Abstract
In this study, the G-bands and nucleolus organizer regions (NORs) of the 2n = 60 chromosomal form of Nannospalax nehringi from Central Anatolia (Konya) are presented. The karyotype possesses nine pairs of submetacentric and subtelosentrics and 20 pairs of acrocentric autosomes gradually decreasing in size (fundamental number = 80 and autosomal fundamental number = 76). NORs were localized in the telomeric regions of the short arms of three large pairs and one medium-sized pair of subtelocentric autosomes (nos. 1, 3, 4, 9).
Introduction
The family Spalacidae is composed of fossorial mole rats which are highly adapted to subterranean life. The systematics and taxonomy of extant species of mole rats are still controversial. Gromov and Baranova (Citation1981) stated that the subfamily Spalacinae is represented by two genera: Nannospalax (Palmer 1903) and Spalax Guldenstaedt, 1770. However, Musser and Carleton (Citation2005) accepted Nannospalax as a synonym of the genus Spalax and treated the family as monogeneric, including only the nominative genus. The authors stated that Spalax leucodon Nordmann, 1840, S. nehringi (Satunin 1898) and S. ehrenbergi (Nehring 1898) are distributed in Turkey. Kryštufek and Vohralik (Citation2009) and Kryštufek et al. (2012) proposed the name xanthodon instead of nehringi and indicated that three morphospecies, Spalax leucodon, S. xanthodon (Nordmann, 1840) and S. ehrenbergi are distributed in Turkey. Both Sözen et al. (Citation2011) and Matur et al. (Citation2011) decided to use N. nehringi instead of S. leucodon. However, Arslan and Bölükbaş (Citation2010), Arslan et al. (Citation2010), Arslan et al. (Citation2011a) and Arslan et al. (Citation2011b) adopted both N. nehringi and N. xanthodon for the mole rats distributed in Central Anatolia.
The genus Nannospalax is distinguished from Spalax with respect to low diploid chromosome number (2n) and fundamental number (NF) as well as possessing acrocentrics in the chomosome set (Kryštufek and Vohralik 2009; Németh et al. 2009). Contrary to Musser and Carleton (2005) and Kryštufek and Vohralik (2009), we attributed the mole rats distributed in Turkey to the genus Nannospalax, having mainly acrocentric chromosomes (Aşan and Yağcı Citation2008; Coşkun et al. Citation2010, Arslan and Bölükbaş Citation2010; Arslan et al. Citation2010; Arslan et al. (2011a,b).; Matur et al. Citation2011; Sözen et al. Citation2011).
To date, more than 50 chromosomal forms have been described for the family Spalacidae distributed in the Palaearctic region, and of them 30 forms were only recorded from Turkey. However, the exact distributional areas of these chromosomal forms are still not known. New cytotypes are being discovered continuously with as more specimens are examined in Turkey (Nevo et al. Citation1995; Sözen Citation2004; Sözen et al. Citation2006).
Recently, Sözen et al. (Citation1999, Citation2006), Kankılıç et al. (Citation2007), Ivanitskaya et al. (Citation2008), Arslan and Bölükbaş (Citation2010), Arslan et al. (2011a,b) have examined the conventional stained and banded karyotypes of specimens from Central Anatolia. All the authors accepted that the diploid chromosome number of examined specimens was 60 (except the Kırıkkale population with 2n = 54) and the autosomal fundamental number ranged between 68 and 80.
The aim of this study is to make a contribution to the karyomorphology of the 2n = 60 cytotype of mole rats distributed in Central Anatolia with regard to presenting the G-banding of the chromosomes and nucleolus organizer region (NOR)-bearing chromosome number.
Material and methods
Five specimens (3 ♂♂, 2 ♀♀) were captured from Konya, Cihanbeyli (38°′37′ N, 32°54′ E) and Kulu (39°05′ N, 33°04′ E) in Central Anatolia. Chromosome preparations were obtained from bone marrow cells according to the technique of Ford and Hamerton (Citation1956). The G-banding was achieved according to technique of Seabright (Citation1971). The location of NORs in the chromosomes was determined using the method of Howell and Black (Citation1980). Chromosomes were classified according to Levan et al. (Citation1964). At least 20 well-spread and banded metaphase plates were photographed and arranged to determine the diploid chromosome number (2n), autosomal fundamental number (NFa) and fundamental number (NF). All stuffed skins and slides are deposited at the Vocational School of Pazaryeri, University of Bilecik and Faculty of Science and Arts, University of Kırıkkale, respectively.
Results
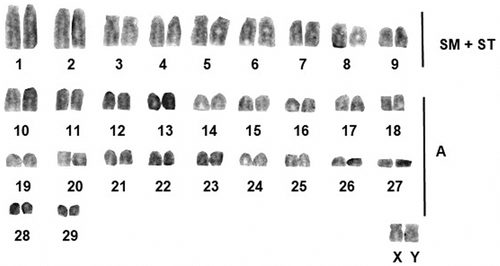
The karyotype had 2n = 60, NF = 80 and NFa = 76. The chromosome set consisted of seven pairs of subtelosentric pairs (nos. 1–3, 5–8), two submetacentric pairs (nos. 4, 9) and 20 pairs of acrocentric pairs (nos. 10–29), gradually decreasing in size. The X chromosome was medium sized submetacentric while the Y was small acrocentric. No secondary constriction or heteromorphic chromosome pair are detected in the examined karyotypes (Figure ).
Figure 1 Conventionally stained karyotype of Nannospalax nehringi.
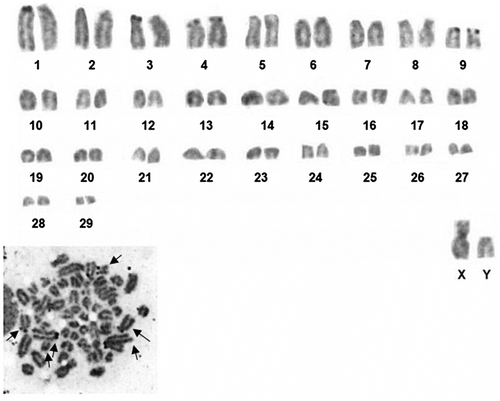
Chromosome shapes of Nannospalax nehringi were determined and are given in Table . G-banding karyotype of the 2n = 60 is given in Figure . NORs are located in the telomeric regions of the short arms of three large pairs (nos. 1, 3, 4) and one medium-sized subtelocentric pair (no. 9). Active Ag-NORs on homologous chromosomes exhibited heteromorphism in some cells (autosome pair nos. 1 and 3) (Figure ).
Table 1. Chromosome classification in Nannospalax nehringi according to Levan et al. (Citation1964).
Figure 2 G-banded karyotype of the 2n = 60 cytotype of Nannospalax nehringi.
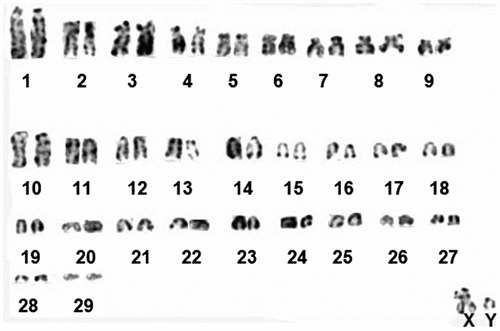
Figure 3 Silver stained metaphase and karyotype of the 2n = 60 cytotype of Nannospalax nehringi (arrows indicate the NOR-bearing chromosomes).
Discussion
According to Nevo et al. (Citation1995), the karyotypic evolution in the genus Spalax is caused by Robertsonian mutations. Therefore, the spalacids represented a remarkable adaptive ecological speciation, caused by these mutations, in its distributional area.
So far, about 150 populations of mole rats have been karyotyped in Turkey (Coşkun et al. Citation2010). The diploid chromosome number and fundamental number of the genus Nannospalax showed variation from 36 to 62 and 66 to 98, respectively (Sözen et al. Citation2006; Ivanitskaya et al. Citation2008; Ulutürk et al. Citation2009). The 2n = 60 chromosomal form is the most common cytotype, although 2n = 36 and 2n = 62 cytotypes are the extreme ones (Kankılıç et al. Citation2007; Ivanitskaya et al. Citation2008; Arslan and Bölükbaş Citation2010; Arslan et al. (2011b)). Despite the numerous conventionally stained karyotypes prepared for the genus, few banded karyotypes are reported from Turkey (Gülkaç and Küçükdumlu Citation1999; Ivanitskaya et al. Citation1997, Citation2008; Arslan and Bölükbaş Citation2010; Arslan et al. (2011 a,b). Gülkaç and Küçükdumlu (Citation1999) examined NOR distribution in the 2n = 60 chromosomal form from Malatya province and active NORs are recorded on the telomeric regions of three pairs of large subtelocentrics. Ivanitskaya et al. (Citation2008) stated that the 2n = 60 cytotype of Nannospalax nehringi (attributed as S. leucodon) possessed two different kinds of karyotype structure; 2n = 60W (wide distribution cytotype with NORs in the telomeric regions of the chromosomes numbered 3, 4, 5, 6 and 13) and 2n = 60R (rare distribution cytotype with NORs in the telomeric regions of the chromosomes numbered 6, 8, 12, 13 and 16) in Turkey. Ag-NORs on homologous chromosomes exhibited heteromorphism in the 2n = 60W cytotype. Arslan and Bölükbaş (Citation2010) determined active Ag-NORS in the telomeric regions of four biarmed chromosomes. Ag-NORs on one of the homologous chromosomes exhibited heteromorphism. However in addition, Arslan et al. (2011b). recorded Ag-NORs in the telomeric regions of four biarmed autosomal pairs of 2n = 54 chromosomal form from Central Anatolia.
Recently, Cihanbeyli (Konya) karyotypes of specimens of the 2n = 60 cytotype were examined by Kankılıç et al. (Citation2007) and Arslan et al. (2011a). All authors considered that the karyotype had 2n = 60, NF = 80 and NFa = 76 with nine pairs of sub- and subtelocentric autosomes and 20 pairs of acrocentric autosomes. However, discrepancy was found in the shape of the X chromosome in both studies. Kankılıç et al. (Citation2007) stated that the X chromosome was metacentric and the Y subtelocentrics, while Arslan et al. (2011a). recorded the X chromosome as submetacentric. Unfortunately, no male specimen was examined by the latter authors. Arslan et al. (2011a) stated that the 2n = 60 chromosomal form differed with respect to the shape and size of the Y chromosome. However, we determined the X chromosome as submetacentric and the Y as acrocentric in this study.
Arslan et al. (2011a) observed Ag-NORs in the telomeric regions of the short arms of four biarmed autosomes (nos. 3, 4, 5 and 6). In addition, we detected an active NOR in the telomeric region of one medium-sized metacentric chromosome (no. 9) in this study. The authors did not mention any active Ag-NORs on this autosome pair. Furthermore, the dissimilarity between NOR-bearing subtelocentric autosomes (no. 6 in Arslan et al. (2011a) and no. 1 in this study) in both studies, was probably due to the different methods used for arrangement of the chromosomes by the authors.
Consequently, despite the different shape of NOR-bearing chromosomes, the number of Ag-NORs is recorded as four from Central Anatolia (Arslan and Bölükbaş Citation2010 a; Arslan et al (2011a,b); this study). For determining intra- and interspecific chromosomal differences the Ag-NOR banding is as useful as the other banding techniques as stated by Arslan et al. (2011a).
References
- Arslan , A and Bölükbaş , F . 2010 . C-heterochromatin and NORs distribution of mole rat, Nannospalax xanthodon from Aksaray . Turkey. Caryologia. , 63 ( 4 ) : 398 – 404 .
- Arslan , E , Gülbahçe , E , Arıkoğlu , H , Arslan , A , Bužan , EV and Kryštufek , B . 2010 . Mitochondrial divergence between three cytotypes of the Anatolian Mole rat, Nannospalax xanthodon (Nordmann, 1840) . Zool. Middle East. , 50 : 27 – 34 .
- Arslan , A , Akan , Ş and Zima , J . 2011a . Variation in C-heterochromatin and NOR distribution among chromosomal races of mole rats (Spalacidae) from Central Anatolia . Turkey. Mamm Biol. , 76 ( 1 ) : 28 – 35 .
- Arslan , A , Toyran , K , Gözütok , S and Yorulmaz , T . 2011b . C-and NOR stained karyotypes of mole rat, Nannospalax xanthodon (2n=54) from Kırıkkale . Turkey. Turk J Biol. , 35 ( 6 ) : 655 – 661 .
- Aşan , N and Yağcı , T . 2008 . Karyotype and hair scale structure of Nannospalax leucodon (Nordmann, 1840) from central Anatolia (Rodentia: Spalacidae) . Turk J Zool. , 32 ( 2 ) : 125 – 130 .
- Coşkun , Y , Ulutürk , S and Kaya , A . 2010 . Karyotypes of Nannospalax (Palmer, 1903) populations (Rodentia: Spalacidae) from Central-Eastern Anatolia . Turkey. Hystrix It J Mamm. , 21 ( 1 ) : 89 – 96 .
- Ford , CE and Hamerton , JL . 1956 . A colchicine-hypotonic-citrate squash sequence for mammalian chromosomes . Stain Technol. , 31 : 247 – 251 .
- Gromov , IM and Baranova , GI . 1981 . Catalogue of mammals in USSR , Leningrad : Nauka .
- Gülkaç , MD and Küçükdumlu , İ . 1999 . Variation in the nucleolus organizer regions (NORs) in two mole rat species (Spalax leucodon and Spalax ehrenbergi) . Turk J Biol. , 23 ( 2 ) : 153 – 158 .
- Howell , WM and Black , DA . 1980 . Controlled silver staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method . Experientia , 36 : 1014 – 1015 .
- Ivanitskaya , E , Coşkun , Y and Nevo , E . 1997 . Banded karyotypes of mole rats (Spalax, Spalacidae, Rodentia) from Turkey . J Zool Syst Evol Res. , 35 ( 4 ) : 171 – 177 .
- Ivanitskaya , E , Sözen , M , Rashkovetsky , L , Matur , F and Nevo , E . 2008 . Discrimination of 2n=60 Spalax leucodon (Spalacidae, Rodentia) in Turkey by means of classical and molecular cytogenetic techniques . Cytogenet. Genome Res. , 122 ( 2 ) : 139 – 149 .
- Kankılıç , T , Kankılıç , T , Çolak , R , Çolak , E and Karataş , A . 2007 . Karyological comparision of populations of the Spalax leucodon Nordmann, 1840 superspecies (Rodentia: S palacidae) in Turkey . Zool Middle East. , 42 : 15 – 24 .
- Kryštufek B, Vohralik V. 2009. Mammals of Turkey and Cyprus. Rodentia II. Cricetinae, Muridae, Spalacidae, Calomyscidae, Capromyidae, Hystricidae, Castoridae. Koper: Knjižnica Annales Majora, University of Primorska Science and Research Centre.
- Krystufek B, Ivanitskaya E, Arslan A, Arslan E, Buzan EV. 2012. Evolutionary history of mole rats (genus Nannospalax) inferred from mitochondrial cytochrome b sequence. Biological Journal of the Linnean Society. 105(2): 446–455.
- Levan , AK , Fredga , K and Sandberg , AA . 1964 . Nomenclature for centromere position on chromosomes . Hereditas. , 52 : 201 – 220 .
- Matur , F , Çolak , F , Sevindik , M and Sözen , M . 2011 . Chromosome differentiation of four 2n=50 chromosomal forms of Turkish mole rat Nannospalax nehringi . Zool Sci. , 28 ( 1 ) : 61 – 67 .
- Musser GG, Carleton MD. 2005. Rodentia: Myomorpha: Muroidea: Spalacidae: Spalacinae. In: Wilson DE, Reeder DM, editors. Mammal species of the world. A taxonomic and geographic reference. 3rd ed. Baltimore: The John Hopkins University Press. p. 917–922.
- Németh , A , Révay , T , Hegyeli , Z , Farkas , J , Czabán , D , Rὀzsás , A and Csorba , G . 2009 . Chromosomal forms and risk assessment of Nannospalax (Superspecies leucodon) (Mammalia: Rodentia) in the Carpathian Basin . Fol Zool. , 58 ( 3 ) : 349 – 361 .
- Nevo , E , Filipucci , MG , Redi , CD , Simson , S , Heth , G and Beiles , A . 1995 . Karyotype and genetic evolution in speciation of subterranean mole rats of the genus Spalax in Turkey . Biol J Linn Soc. , 54 ( 3 ) : 203 – 209 .
- Seabright , MA . 1971 . Rapid banding technique for human chromosomes . Lancet. , 2 : 971 – 972 .
- Sözen , M . 2004 . A karyological study on subterranean mole rats of the Spalax leucodon Nordmann, 1840 superspecies in Turkey . Mamm. Biol. , 69 : 420 – 429 .
- Sözen , M , Çolak , E , Yiğit , N , Özkurt , Ş and Verimli , R . 1999 . Contributions to the karyology and taxonomy of the genus Spalax Güldenstaedt, 1770 (Mammalia: Rodentia) in Turkey . Zeitschrift fur Sauget. , 64 : 210 – 219 .
- Sözen , M , Matur , F , Çolak , E , Özkurt , Ş and Karataş , A . 2006 . Some karyological records and a new chromosomal form for Spalax (Mammalia: Rodentia) in Turkey . Fol Zool. , 55 ( 3 ) : 247 – 256 .
- Sözen , M , Çataklı , K , Eroğlu , F , Matur , F and Sevindik , M . 2011 . Distribution of chromosomal forms of Nannospalax nehringi (Satunin, 1898) (Rodentia: Spalacidae) in Çankırı and Çorum provinces . Turkey. Turk J Zool. , 35 ( 3 ) : 367 – 374 .
- Ulutürk , S , Coşkun , Y and Arıkan , H . 2009 . A comparative karyological and serological study on two populations of Nannospalax nehringi (Satunin 1898) from Turkey . North-West J Zool. , 5 ( 2 ) : 349 – 356 .