FP did not substitute for foraging behaviour. Bilčἰk and Keeling (Citation2000) showed that individuals that ground pecked more also FP more. Lambton et al. (Citation2010) found increased FP when birds’ feed was spread on litter.
The present study set out to elucidate the effects of different foraging stimulants on pecking behaviours in growing pullets and laying hens. According to the current hypothesis that FP is a redirection of foraging for food, it was expected that stimulants that encouraged more foraging behaviours would be more effective at reducing FP than those that did not. Environmental enrichments were chosen that could be applicable to normal housing systems.
Materials and methods
This experiment was carried out under approval from the Animal Experimental Ethics Committee, University of Queensland. No unusual procedures were carried out, and deaths due to cannibalism were kept to a minimum by daily monitoring during routine care and behavioural observations, and by removing injured birds immediately upon detection.
Birds and housing
One-day-old vaccinated layer pullets with untrimmed beaks (Black SIRO-CB – a layer breed developed by the Animal Genetics section of The Commonwealth Scientific and Industrial Research Organisation (CSIRO) in Australia, and popular as a commercial layer breed at the time of the study) were randomly divided into four groups containing a little over 400 chicks each. Each group was randomly assigned a treatment – Control, Grain, Chaff or Hay. Each group was enclosed within 60 cm high surrounds over a litter floor area of approximately 200 cm diameter and artificially brooded for 20 d. The chicks in each group were then randomly separated into four groups of 100 birds, and each of the 16 groups randomly allocated to a litter floor pen and their initial treatments continued. Each pen measured 370 × 380 cm, so that space allowance was approximately 1400 cm2 per bird, which was close to the commercial space allowance of 1390 cm2 recommended by Nowland (Citation1978). A strip of hardboard 60 cm high was placed between neighbouring pens to restrict each groups’ view of the other. Two tube feeders and two bell waterers were provided in each pen and litter was sawdust spread on the concrete floor. Birds were routinely vaccinated.
On the 24th week of age, 24 birds were randomly selected from each pen, and housed in pairs in cages, with 12 birds per treatment in six cages. In this way, two cage groups were created from each pen group. Each cage group was randomly assigned a position in a shed and given the same treatment that corresponded to the previous enrichment in the pen. The cages were 440 mm deep by 310 mm wide by 400 mm high and were constructed with open wire mesh. A feed trough was placed across the front of the cages. A piece of metal was placed between groups to prevent interaction between the groups. The birds were housed in pairs so that both birds could comfortably reach the rack at the same time. This simulated the conditions in modern wide fronted cages.
The birds were reared from Spring through Summer. Natural daylight was provided during the rearing period. In the laying period, the hens received artificial lighting as well as natural daylight, with lights on at 04.00 h and off at 20.00 h. Light intensity in the pen shed averaged 170 lux (range 60–480 lux) and in the cage shed averaged 160 lux (range 70–400 lux). Birds were fed ad libitum with starter crumbles to the fifth week, followed by finely ground grower mash to the 22nd week of age, and finely ground layer mash thereafter.
Treatments – floor pens
In floor pens, the following treatments were provided from the 11th day; 1. Control: no added stimuli; 2. Grain: wheat and sorghum (lightly rolled in the first eight weeks) mixed by weight in the proportion of three parts of wheat to one part of sorghum; 3. Chaff: grain as above, plus lucerne (Medicago sativa) and oaten chaff (Avena sativa) mixed by weight in the proportion of two parts of lucerne to one part of oaten chaff, plus a measured amount of insoluble grit (white quartz granules approximately 2–3 mm diameter). The chaff mix contained approximately 50% dry leaf and 50% stalks. The median length of stalks was 9 mm with a range of 1 mm to 100 mm with the majority (70%) between 5 mm and 11 mm. 4. Hay: field dried lucerne which contained a small admixture of unidentified grass and weeds depending on season, but the bulk of the material was lucerne. This consisted of dried leaves and stems whose lengths varied from approximately 20 mm to 450 mm.
Total food consumption was measured and compared with the consumption of the enrichment stimuli. By weight, grain formed approximately 8% of the total diet of the Grain groups. Grain was 8%, lucerne 1% and oaten chaff 0.5% of the total diet in Chaff groups, and lucerne 3% assuming 50% of the hay was swallowed (many stalks were not swallowed) of the Hay groups.
Grain and chaff mixes were scattered on the floor six mornings a week and hay was provided in racks and was replenished, as required, each morning. Quantities of grain, chaff and grit were gradually increased until by the sixth week each Grain group was receiving a one litre measure of grain, and each Chaff group was receiving one litre of grain, two litres of chaff and 100 ml of grit, which were premixed before being scattered on the floor.
Hay was at first provided in various perforated containers until the fifth week, when hay racks were constructed using 1.2 cm square wire netting, rolled into a cylinder 45 cm in diameter by 90 cm in length with the ends being closed by netting. These racks were initially approximately 10 cm from the floor, then raised as the birds grew.
All birds were weighed when the groups were set up at five weeks of age and when feather scored at 16 and 24 weeks. Ration and treatment foods were weighed, and the amount consumed by each group was estimated over the two time periods between weighing and food efficiency calculated. The effects of treatments on nutrient intakes were estimated using tables of analysed nutrient levels of local feedstuffs (Evans Citation1985).
Treatments – cages
In cages, the following treatments were presented in racks suspended above the food trough; 1. Control: no stimulus and housed in the normal way without a rack. 2. Grain: a complete layer ration mix containing the following proportions by weight; 10% lightly rolled barley; 57.9% lightly rolled wheat; and 8% limestone in the form of coarse chips (mean diameter 3 mm, range of 1–5 mm). Grains were rolled because insoluble grit was no longer available. Over weeks 27–31 and 61–64 powdered limestone was substituted because chips were unavailable. In the first five weeks after caging, limestone, yolk colourant, oil and vitamin mix were omitted. However, when it became apparent that the contents of the racks formed approximately 50% of the intake of the birds, the complete mix was used as the stimulus ration. 3. Chaff: one part of the coarse food mix (without the layer mix), mixed by volume with three parts of a chaff mix (pre-mixed in the proportions of three parts of lucerne chaff to one part by weight of oaten chaff). 4. Hay: the stimulus in racks was lucerne hay similar to that used in the pens.
The grain and chaff racks had solid sides and backs with 0.6 mm square wire mesh fronts. The mesh on the front of the grain racks was doubled to prevent the grain mix from falling through. The hay racks had 12 mm square mesh on fronts, backs and sides. Pecked material that spilled from the racks fell into the food troughs. Both birds in each cage could comfortably peck at the rack at the same time.
During the sixth week in the pens, all birds in one of the control groups and one of the grain groups were beak-trimmed after an outbreak of cannibalism. The trimmed birds were excluded from all analyses. This resulted in, for pens, three replicates of 100 untrimmed birds in the Control and Grain treatments, and four in the Chaff and Hay treatments, and, in cages, six replicates of 12 untrimmed birds in the Control and Grain treatment groups, and eight replicates of 12 birds in the Chaff and Hay treatments.
Feed ration and stimulus use were measured at four to 10 weekly intervals (weeks 24, 28, 35, 40, 45, 51, 58, 61, 64, 68, 78). Eggs produced by each group were counted daily and were weighed each day for the first 20 weeks, then three days a week subsequently. Feed efficiency was calculated for each time period between food weighings. All chaff and hay removed from racks were included. The effects of treatments on nutrient intakes were estimated using tables of analysed nutrient levels of local feedstuffs (Evans Citation1985).
Observations
Ten birds were randomly selected from each pen group and six from each cage group, and their behaviour observed. In pens, selected birds were identified with a coloured numbered band on each leg and marked with acrylic paint on head and back for ease of recognition. In the cages, all birds were banded on each leg but were not marked with paint.
Groups, and birds within them, were observed in a predetermined random order, at a random time of day, with each selected bird being observed for five minutes. The birds were observed from within the pens, or directly in front of the cages, and five minutes were allowed to elapse before any observations were made. Care was taken to wear similar clothes each day, and to avoid sudden movements or sounds, to ensure the birds ignored the observer. To measure typical activity patterns of the birds over the greater part of the day, birds were not observed within one hour of the morning attendance. The selected animals in each pen group were observed twice at three ages (at 8–9, 13–15, and 17–19 weeks). In each cage group they were observed once each at six ages (at 27–29, 31 − 33, 40–44, 47–49, 66–70, and 75–77 weeks).
Behaviours measured included the following: 1. Food pecks: number of pecks directed at food in feeders in pens or troughs in cages. 2. Rack pecks in cages: number of pecks directed at the racks. 3. Ground pecks: in pens number of pecks directed at the ground. 4. Ground scratching: in pens, each series of backward leg scrapes usually followed by ground pecking was recorded as one event. 5. Feather pecks: number of non-aggressive damaging pecks at feathers either directed at the focal animal or by the focal animal (categories described as ‘pull’ and ‘barb pulling’ by Wood-Gush and Rowland Citation1973). Not included were pecks at foreign matter on feathers (such as sawdust) and very light pecks that did not disturb the feathers (classified as ‘peck’ by Wood-Gush and Rowland Citation1973). 6. Other pecks: in pens, those that did not fit into the above categories, apart from aggressive pecks; and included pecks at the racks, pen structures and furnishings, air pecks, floating particles, and at the observer’s clothing. In cages, this included pecks at cage structures, air pecks and pecks at floating particles. 7. Total pecks: all those delivered by the focal animal apart from aggressive pecks. It included counts of all other and feather pecks. Aggressive pecks were recorded but were not included in the analysis, and were usually readily distinguished from feather pecks. These pecks were generally sudden single pecks and directed towards the back of the head and neck with a downward strike. FP were generally carefully aimed and directed at other parts of the body with a horizontal strike.
Feather scoring
Every bird in each group was feather scored at three ages in pens, and at six ages in cages (7, 16 and 24 and 30, 36, 44, 53, 61 and 71 weeks, respectively). Each of six body parts (back, tail, neck and crop, wings, sides, and abdomen) was separately scored out of five, and added to give a total score out of 30 for each bird using the following criteria:
5 – perfect feathering with little or no sign of feather damage.
4 – evidence of pecking, such as broken feathers and stubs of feather shafts but no areas devoid of feathers.
3 – some areas devoid of feathers but occupying less than ¼ of the body part.
2 – areas devoid of feathers occupying at least ¼ but less than ½ of the body part.
1 – in pens, areas devoid of feathers extending over at least ½ of the body part, and in cages, bare areas occupying at least ½ but less than 4∕5 of the body part.
0 – in pens, presence of wounds, and in cages, at least 4∕5 of the body part being devoid of feathers. The method of scoring was changed slightly in cages, as wounded birds were more readily discovered and removed.
When an assistant re-scored four cage groups at 71 weeks, the Spearman’s correlation coefficient was 0.9963 (P < 0.00003).
Statistical analyses
All statistical analyses were carried out using the General Linear Models procedure in the SAS System for Windows Version 6.10 (Citation1991, 94). All data on individual birds were averaged so that all variables were reduced to a per bird per group basis. The appropriateness of combining the data over time was evaluated using a sphericity test, and the significance of interactions was tested using the Wilks’ Lambda test statistic from repeated measures analysis. Where the data passed the sphericity test, and interaction with age was not significant (i.e. total pecks and food pecks in pens and cages; ground pecks and ground scratching in pens; and rack pecks and feather pecks in cages), a split-plot over time analysis of variance model estimating the combined effect of age, treatment, and their interaction was used. Where the sphericity test failed, but interaction with age was not significant (i.e. feather pecks and other pecks in pens), treatment effects were estimated from the split-plot model. Where interaction with age was significant (i.e. feather scores in pens and cages), treatment effects were estimated at each time level with one-way analysis of variance. Pair-wise comparisons were made using Least Squares from the relevant models.
Feather scores were expressed as a proportion of total possible score, and were arcsine transformed before analysis. The common behaviours of ground pecking, food pecking in cages and total pecks were transformed using loge (x + 0.5) before analysis. The rarer or more sporadic behaviours of rack and feather pecks, and of ground scratching and food pecks in pens, were expressed as the probability of a focal bird engaging in the behaviour in any one observation period, and were arcsine transformed before analysis. Residual diagnostics were used to check the validity of the models used.
To test whether cage groups created from the same pen of origin had a greater degree of similarity than those from different pens, variation between groups was partitioned into a pen of origin component and a residual component. Where the residual component was significantly less than the between pen component, then the between-pen variance was used to assess the significance of treatment effects. Pen of origin had a significant effect on feather score data, but not on any peck data.
Rate of decline of feather quality in cages was estimated for each pen of origin by regressing feather scores against the inspection time. The slopes of the regressions for each pen of origin were then analysed for treatment effects using analysis of variance.
Results
Behaviour rates – growing pullets in pens
There was no evidence that total peck rates, ground or food pecks or ground scratching differed between treatments. FP did not significantly interact with age (F6,18 = 1.7610, P < 0.164). There was a significant treatment effect for the incidence of FP in the Hay groups where the FP was significantly less than all other treatment groups. Chaff groups tended to FP less than Grain groups (P < 0.058). Other pecks for Control, Grain and Chaff groups did not differ, but Hay groups ‘other’ pecked significantly more than the Control and Chaff groups. Although the score for other pecks in the Hay groups included pecks at the racks, they did not differ significantly from the results for the Grain groups that only included pecks at various parts of the furnishings (). Other pecks formed a low proportion of total pecks in all groups, being 1.6, 2.6, 0.7 and 4.8% of total pecks in the Control, Grain, Chaff and Hay groups, respectively.
Table 1. Behaviour rates of pullets in each treatment in pens. Values for feather pecks, food pecks, other pecks and ground scratching are mean per cent probability of a focal animal engaging in the behaviour, and values for ground and total pecks are mean peck rate per five minutes. Means have been backtransformed
Hens in cages
There was no evidence that total or food peck rates differed between treatments. There was a significant treatment effect for FP (F3,24 = 7.12, P < 0.001; ). Control and Grain groups FP significantly more than the Hay and Chaff groups. FP rates in all treatments were comparatively low, means being 0.1% to 0.9% of total pecks. There was a significant treatment effect for rack pecks (F2,24 = 6.17, P < 0.009) for birds in the Grain and Hay groups, where they pecked racks significantly more than birds in the Chaff groups ().
Table 2. Behaviour rates of hens in each treatment in cages. Values for total and food pecks are mean peck rate per five minutes, and values for feather pecks and rack pecks are mean percent probability of a focal animal engaging in the behaviour. Means have been back transformed
Rack pecking was a comparatively rare behaviour (means ranged from 2% to 3% of total peck rate). There were no overall trends with time. The behaviour of the hens at the racks was variable. Pecks at grain racks caused food to spill into the trough. Sometimes pecks were directed at the rack, and, at other times, only a few pecked at the rack which spilled stimulus food. Sometimes one bird pecked the rack while the neighbours pecked at the spilled stimulus food. Hay was pecked from both the fronts and tops of the racks, causing some particles to fall into the trough where leaves and small stalks were eaten, while larger stalks were mandibulated and either dropped or, if small enough, swallowed. Fifty per cent of the food eaten by the Grain groups was stimulus food, while 9% if the stimulus food was eaten in the Chaff groups, and 13% of the stimulus hay eaten by Hay groups (if all material removed from the racks was eaten).
Feathering – pullets in pens
There was a significant interaction between feather quality and age (Wilks’ Lambda F6,18 = 4.4945, P < 0.006). At 16 weeks the feathering of growing pullets in the Hay treatment was significantly better than all other treatments (P < 0.019, 0.001, 0.017 when compared with Control, Grain, and Chaff respectively). Grain treatments were significantly worse than all other treatments at 7 and 16 weeks of age (). Feathering tended to worsen with age in all treatment groups. Control treatment birds’ feathering worsened significantly at each age, and, in the Hay treatment group, feathering worsened significantly from 16 weeks to 24 weeks of age. On the other hand, in Grain and Chaff treatment groups, feathering worsened significantly between 7 and 16 weeks of age, but not between 16 and 24 weeks ().
Figure 1. Feather scores over the growing period (arcsine of mean±SEM). Higher scores indicate better plumage condition.
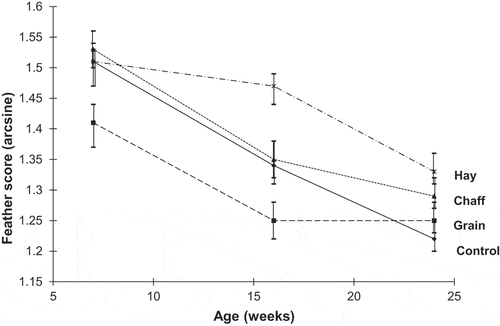
Except for the occasional individual, backs and/or tails were the only body parts affected in pecked individuals. Back of the birds with Hay treatments were significantly better than all other treatments at 16 weeks of age. Their tails were significantly better than birds in the Grain treatments at 16 weeks and better than Control and Grain treatments at 24 weeks of age ( and ).
Table 3. Tail feather scores (arcsine of mean) of growing pullets in pens at three ages
Table 4. Back feather scores (arcsine of mean) of growing pullets in pens at three ages
Feathering – hens in cages
Feather scores interacted significantly with age (Wilks’ Lambda F2,16.96 = 2.4177, P < 0.042). Feather quality varied significantly between treatments from 44 weeks of age, with the best feather quality seen in the Hay groups and worst in the Control groups (; ).
Table 5. Mean feather scores (out of 30) of layer hens in cages. Higher scores indicate better plumage condition
Figure 2. Feather scores over the laying period (arcsine of mean±SEM). Higher scores indicate better plumage condition.
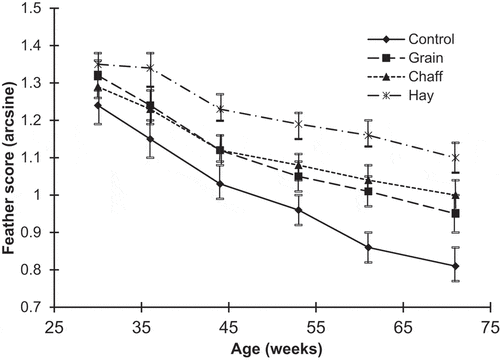
Pen of origin had a significant effect on feather scores in the Control and Grain groups, so between pen variance was used as the error term for all feather score analyses. Chaff and Hay group birds’ feathering was not affected by pen of origin. Analyses using between-group variance as the error term found that Hay group birds' feathering was significantly better than Chaff group at 44 and 53 weeks of age, as well as at 61 weeks (P < 0.016, 0.015, and 0.015, respectively). This is compared with P < 0.073, 0.082, and 0.049, respectively, when between pen variances were used as the error term.
The average weekly decrease in feather scores was 0.270, 0.201, 0.157, and 0.115 for Control, Grain, Chaff and Hay treatment groups, respectively (). Analysis of variance on slopes showed a significant treatment effect (F3,10 = 10.32, P < 0.002). Feather quality of Hay group birds did not decline as rapidly as seen in the Control and Grain groups over the laying period (P < 0.003 and 0.014, respectively), and, for birds in the Chaff and Grain groups, feather quality did not decline as rapidly as in the Control groups (P < 0.003 and 0.049 respectively).
Feathers on all parts of the body were pecked, but the most severely affected areas were the neck and crop (). The damage to neck and crop areas was probably due to between cage pecking, as birds were often seen reaching across the fronts of cages to peck their neighbours’ feathers (the piece of metal between groups prevented this type of pecking between groups).
Table 6. Mean feather scores (out of 5) for separate parts at end of laying period (71 weeks), backtransformed from arcsined form. Higher scores indicate better plumage condition
Nutrient levels
The effects of treatments on nutrient levels were minor in pen housed birds. In laying hens, tryptophan and threonine levels were unaltered, while lucerne chaff and hay increased dietary fibre slightly. Concentration of fibre was 3.5% in ration, 26.0% in lucerne and 32.5% in oaten chaff (Evans Citation1985). In cages, lucerne formed 13% and 9% of food eaten in the Hay and Chaff groups, respectively, assuming all contents removed from the racks was eaten. This raised total fibre in the diet, from 3.5% to 6.4% and 5.1% for Hay and Chaff groups, respectively, if all the hay or chaff was eaten; or to 5.1% and 4.4% for the hay and chaff groups if half the hay or chaff was eaten.
Levels of most nutrients were a little lower in birds receiving lucerne than in the layer ration, which may have slightly lowered their total intake. These included calcium, available phosphorus, sodium, linoleic acid, total protein; and the amino acids methionine, cystine, lysine, arginine, isoleucine, leucine, histidine, valine, phenylalanine and tyrosine.
Feed conversion
Treatments did not affect feed conversion significantly during the penned period, but in cages, feed to egg conversion was significantly reduced in Grain, Chaff and Hay treatments compared to the Control group (). However, egg production and egg weights were not significantly altered, even though all chaff or hay removed from racks were included in the calculation.
Table 7. Mean feed efficiency. For pullets in pens: period 1 weeks 5 to 16, period 2 weeks 16 to 24. As hens in cages over the 12 month laying cycle. Feed efficiency was calculated as if all the hay or chaff loss was consumed
Discussion
This study found that some enrichments were more successful than others at reducing FP without altering total peck rates in both rearing pens and layer cages, but this did not correlate with the amount of activity directed at the stimuli provided. Feather quality followed this trend. In pens, ground directed behaviour over the greater part of the day was not increased by adding grain or grain plus chaff to the litter. Yet, the birds in the Chaff group tended to FP less than in the Grain groups. Although the racks were pecked by birds in the Hay groups, it did not increase overall peck rates, yet their FP was significantly less than those in other treatment groups and feathering followed this trend, with significantly better feathering at 16 weeks of age. Moreover, although other pecks in pens included those directed at the racks in the Hay groups, these were not significantly greater than seen in the Grain groups, which only included pecks at furnishings. Yet, as above, the birds in the Grain groups showed FP more than Chaff or Hay groups.
In cages, rack pecking did not reflect FP. Birds in the Chaff groups rack pecked significantly less than in the Grain or Hay groups. Yet the FP was significantly less than in the Grain group and at about the same rate as the Hay groups. In spite of the fact that birds in the Grain groups rack pecked as much as Hay groups, the FP was similar to the Control groups. This was in contradiction to many studies that showed that stimuli which increased activity decreased FP (e.g. Blokhuis and Arkes Citation1984; Blokhuis Citation1989; Huber-Eicher and Wechsler Citation1997, Citation1998) which led to the hypothesis that the motivation leading to FP was a redirection of pecking associated with foraging for food. It seems that there was something about the stimuli in the present study that reduced FP without increasing activity rates.
It was possible that the lucerne used as the stimulus was successful because it provided extra fibre. Increased coarse fibre in the diet is known to reduce FP as both fibre and feather intake increases the passage of food through the gut (Van Krimpen et al. Citation2005; Harlander-Matauschek et al. Citation2006) and may be reinforced by palatability (Harlander-Matauschek et al. Citation2008). Van Krimpen et al. (Citation2005) recommended increasing fibre in the diet from 29 to 123 g/kg to reduce feather pecking. However, in the present study, in pens, the lucerne formed just 3% (30 g/kg) of the diet and in cages, lucerne increased dietary fibre levels from 3.5% to 4.4% (35 to 44 g/kg) in the Chaff diet, and to 5.1% (51 g/kg) in the Hay diet, assuming half the hay or chaff loss from the racks was consumed. This was an estimation only. It was likely that many stalks in the chaff, and considerably less than half the hay was actually swallowed. This was considerably less than that recommended by Van Krimpen et al. (Citation2005). Furthermore, supplementary lucerne did not significantly improve feathering in the study by Kalmendal and Wall (Citation2012). It was notable that the chaff on the floor in the pens in the present study was less effective at reducing FP than chaff in racks in the cages.
Treatments did not affect feed conversion in pens, but in cages, feed conversion was significantly poorer in Grain, Chaff and Hay than in Control groups. Weight gain did not differ significantly for the pullets in pens, and egg production and egg mass did not differ significantly for laying hens in cages. Therefore, it seemed unlikely that lucerne increased feed intake in pens or cages. This indicated that increased fibre provided by the lucerne in the present study was probably not an important contributing factor which influenced the results. However, it could not be discounted that some fibre accumulated in the digestive tract, making feathers less attractive.
Nutrient deficiency is known to exacerbate FP (Hughes and Duncan Citation1972; Van Krimpen et al. Citation2005), but the birds were provided a complete feed mix. In the early weeks after caging, the grain group birds obtained half their food from the racks. This may have exacerbated FP before the complete mix was added to their racks. Examination of nutrients provided by lucerne showed some reduction, rather than an increase, in nutrient levels, so it was unlikely to be a factor in this study. However, it was possible lucerne could have been providing some missing unknown nutrient.
The present study was designed to test the motivational causes of FP, which have been associated with foraging behaviour. Any motivational state is said to be controlled by feedback systems. Negative feedback can occur from the performance of a behaviour pattern (appetitive) or from the result of the behaviour (consummatory). If an animal is prevented by lack of stimuli to limit the behaviour, or from providing the goal, it may result in the development of stereotypies or injurious behaviour (Wiepkema Citation1983; Hughes and Duncan Citation1988; Dantzer Citation1991; Jensen and Toates Citation1993).
Hughes and Duncan (Citation1972), Nicol et al. (Citation2001), Lambton et al. (Citation2010), and Jung and Knierim (Citation2019) showed reduced FP in birds with access to foraging material. Blokhuis and Arkes (Citation1984), Blokhuis (Citation1989), Huber-Eicher and Wechsler (Citation1997), and Huber-Eicher and Wechsler (Citation1998) showed a negative relationship between amount of foraging activity and the rate of FP. These studies were consistent with the idea that enhancement of the environment stimulates foraging behaviour, hence reducing FP by redirection of a motivational state associated with feeding or foraging for food.
Other studies, as in the present trial, were inconsistent with this view (Nørgaard-Nielsen Citation1989; Rudkin Citation1990; Blokhuis and Van Der Haar Citation1992; Hansen Citation1994; Lindberg and Nicol Citation1994; Bilčἰk and Keeling Citation2000; Newberry et al. Citation2007; Bessei et al. Citation2018; Dixon et al. Citation2010). This led Newberry et al. (Citation2007) to reject the redirection hypothesis. However, where there is no relationship, it may be the case that the birds are unable to reach the goal of the behaviour, and the appetitive component continues unabated.
Foraging is thought to be an expression of two motivations. One is extrinsic; to search for food, and the other is intrinsic; to update information about the environment (Birke and Archer Citation1983) and the two may be difficult to distinguish (Wood-Gush et al. Citation1983).
The goal of foraging is to find food, but, if this were the case, more food-like stimuli should be more successful at reducing FP. However, the addition of grain on the floor of the rearing pens or in racks in laying cages did not reduce FP. Other studies did not reduce FP with grain; indeed, in some, grain stimulus increased FP when added to litter (Lambton et al. Citation2010) or when presented in operant feeders (Lindberg and Nicol Citation1994). Lambton et al. (Citation2010) suggested that although the grain stimulated foraging behaviour, it was not sufficiently rewarding.
There is evidence that chickens are able to balance their nutrient requirements (Overmann Citation1976; Rozin Citation1976; Emmans Citation1977; Hughes Citation1984), and are perhaps motivated to search for a variety of food sources. This would explain the tendency seen for grain plus chaff in the rearing pens, and the success of grain and chaff to reduce FP in racks of laying cages. However, this did not explain the success of lucerne hay in racks at reducing FP. Other studies have been able to reduce FP by presenting a less food-like stimulus, such as straw, when presented in bundles or baskets, but not as loose straw (Nørgaard-Nielsen Citation1989; Huber-Eicher and Wechsler Citation1998). Some other characteristic of the lucerne or straw may be feeding back into motivation leading to FP. This may have been because the birds had to remove the lucerne or straw from the containers and mandibulate them to assess edibility.
Inglis (Citation1983) and Inglis et al. (Citation2001) argued that information gathering is a prime need which is continuously present and underlies goal directed behaviours. Newberry (Citation1999) showed that hens are motivated to explore novel stimuli and stated that birds have an intrinsic motivation to perform exploratory behaviour (i.e. exploring for its own sake rather than searching for currently needed resources) and that the motivation is not satisfied when chickens are kept in a relatively constant environment. Inglis (Citation1983) wrote that animals experience satiation when given an opportunity to explore, and stimulus change can be rewarding in itself. Therefore, when the birds extracted lucerne from the racks, they may have been able to investigate them for edibility and experienced stimulus change.
This motivation may be adaptive in the wild, as it would lead the birds to find new food resources when feedstuffs may be only available for a few weeks in the life of any one individual. Learning a beneficial food requires several tastings and is more slowly acquired than reacting to its toxic effects (Rozin Citation1976). This may be retained in the domestic situation, as there is no selective pressure against it (Wood-Gush et al. Citation1983).
Conclusions
This study examined the motivation leading to FP. It found that neither overall activity rates, nor the activity directed at stimuli predicted FP. Based on these findings, it can be suggested that the motivation leading to FP, rather than being a redirection of the appetitive behaviour of foraging for food, is a redirection of the appetitive component of intrinsic exploration. The ability to reach the goal of updating information about the environment is what appeared to help control motivation leading to FP. This explanation is consistent with the results of this study and many other studies that were found to be inconsistent with the usual explanation.
Based on the findings of this study, it can be recommended that lucerne hay is provided in perforated containers to reduce the incidence of FP. It is not advisable to add grain to litter or in containers. This could be combined with other factors known to reduce FP, such as genetic selection, reducing maternal stress, improving the stockmanship skills of staff, reducing housing density, provision of perches, non-pelleted feeds, extra fibre, litter floors, early access to litter and employing methods to improve range use.
Acknowledgments
Many thanks to the anonymous reviewers and their helpful comments. The revised manuscript is now clearer and adheres better to the results of this study. This work was undertaken as part of the work for the degree of Doctor of Philosophy. Thanks go to my PhD supervisor, Dr John Dingle. I am indebted to Allan Lisle for assistance with the statistical analysis. I wish to thank members of the Department of Animal Production (now the School of Agriculture and Food Sciences) and Gatton Farms who helped in various ways, and, in particular, to thank Felix Gorbacz, Mark Davies, Sue Berry and Dominic Berry for assistance at critical times. Thanks to Associate Professor Dominic W. Berry for his help in preparation of this manuscript. I wish to acknowledge my mentor Emeritus Professor Glen McBride for his patient guidance during preparation of my thesis for the Master of Science.
The Alan White Scholarship, awarded by the Royal Society for the Prevention of Cruelty to Animals (RSPCA), helped with expenses, and the work was performed in facilities provided by the Department of Animal Production (now the School of Agriculture and Food Sciences) and Gatton Farms at the University of Queensland Gatton.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Aerni, V., H. El-Lethey, and B. Wechsler. 2000. “Effect of Foraging Material and Food Form on Feather Pecking in Laying Hens.” British Poultry Science 41: 16–21. doi:https://doi.org/10.1080/00071660086349.
- Appleby, M. C., B. O. Hughes, and H. A. Elson. 1992. Poultry Production Systems: Behaviour, Management and Welfare. Wallingford, Oxon, UK: CAB International.
- Bessei, W., J. B. Lutz, L. B. Kjaer, M. Kjaer, M. Grashorn, and J. Bennewitz. 2018. “Relationships between Foraging and Open-Field Activity in Young Chicks and Feather Pecking in Adult Birds: Results of Analyses Using Quantitative Genetics and Structural Equation Models.” European Poultry Science 82. 1612–9199. doi:https://doi.org/10.1399/eps.2018.242.
- Bilčἰk, B., and L. J. Keeling. 2000. “Relationship between Feather Pecking and Ground Pecking in Laying Hens and the Effect of Group Size.” Applied Animal Behaviour Science 68: 55–66. doi:https://doi.org/10.1016/S0168-1591(00)00089-7.
- Birke, L. I. A., and J. Archer. 1983. “Some Issues and Problems in the Study of Animal Exploration.” In Exploration in Animals and Humans, edited by J. Archer and L. I. A. Birke, 1–21. Wokingham: Van Nostrand Reinhold, UK, Co.
- Blokhuis, H. J. 1986. “Feather Pecking in Poultry: Its Relation with Ground-Pecking.” Applied Animal and Behavioural Science 16: 63–67. doi:https://doi.org/10.1016/0168-1591(86)90040-7.
- Blokhuis, H. J. 1989. “The Effect of a Sudden Change in Floor Type on Pecking Behaviour in Chicks.” Applied Animal Behaviour Science 22: 65–73. doi:https://doi.org/10.1016/0168-1591(89)90080-4.
- Blokhuis, H. J., and J. G. Arkes. 1984. “Some Observations on the Development of Feather-Pecking in Poultry.” Applied Animal Behaviour Science 12: 145–157. doi:https://doi.org/10.1016/0168-1591(84)90104-7.
- Blokhuis, H. J., and J. W. Van Der Haar. 1992. “Effects of Pecking Incentives during Rearing on Feather Pecking of Laying Hens.” British Poultry Science 33 (1): 17–24. doi:https://doi.org/10.1080/00071669208417440.
- Blokhuis, H. J., T. Fiks-Van Niekerk, W. Bessei, A. Elson, D. Guemene, J. Kjaer, G. A. Maria Levrino, et al. 2007. “The LayWel Project: Welfare Implications of Changes in Production Systems for Laying Hens.” World’s Poultry Science Journal 63 :101–114. doi:https://doi.org/10.1017/S0043933907001328.
- Chow, A., and A. Hogan. 2005. “The Development of Feather Pecking in Burmese Red Junglefowl: The Influence of Early Experience with Exploratory-Rich Environments.” Applied Animal Behaviour Science 93: 283–294. doi:https://doi.org/10.1016/j.applanim.2005.01.004.
- Cloutier, S., R. C. Newberry, C. T. Forster, and K. M. Girsberger. 2000. “Does Pecking at Inanimate Stimuli Predict Cannibalistic Behaviour in Domestic Fowl?” Applied Animal Behaviour Science 66: 119–133. doi:https://doi.org/10.1016/S0168-1591(99)00068-4.
- Dantzer, R. 1991. “Stress, Stereotypies and Welfare.” Behavioural Processes 25: 95–102. doi:https://doi.org/10.1016/0376-6357(91)90012-O.
- Dixon, L. M., I. J. H. Duncan, and G. J. Mason. 2010. “The Effects of Four Types of Enrichment on Feather-Pecking Behaviour in Laying Hens Housed in Barren Environments.” Animal Welfare 19: 429–435. http://hdl.handle.net/10214/4956
- El-Lethey, H., V. Aerni, T. W. Jungi, and B. Wechsler. 2000. “Stress and Feather Pecking in Laying Hens in Relation to Housing Conditions.” British Poultry Science 41: 22–28. doi:https://doi.org/10.1080/00071660086358.
- Emmans, G. C. 1977. “The Nutrient Intake of Laying Hens Given a Choice of Diets, in Relation to Their Production Requirements.” British Poultry Science 18: 227–236. doi:https://doi.org/10.1080/00071667708416358.
- Evans, M. 1985. Nutrient Composition of Feedstuffs for Pigs and Poultry. Brisbane: Queensland Department of Primary Industries.
- Hansen, I. 1994. “Behavioural Expression of Laying Hens in Aviaries and Cages: Frequencies, Time Budgets and Facility Utilisation.” British Poultry Science 35 (4): 491–508. doi:https://doi.org/10.1080/00071669408417715.
- Harlander-Matauschek, A., F. Wassermann, J. Zentek, and W. Bessei. 2008. “Laying Hens Learn to Avoid Feathers.” Poultry Science 87: 1720–1724. doi:https://doi.org/10.3382/ps.2007-00510.
- Harlander-Matauschek, A., H. P. Piepho, and W. Bessei. 2006. “The Effect of Feather Eating on Feed Passage in Laying Hens.” Poultry Science 85: 21–25. doi:https://doi.org/10.1093/ps/85.1.21.
- Herremans, M., E. Decuyperere, and O. Siau. 1989. “Effects of Feather Wear and Temperature on Prediction of Food Intake and Residual Food Consumption.” British Poultry Science 30 (1): 15–22. doi:https://doi.org/10.1080/00071668908417121.
- Huber-Eicher, B., and B. Wechsler. 1997. “Feather Pecking in Domestic Chicks: Its Relation to Dustbathing and Foraging.” Animal Behaviour 54: 757–768. doi:https://doi.org/10.1006/anbe.1996.0506.
- Huber-Eicher, B., and B. Wechsler. 1998. “The Effect of Quality and Availability of Foraging Materials on Feather Pecking in Laying Hen Chicks.” Animal Behaviour 55: 861–873. doi:https://doi.org/10.1006/anbe.1997.0715.
- Hughes, B. O. 1984. “The Principles Underlying Choice Feeding Behaviour in Fowls - with Special Reference to Production Experiments.” World’s Poultry Science Journal 40 (2): 141–150. doi:https://doi.org/10.1079/WPS19840012.
- Hughes, B. O., and I. J. H. Duncan. 1972. “The Influence of Strain and Environmental Factors upon Feather Pecking and Cannibalism in Fowls.” British Poultry Science 13: 525–547. doi:https://doi.org/10.1080/00071667208415981.
- Hughes, B. O., and I. J. H. Duncan. 1988. “The Notion of Ethological ‘Need’, Models of Motivation and Animal Welfare.” Animal Behaviour 36: 1696–1707. doi:https://doi.org/10.1016/S0003-3472(88)80110-6.
- Hughes, B. O., and M. J. Gentle. 1995. “Beak Trimming of Poultry, Its Implications for Welfare.” World’s Poultry Science Journal 51: 51–61. doi:https://doi.org/10.1079/WPS19950005.
- Inglis, I. R. 1983. “Towards a Cognitive Theory of Exploratory Behaviour.” In Exploration in Animals and Humans, edited by J. Archer and I. A. Birke Lynda, 72–116. Wokingham: Van Nostrand Reinhold, UK, Co.
- Inglis, I. R., S. Langton, B. Forkman, and J. Lazarus. 2001. “An Information Primacy Model of Exploratory and Foraging Behaviour.” Animal Behaviour 62: 543–557. doi:https://doi.org/10.1006/anbe.2001.1780.
- Jensen, P., and F. M. Toates. 1993. “Who Needs ‘Behavioural Needs’? Motivational Aspects of the Needs of Animals.” Applied Animal Behaviour Science 37: 161–181. doi:https://doi.org/10.1016/0168-1591(93)90108-2.
- Jung, L., and U. Knierim. 2019. “Differences between Feather Pecking and Non-Feather Pecking Laying Hen Flocks regarding Their Compliance with Recommendations for the Prevention of Feather Pecking – A Matched Concurrent Case-Control Design.” Applied Animal Behaviour Science 219: article 104839. doi:https://doi.org/10.1016/j.applanim.2019.104839.
- Kalmendal, R., and H. Wall. 2012. “Effects of a High Oil and Fibre Diet and Supplementary Roughage on Performance, Injurious Pecking and Foraging Activities in Two Layer Hybrids.” British Poultry Science 53 (2): 153–161. doi:https://doi.org/10.1080/00071668.2012.682146.
- Lambton, S. L., T. G. Knowles, C. Yorke, and C. J. Nicol. 2010. “The Risk Factors Affecting the Development of Gentle and Severe Feather Pecking in Loose Housed Laying Hens.” Applied Animal Behaviour Science 123: 32–42. doi:https://doi.org/10.1016/j.applanim.2009.12.010.
- Lindberg, A. C., and C. J. Nicol. 1994. “An Evaluation of the Effect of Operant Feeders on Welfare of Hens Maintained on Litter.” Applied Animal Behaviour Science 41: 211–227. doi:https://doi.org/10.1016/0168-1591(94)90024-8.
- Newberry, C., L. J. Keeling, I. Estevez, and B. Bilcˇi´k. 2007. “Behaviour When Young as a Predictor of Severe Feather Pecking in Adult Laying Hens: The Redirected Foraging Hypothesis Revisited.” Applied Animal Behaviour Science 107: 262–274. doi:https://doi.org/10.1016/j.applanim.2006.10.010.
- Newberry, R. C. 1999. “Exploratory Behaviour of Young Domestic Fowl.” Applied Animal Behaviour Science 63: 311–321. doi:https://doi.org/10.1016/S0168-1591(99)00016-7.
- Nicol, C. J., A. C. Lindberg, A. J. Phillips, S. J. Pope, L. J. Wilkins, and L. E. Greene. 2001. “Influence of Prior Exposure to Wood Shavings on Feather Pecking, Dustbathing, and Foraging in Adult Laying Hens.” Applied Animal Behaviour Science 73: 141–155. doi:https://doi.org/10.1016/S0168-1591(01)00126-5.
- Nicol, C. J., M. Bestman, A.-M. Gilani, E. N. De Haas, I. C. De Jong, S. Lambton, J. P. Wagenaar, et al. 2013. “The Prevention and Control of Feather Pecking: Application to Commercial Systems.” World’s Poultry Science Journal 69 :775–788. doi:https://doi.org/10.1017/S0043933913000809.
- Nørgaard-Nielsen, G. 1989. “The Effect of Access to Straw in Baskets on Feather Pecking in Laying Hens.” In Third European Symposium on Poultry Welfare, edited by J. M. Faure and A. D. Mills, 269–271. Tours, France: World Poultry Science Association.
- Nowland, W. J. 1978. Modern Poultry Management in Australia. Adelaide: Rigby Limited.
- Overmann, S. R. 1976. “Dietary Self-Selection by Animals.” Psychological Bulletin 83 (2): 218–235. doi:https://doi.org/10.1037/0033-2909.83.2.218.
- Peguri, A., and C. Coon. 1993. “Effect of Feather Coverage and Temperature on Layer Performance.” Poultry Science 72: 1318–1329. doi:https://doi.org/10.3382/ps.0721318.
- Rodenburg, T. B., M. M. Van Krimpen, I. C. De Jong, E. N. De Haas, M. S. Kops, B. J. Riedstra, R. E. Nordquist, et al. 2013. “The Prevention and Control of Feather Pecking in Laying Hens: Identifying the Underlying Principles.” World’s Poultry Science Journal 69 :361–374. doi:https://doi.org/10.1017/S0043933913000354.
- Rozin, P. 1976. “The Selection of Foods by Rats, Humans, and Other Animals.” Advances in the Study of Behavior 6: 21–76. doi:https://doi.org/10.1016/S0065-3454(08)60081-9.
- Rudkin, C. 1990. “Pecking and Feather Pecking in Growing Chickens.” Thesis for the degree of Master of Science, University of Queensland.
- SAS Institute Inc. 1991. SAS User’s Guide: Statistics, Version 6.10. Cary, NC: SAS Institute.
- Schreiter, R., K. Damme, E. von Borell, I. Vogt, M. Klunker, and M. Freick. 2019. “Effects of Litter and Additional Enrichment Elements on the Occurrence of Feather Pecking in Pullets and Laying Hens – A Focused Review.” Veterinary Medicine and Science 5: 500–507. doi:https://doi.org/10.1002/vms3.184.
- Van de Weerd, H. A., and A. Elson. 2006. “Rearing Factors that Influence the Propensity for Injurious Feather Pecking in Laying Hens.” World’s Poultry Science Journal 62: 654–664. doi:https://doi.org/10.1079/WPS2006119.
- Van Krimpen, M. M., R. P. Kwakkel, B. F. J. Reuvekamp, C. M. C. Van Der Peet-Schwering, L. A. Den Hartog, and M. W. A. Verstegen. 2005. “Impact of Feeding Management on Feather Pecking in Laying Hens.” World’s Poultry Science Journal 61 (4): 663–686. doi:https://doi.org/10.1079/WPS200478.
- Wiepkema, P. R. 1983. “Umwelt and Animal Welfare.” In Farm Animal Housing and Welfare, edited by S. H. Baxter, M. R. Baxter, and MacCormack, 45–50. Boston: Martinus Nijhoff.
- Wood-Gush, D. G. M., and C. G. Rowland. 1973. “Allopreening in the Domestic Fowl.” Revue De Comportement Animal 7: 83–91.
- Wood-Gush, D. G. M., A. Stolba, and C. Miller. 1983. “Exploration in Farm Animals and Animal Husbandry”. In Exploration in Animals and Humans, edited by J. Archer and L. Birke, 198–209. Wokingham: Van Nostrand Reinhold UK Co.