Abstract
The nuclear structures in different tissues of a number of dicotyledonous angiosperms show that, in certain species, nuclear repatternings due to the close association of different heterochromatic chromosomal regions occur in differentiating and differentiated cells. Chromocenter association plays a role in or is a marker of events that bring into play a large-scale control mechanism of the functional activity of the nucleus that is exploited in cell differentiation from its early stages. The somatic chromosome numbers of the genus Oroxylum indicum reported are 2n = 28, 2n = 30 and 2n = 38. The present work attempts to gives an account of the correct chromosome number in relation to chromocenters in mitotic and meiotic cells of Oroxylum indicum (L.) Vent.
Introduction
It is known that the interphase nucleus of many plant cells contains chromatin lumps (chromocenters). The chromocenter is characterized by several darkly stained bodies with an irregularly protruding rough surface (Tanaka Citation1971, Citation1977). The occurrence of compound chromocenters (Heitz Citation1929) and of changes in the number and size of chromatin lumps in cells from different tissues of a single plant was noticed many years ago (Delay Citation1946–1947, Citation1948) and has lately been investigated in detail in 67 plant species of different families (Ceccarelli et al. Citation1998). The results obtained by studying the nuclear structure in different tissues of a number of dicotyledonous angiosperms show that, in certain species, nuclear repatternings due to the close association of different heterochromatic chromosomal regions occur in differentiating and differentiated cells. Chromocenters are markers of events that bring into play a large-scale control mechanism of the functional activity of the nucleus that is exploited in cell differentiation from its early stages (Ceccarelli & Cionini Citation1993; Ceccarelli et al. Citation1998). Since chromocenters are believed to be composed primarily of heterochromatin/repetitive DNA (which has been postulated to shorten the cell cycle), it is likely that chromocenters modulate species size/biomass. Among closely related species, larger/more biomassive cultivars contained larger, more numerous, and more intensely staining chromocenters than smaller/less biomassive cultivars (Edelman & Lin Citation1996).
In all the species investigated, highly significant, positive correlations have been found between the number of chromatin lumps in a nucleus and the RNA synthetic activity in it. Repeated DNA sequences in the heterochromatin play a role in recognition processes between chromosomes (Ceccaralli et al. 1998). The extent of chromocenter association correlates positively with the number of chromosomes in the complement and the level of ploidy as well as with the maximum number of chromocenters in meristematic nuclei.
The occurrence of chromocenter association in a species may be linked to its phylogeny and position on an evolutionary scale. Indeed, the phenomenon was observed in all the species studied that belong to certain families, such as the Brassicaceae, and was never observed in the species of other families, such as the Fabaceae. Lack of reduction in the number of the chromocenters per nucleus in differentiated tissues of species of this latter family was also observed by Patankar & Ranjekar (Citation1984a, Citation1984b). Moreover, chromocenter association seems to accompany tissue differentiation in all species of the Magnoliidae, Caryophyllidae, Hamamelidae and Dilleniidae which are the subclasses at the base of the phyletic diagram of the Magnoliopsida (Cronquist Citation1988), while only 26% of those belonging to the Rosidae make use of this control mechanism for nuclear activity. The phyletic and evolutionary traits bring about the occurrence of chromocenter association in a species.
In the family Bignoniaceae, to which the plant Oroxylum indicum (L.) Vent. belongs, chromocenters have been karyomorphologically investigated in 11 species in Incarillea (Chen et al. Citation2004). In the present study the occurrence of chromocenters in root tip and in spore mother cell is studied in Oroxylum indicum (L.) Vent.
Shivnak, shyonak, sonpatha or midnight horror [Oroxylum indicum (L.) Vent., family Bignoniaceae] is a small deciduous, soft wooded tree. The tree measures up to 12 m in height and bears soft, light brown bark with corky lenticels. The tree has large, bilabiate flowers and long sword-shaped fruits. Its leaflet rachis is very soft and swollen at the junction of the branches and the leaflets are in 2–4 pairs, ovate, elliptic or acuminate in shape and glabrous in texture. Flowers are numerous, are arranged in large erect racemes, and are reddish purple outside and dull or pale yellow pinkish inside. Fruits are long woody capsules measuring 1 m in length. Each capsule contains numerous papery, flat, winged, light seeds. The fresh root bark is soft and juicy and creamy yellow to grayish in color, tasting sweet initially and bitter later. On drying, the bark shrinks, adheres closely to the wood and becomes faintly fissured (Gokhale & Bansal Citation2006).
Root and stem bark of this tree contain alkaloids and flavonoids of medicinal value used in the cure of many important diseases (Chen et al. Citation2003). The plant is used in many ayurvedic preparations such as shyonaka patpak, Bruhat pancha mulayadi kwath and chyawanprash, which are widely used for health care. Dichloromethane extract of the stem bark and root possesses antimicrobial, antifungal, anti-inflammatory and anticancerous properties (Yasodha et al. Citation2004). Owing to over-exploitation and indiscriminate collection, especially of whole plants bearing roots, this valuable tree has become vulnerable and it is feared that it may become endangered in India (Darshan & Ved Citation2003; Jayram & Prasad Citation2008).
The literature on the cytology of this plant is comparatively meager. Earlier cytological investigations (Raghavan & Venkatasubban Citation1940; Venkatasubban Citation1945; Darlington & Janaki Ammal Citation1945; Bir et al. Citation1982) refer mainly to the chromosome numbers of the various genera of the family Bignoniaceae. The somatic chromosome numbers of genus Oroxylum indicum reported are 2n = 28 (Ghatak Citation1956a, Citation1956b; Mangenot & Mangenot Citation1962), 2n = 30 (Venkatasubban Citation1944; Mehra & Bawa Citation1969) and 2n = 38 (Goldblatt Citation1989). The present work attempts to gives an account of the correct chromosome number with reference to presence of chromocenters in mitotic and meiotic cells of Oroxylum indicum (L.) Vent.
Materials and methods
For cytological investigations root tips (5–7 mm) of germinating seeds were treated with 1% α bromonaphthalene for 4 hrs at 4°C, and subsequently fixed in Carnoy’s fluid II (glacial acetic acid: chloroform: ethanol 1:3:6) for 1 hour (Goldblatt Citation1989).
The plant tissue was softened through hydrolysis with 10% HCl for 10 min at 60°C. The structure and behavior of the chromosomes was studied by staining with 2% FLP orcein (formo:lacto:propiono orcein). The stain was prepared by dissolving 2 gm orcein powder in equal volumes of formic acid:lactic acid:propionic acid (Jackson 1983).
The material was squashed in a drop of 45% acetic acid. Photomicrographs were taken through a Nikon research microscope with photographic attachment using 45× and 100× (oil immersion) objectives. The data presented here are based on the observation of a total 1065 cells (705 root tip and 360 spore mother cells). Column charts have been used to display the trend of contribution of each value of two parameters, i.e. MI (mitotic index) and CI (chromocentric index). MI and CI were calculated using the following formulae.
Results and discussion
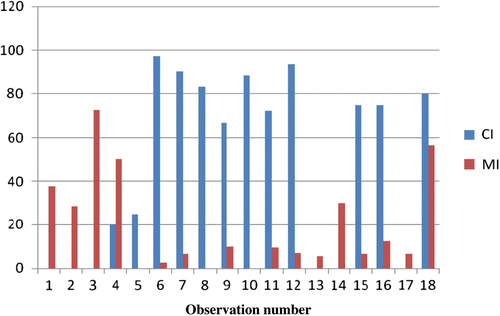
Chromocenters, the dark staining heterochromatic bodies, were observed in all dividing cells during mitosis and meiosis. depict the mean chromocenter numbers in dividing cells of O. indicum. Chromocenter number in this species ranged from 0 to a maximum of 14 (Figures –6). On the basis of cytological observations during mitosis and meiosis, the average number of chromocenters was 7.53. The base number of family Bignoniaceae as reported earlier was 7 (Goldblatt & Gentry Citation1979). The somatic chromosomes seen during mitosis in the root tip cells (Figures –12) and during meiosis in the spore mother cell (Figures –16) in the present work offered the first visible evidence of the chromocenters and chromosome number in O. indicum. A comparison of the values obtained by measuring MI (mitotic index) and CI (chromocentric index) indicates a definite relationship between them, and thus supports a correlation between chromosome number and number of chromocenters (Figures , ). On the basis of the present study the somatic chromosome number of genus Oroxylum indicum supports a chromosome count of 2n = 28 (Ghatak Citation1956a, Citation1956b; Mangenot & Mangenot Citation1962). Earlier studies demonstrated that the amount and the distribution of constitutive heterochromatin as inferred from the chromocenter counts in the interphase nuclei are under the control of the genotype and related with homo- and heterozygosity.
Table 1. Comparative analysis of chromocentric and mitotic index in dividing cells during mitosis in Oroxylum indicum L. (Vent.).
Table 2. Comparative observation of chromocentric and mitotic index in dividing cells during meiosis in Oroxylum indicum L.Vent.
Figures 1–16 (Colour online) Cytological observations in Oroxylum indicum L. Vent. (1) Chromocenter 7, the base chromosome number of family Bignoniaceae. (2–6) Different numbers and orientations of chromocenters. (7–12) Chromosomes in different stages of mitosis. (13) Seven bivalents showing a specific type of orientation during meiosis. (14–16) Chromosomes in different stages of meiosis.
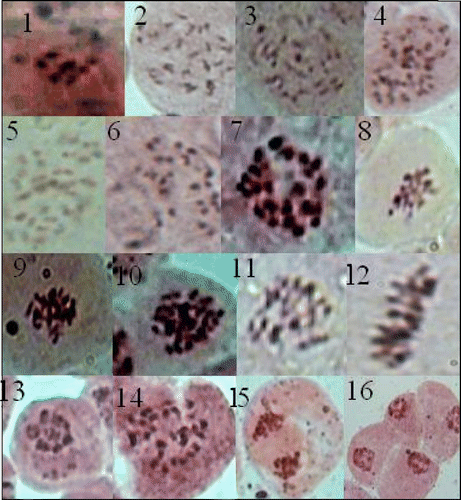
Figure 17 (Colour online) Column chart comparing values between MI and CI in dividing root tips in Oroxylum indicum L. Vent.
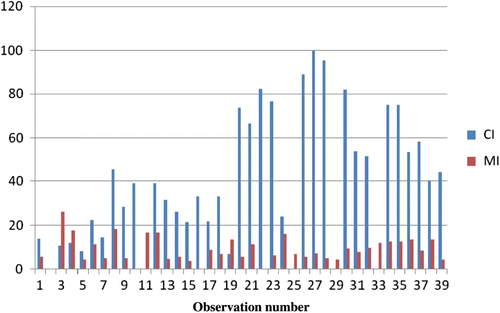
Figure 18 (Colour online) Column chart comparing values between MI and CI in spore mother cells in Oroxylum indicum L. Vent.
The diploid chromosome number in the present study has been observed to be 2n = 28. The most abundant stages among the dividing cells in the present work were metaphase and anaphase. The size of the chromosome was too small (approx 1.12–3.67 μm) to observe its structure.
The chromocenter, a heavily stainable heterochromatic area with highly repetitive DNA, is a reliable taxonomic trait and a good marker to establish the accurate number of chromosomes and the phylogeny of a genus, and to determine the relationship between closely related species (Gajardo et al. Citation2001, Zhiyun et al. Citation2003, Zhang et al. Citation2010). A medicinally important tree of family Bignoniaceae, Oroxylum indicum has presented a confusing picture due to different reported chromosome numbers, i.e. 2n = 28 (Ghatak Citation1956a, Citation1956b; Mangenot & Mangenot Citation1962), 2n = 30 (Venkatasubban Citation1944; Mehra & Bawa Citation1969) and 2n = 38 (Goldblatt Citation1989). The establishment of correct chromosome number in plants is important for determining polyploidy caused by different reasons, e.g. mutation and in vitro regeneration. The chromosome number in this species has remained controversial for a long time. According to Goldblatt & Gentry (Citation1979), the chromosome number in O. indicum is n = 14, and the one reported by Venkatasubban (Citation1944) and Mehra & Bawa (Citation1969) (n = 15), is erroneous. However, Goldblatt (Citation1989) reported 2n = 38 for this species.
Conclusion
The present study correlates the average number of chromocenters in the species O. indicum with the mitotic index and gives an accurate chromosome number of 2n = 28.
Acknowledgment
The authors are thankful to the Department of Science and Technology (DST), New Delhi, India for financial assistance to M. Gokhale (Project No. DST WOS-A/SR/LS-07/2009).
References
- Bir , SS , Gill , BS and Bedi , YS . 1982 . Cytopalynological studies on some members of Bignoniaceae from India . Indian J Bot , 5 ( 1 ) : 7 – 12 .
- Ceccarelli , M and Cionini , PG . 1993 . Tissue-specific nuclear repatterning in plant cells . Genome , 36 : 1092 – 1098 .
- Ceccarelli , M , Morosi , L and Cionini , PG . 1998 . Chromocenter association in plant cell nuclei: determinants, functional significance and evolutionary implications . Genome , 41 : 96 – 103 .
- Chen , L , Games , DE and Jones , J . 2003 . Isolation and identification of four flavonoid constituents from the seeds of Oroxylum indicum by high speed counter current chromatography . J Chromatogr A , 988 : 95 – 105 .
- Chen , ST , Zhou , ZK , Guan , KY and Masashi , N . 2004 . Karyomorphology of Incarvillea (Bignoniaceae) and its implications in distribution and taxonomy . Bot J Linn Soc , 144 ( 1 ) : 113 – 121 .
- Cronquist A. 1988. The evolution and classification of flowering plants. 2nd ed. New York: The New York Botanical Garden. 555 p.
- Darlington CD, Janaki Ammal EK. 1945. Chromosome atlas of cultivated plants, London: George Allen and Unwin.
- Darshan , S and Ved , DK . 2003 . A balanced perspective for management of Indian Medicinal Plants . Indian For , 129 ( 2 ) : 275 – 288 .
- Delay C. 1946–1947. Recherches sur la structure des noyaux quiescents chez les Phanérogames. Rev Cytol Cytophys Vég. 9:169–222.
- Delay , C . 1948 . Recherches sur la structure des noyaux quiescents chezles Phanérogames . Rev Cytol Cytophys Vég , 10 : 103 – 228 .
- Edelman , JR and Lin , YJ . 1996 . Relationship of chromocenter number, size, and density to plant biomass . Caryologia , 49 ( 3–4 ) : 207 – 218 .
- Gajardo , G , Beardmore , JA and Sorgeloos , P . 2001 . International study on Artemia LXII. Genomic relationships between Artemia franciscana and A. Persimilis, inferred from chromocenter numbers . Heredity , 87 : 172 – 177 .
- Ghatak J. 1956a. Oroxylum Vent. In: Kumar V, Subramanium B, editors. Chromosome Atlas of Flowering Plants of the Indian Subcontinent. Botanical Survey of India. P-8, Brabourne Road, Calcutta-700001. I. p. 93.
- Ghatak , J . 1956b . A contribution to the life history of Oroxylum indicum Vent Proceedings . Indian Acad Sci B , 43 ( 1 ) : 72 – 87 .
- Gokhale , M and Bansal , YK . 2006 . An avowel of importance of an endangered tree Shivnak (Oroxylum indicum (L) Vent) . Nat Prod Radiance , 5 ( 2 ) : 112 – 114 .
- Goldblatt , P . 1989 . Miscellaneous chromosome count in Asteraceae, Bignoniaceae, Proteaceae and Fabaceae . Ann MO Bot Gard , 76 ( 4 ) : 1186 – 1188 .
- Goldblatt , P and Gentry , AH . 1979 . Cytology of Bignoniaceae . Botaniska Notiser , 132 : 475 – 482 .
- Heitz , E . 1929 . Heterochromatin, Chromocentren . Chromosomen. Berichte der Dentschen Botanischen Gesellschaft , 47 : 274 – 284 .
- Jackson , RC . 1973 . Chromosomal evolution in Haplopappus gracilis: a centric transposition race . Evolution , 27 : 243 – 256 .
- Jayram , K and Prasad , MNV . 2008 . Genetic diversity in Oroxylum indicum (L) Vent. (Bignoniaceae) a vulnerable medicinal plant by random amplified polymorphic DNA marker . Afr J Biotechnol , 7 ( 3 ) : 254 – 262 .
- Mangenot , S and Mangenot , G . 1962 . Enquete surles nombres chromosomiques dans une collection d’especes tropicales . Rev Cytol Biol Veg , 25 : 411 – 447 .
- Mehra , PN and Bawa , KS . 1969 . Chromosomal evolution in tropical hardwoods . Evolution , 23 : 466 – 481 .
- Patankar , S and Ranjekar , PK . 1984 . Interphase nuclear structure and heterochromatin in Phaseolus plant species . Plant Cell Rep , 3 : 130 – 133 .
- Patankar , S and Ranjekar , PK . 1984 . Condensed chromatin and its underreplication during root differentiation in Leguminosae . Plant Cell Rep , 3 : 250 – 253 .
- Raghavan , TS and Venkatasubban , KR . 1940 . Studies in the Bignoniaceae . J Indian Bot Soc , 19 : 293 – 298 .
- Tanaka , R . 1971 . Types ofresting nucei in Orchidaceae . Bot Mag Tokyo , 84 : 118 – 122 .
- Tanaka , R . 1977 . “ Recent karyotype studies ” . In Plant Cytology , Edited by: Ogawa , K , Kurozumi , K , Koike , S and Sato , S . Tokyo : Asakura Shoten .
- Venkatasubban KR. 1944. Cytological studies in Bignoniaceae. Annamalai Univ Publ. 1, 2 and 3:1–207.
- Venkatasubban , KR . 1945 . Cytological studies in Bignoniaceae . Proc. Indian Acad Sci , 21B : 77 – 92 .
- Yasodha , R , Ghosh , M , Santan , B and Gurumurthi , K . 2004 . Importance of biotechnological research in tree species of Dashmula . Indian Forester , 130 : 9 – 88 .
- Zhang X-H, Jaime A, Da Silva T, Ma G. 2010. Karyoyype analysis of Santalum album L. Caryologia. 63(2):142–148.
- Zhiyun , Y , Tingshuang , Y , Heng , L and Gong , X . 2003 . A cytological study in three species of Colocasia (Araceae) from Yunnan . Caryologia , 56 ( 3 ) : 323 – 327 .