Abstract
A putatively endemic infraspecific taxon of the Hieracium villosum group from the northern Apennines was investigated from nomenclatural, morphological and karyological points of view. H. villosum Jacq. var. zangherii Vignolo-Lutati proved to be invalidly described, and for its morphological features was referred to H. dentatum Hoppe subsp. xanthostylophorum Furrer & Zahn, a taxon hitherto only known from Abruzzo (Central Apennines). Karyological investigations resulted in the first report for an Italian specimen belonging to the collective species H. dentatum. H. dentatum subsp. xanthostylophorum proved to be tetraploid, with a formula 2n = 4x = 36 = 2M + 14m + 18sm + 2st. Chromosomes were generally of large size and showed a low degree of polymorphism in their shapes. The karyological parameters of H. dentatum subsp. xanthostylophorum have been compared with some available karyological parameters from the literature, belonging to related entities of Hieracium subgen. Hieracium. The Italian Hieracium subgen. Hieracium populations proved to have the most asymmetric karyotypes with respect to other European populations. H. dentatum subsp. xanthostylophorum showed an intermediate karyotype asymmetry between Hieracium subgen. Hieracium populations of the Balkan Peninsula and those of southern Tuscany. From a broader point of view, this study confirms in the northern Apennines a trend pointed out in other central and southern European mountain areas: the core taxon (H. villosum) is surrounded by tetraploid apomictic lineages that show some morphological characters of other Hieracium species; in the case of H. dentatum subsp. xanthostylophorum (= H. villosum “var. zangherii”), characters of H. bifidum.
Introduction
While studying the flora and vegetation of the National Park “Foreste Casentinesi, M. Falterona, Campigna” (Viciani and Agostini Citation2009; Viciani et al. Citation2010), located in the Northern Apennines between Tuscany and Emilia Romagna (Figure ), we collected several specimens of a Hieracium subgen. Hieracium (villosum/dentatum group) difficult to identify (Figure ). It is well known that the genus Hieracium L. is one of the taxonomically most intricate groups of vascular plants, and that in Hieracium subgen. Hieracium group agamospermy is widespread (Chrtek et al. Citation2004). Moreover, according to Bedini et al. (Citation2012a), Hieracium is one of the least studied genera of the Italian flora, on karyological grounds. This is further confirmed when one considers just the taxa endemic to Italy (Bedini et al. Citation2012b). The section to which our specimens could be referred is the sect. Villosa; H. villosum Jacq. belongs, together with its intermediate taxa, to one the most interesting groups of mountain hawkweeds in central and southern Europe (Severa and Chrtek Citation2002; Chrtek et al. Citation2004). In the studied territory, some specimens corresponded from a morphological point of view to an endemic taxon mentioned for this area as H. villosum Jacq. var. zangherii Vignolo-Lutati in Zangheri (Citation1966). To check the actual distinctness of such an infraspecific taxon, karyological and morphological analyses have been carried out on fresh collected material and original herbarium specimens. The results are reported below.
Figure 1 Localization in Italy of the study area.
Figure 2 The plant in the wild (a whole plant with flowers; inset: a fruiting capitulum).

Materials and methods
Specimens
During the period 2008–2010, several specimens of plants morphologically corresponding to the original material and the description of H. villosum var. zangherii were collected in the locus classicus (northern slopes of Mt. Falco, Northern Apennines, altitude about 1600 m asl., 1 × 1 km UTM grid: 32T, 717 E, 4861 N). Mature seeds were used for the karyological analyses (seeds were present also in the original specimen but they proved not to be viable).
We considered the original specimen (Figure ) [M. Falco (M. Falterona) dirupi volti a settentrione, alt. 1600 m., 8 agosto 1934, leg. P. Zangheri, det. F. Vignolo-Lutati in 1953, Herb. Zangherii n. 13779, VER] and all the other original allied Hieracium specimens (H. villosum, H. dentatum Hoppe, H. bifidum Hornem.) coming from the same area and belonging to Herbarium Zangherii, hosted in the herbarium of the Verona Natural History Museum (VER). We checked the main macro-morphological characters with regard to recent research on Italian Peninsula Hieracium (Gottschlich Citation2005, Citation2009a, Citation2009b, Citation2011).
Figure 3 The original specimen on which Hieracium villosum var. zangherii was invalidly described.

Karyological analyses
We took into consideration several recent cytogenetic analyses concerning other taxa of genus Hieracium and related genera (Coşkuncelebi and Hayirlioğlu-Ayaz Citation2006; Ilnicki et al. Citation2010; Ozcan et al. Citation2011) and other Asteraceae (Kiran et al. Citation2012; Meng et al. Citation2012).
Karyological analyses were obtained from somatic mitoses taken directly from the first phases of the developing embryo root tips after seed germination. After a pre-treatment of about 3 hours in 8-hydroxichinoline saturated solution at room temperature, the material was fixed in Carnoy’s fluid, second fixative solution (ethanol and acetic acid 3:1) (Johansen Citation1940) for several hours, also used as a preservative solution in the test tubes stocked at 5°C temperature. Afterwards, the material was washed in distilled water and then hydrolyzed in HCl 1N at 60°C for 5 minutes. Finally, after HCl removing and after briefly washing the tips in distilled water, they were stained with 40% water solution of lacto-propionic-orceine (Dyer Citation1979) for 18–48 hours at room temperature.
Karyological investigations were carried out on mitotic metaphase plates of meristematic cells, with the technique of fresh squashes of root tips (Dyer Citation1979): a very small piece of stained meristematic tissue was placed on a slide in a drop of 45% acetic acid, a clean coverslip was lowered over the drop and then gently pressured with the tip of a mounted needle until all the cells were dispersed in a large squash. Chromosome counts were made both during direct observations with the microscope, and on enlarged images of the original micrographs. Images were recorded with a microscope ZEISS Axiophot and digital photocamera Canon G5 connected to a personal computer. Measurements and values were processed in order to obtain chromosome ordering and homologue recognition, idiogram, karyotype formula and chromosome nomenclature proposed by Levan et al. (Citation1964). Several other caryological parameters were calculated: [TF%] index by Huziwara (Citation1962), centromeric index [As.k%] (Arano and Saito Citation1980), asymmetry [A] by Watanabe et al. (Citation1999), the intra-[A1] and inter-chromosomal [A2] asymmetry indices proposed by Romero Zarco (Citation1986), and also more recently proposed indices as the CVCL, CVCI (Paszko Citation2006), or MCA (Peruzzi and Eroğlu Citation2013) (Table ). Some of these parameters have been recently criticized (for an updated review see Peruzzi and Eroğlu Citation2013). The aim of the performed comparative analysis was to have more karyotype information for comparing among populations or taxa supposed to be more or less closely related. Morphological or biochemical characters depend on the expression of some genes, but chromosomal characters do not depend on gene expressions: karyotype is nothing less than the “shape” of the genome itself (Guerra Citation2012), adding a priori information, so that karyological analyses are very important in biosystematic research.
Results
Nomenclature and taxonomy
According to art. 39 ICBN (McNeill et al. Citation2012) H. villosum var. zangherii is invalidly described (only with an Italian and not a Latin description). Despite this, the checks both on fresh material (Figure ) and on the original specimen demonstrated that the plants must be attributed not to an infraspecific taxon of H. villosum but to H. dentatum, a taxon considered as an intermediate between H. villosum and H. bifidum (Pignatti Citation1982; Severa and Chrtek Citation2002). Also H. villosum, H. pilosum Froel. and H. bifidum are present in the same restricted area where H. villosum var. zangherii (= H. dentatum) lives. The interpretation of H. villosum var. zangherii as belonging to H. dentatum is due to the fact that the original specimen and the recently collected material have the following morphological main characters: basal leaves +/– petioled; stem leaves only rounded at the base and not more or less amplexicaul as in H. villosum; involucrum less hairy than in H. villosum. Because of the broadly spathulate basal leaves the plant can be assigned to H. dentatum subsp. xanthostylophorum Furrer & Zahn, a taxon hitherto only known from Abruzzo (Gottschlich Citation2009a, there erroneously mentioned as “xanthostylomorphum”).
Figure 4 A recent specimen of Hieracium dentatum subsp. xanthostylophorum (= “H. villosum var. zangherii”).

Karyological analyses
Hieracium dentatum subsp. xanthostylophorum (= “H. villosum var. zangherii”) – 2n = 36 (Figure , 5b). This is the first report for an Italian specimen belonging to the collective species H. dentatum (Bedini et al. Citation2010).
Figure 5 (a) Photomicrograph of mitotic metaphase plate of Hieracium dentatum subsp. xanthostylophorum (= “H. villosum var. zangherii”); (b) karyogram.
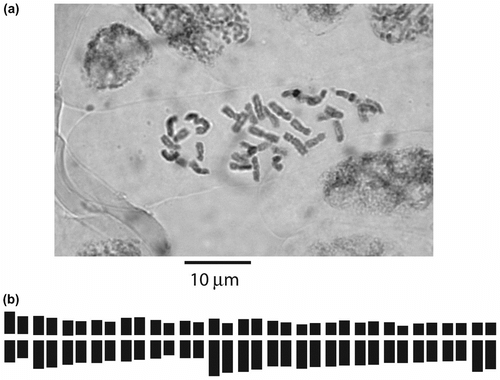
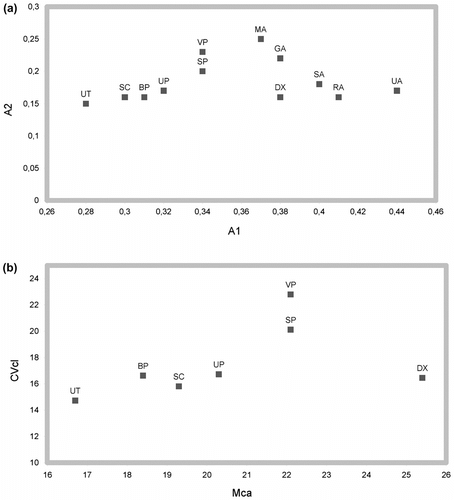
The count corresponding to the tetraploid number 2n = 36 (x = 9) was made on 12 mitotic metaphase plates, but the karyotype is related to the selected micrograph for Figure a. The formula is 2n = 4x = 36 = 2M + 14m + 18sm + 2st (Figure b). The total chromosome length varied from 6.4 μm to 3 μm. The karyotype therefore was a little asymmetric, because of a large number of subtelocentric and telocentric chromosomes. Chromosomes were generally of large size and showed a low degree of polymorphism in their shapes. All karyological parameters of H. dentatum subsp. xanthostylophorum (labeled as DX) are shown in Table . They have been compared with some available karyological parameters from the literature, belonging to taxa of the large agamic complex H. villosum/H. morisianum (sect. Villosa) (Pignatti Citation1982) or other related entities of Hieracium subgen. Hieracium. The Table dataset allowed us to make the scatter plots of Figure . The first scatter plot (Figure a), between A1 and A2 indices (Romero Zarco Citation1986), points out that the most symmetric karyotypes are those of taxa living in northern Europe, the Carpathian Region and Turkey (on the left side of the plot). On the other hand, the Italian Hieracium subgen. Hieracium populations appear to be the most asymmetric ones (on the right side of the plot). The second scatter plot (Figure b), between CVCL and MCA indices (Paszko Citation2006, Peruzzi & Eroğlu Citation2013), which, according to Peruzzi & Eroğlu (Citation2013) are the most appropriate parameters to estimate respectively interchromosomal and intrachromosomal asymmetries, points out that H. dentatum subsp. xanthostylophorum lies to the right end of the plot, quite far from Polish H. villosum population (VP), that should be the most similar, from a morphological and taxonomical point of view. This could be due to the different evolutionary processes that the two populations, inhabiting distant lands, have undergone over time.
Table 1. Karyological parameters of some taxa belonging to Hieracium subgen. Hieracium.
Figure 6 Scatter plots comparing the karyological parameters of some taxa belonging to Hieracium subgen. Hieracium, obtained (a) between A1 and A2 indices, and (b) between CVCL and MCA indices. For the meaning of the labels and index references, see Table .
Discussion and conclusion
Hieracium s. str. comprises an immense agamic complex with a base-number of x = 9. Triploids and tetraploids form the bulk of taxa investigated so far. Diploids (2n = 18) are rather rare and supposed to be confined to restricted geographical areas (Chrtek et al. Citation2004), reported mostly from SW Europe (e.g. in southern Tuscany, see Selvi and Fiorini Citation1996). In central and southern European mountains, H. villosum proved to be both an apomictic triploid (2n = 27) and an apomictic tetraploid (2n = 36) (Schuhwerk Citation1996; Severa and Chrtek Citation2002; Chrtek et al. Citation2004; Szeląg and Vladimirov Citation2005; Kruk Citation2011). In the same territories, H. dentatum was found to be an apomictic tetraploid (2n = 36) with, surprisingly, the same genotype as detected in H. villosum (Severa and Chrtek Citation2002). In the Balkan Peninsula (Niketić et al. Citation2006), in Austria and France (Tyler and Jönsson Citation2009), taxa considered of the H. dentatum group were found to be triploid or tetraploid. Our analysis seems to point out similar trends in the northern Apennine study area. The core taxon (H. villosum) is surrounded by tetraploid apomictic lineages that show some morphological characters of other Hieracium species, in the case of H. dentatum subsp. xanthostylophorum (= H. villosum “var. zangherii”), characters of H. bifidum.
This biosystematic interpretation well agrees with karyological analysis made on H. dentatum subsp. xanthostylophorum, which in the northern Apennines confirms the presence of tetraploid populations having an intermediate karyotype asymmetry between the populations of the Balkan Peninsula and those of southern Tuscany.
Even if this supposed locally endemic Hieracium variety cannot be confirmed, at least from a nomenclatural and morphological point of view, the presence in this small area of a high number of Hieracium taxa corroborate the highly relevant role in plant diversity and conservation (Viciani et al. Citation2010) of this territory, that for its geographical position can be considered a “trait d’union” between two very different phytogeographical typologies, a northern European one and a southern Mediterranean one.
Acknowledgements
We would like to thank Lorenzo Peruzzi and two other anonymous reviewers for their comments and suggestions that helped to improve this work.
Related Research Data
References
- Arano , H and Saito , H . 1980 . Cytological studies in family Umbelliferae 5. Karyotypes of seven species in subtribe Seselinae . Kromosomo , 2 : 471 – 480 .
- Bedini G, Garbari F, Peruzzi L, editors. 2010 onwards. Chrobase.it – Chromosome numbers for the Italian flora [online database]; cited 2012 Oct 10] Available from: http://www.biologia.unipi.it/chrobase/
- Bedini , G , Garbari , F and Peruzzi , L . 2012a . Karyological knowledge of the Italian vascular flora as inferred by the analysis of “Chrobase.it” . Plant Biosyst , 146 ( 4 ) : 889 – 899 .
- Bedini , G , Garbari , F and Peruzzi , L . 2012b . Chromosome number variation of the Italian endemic vascular flora. State-of-the-art, gaps in knowledge and evidence for an exponential relationship among even ploidy levels . Comp Cytogenet , 6 ( 2 ) : 192 – 211 .
- Coşkuncelebi , K and Hayirlioğlu-Ayaz , S . 2006 . Notes on chromosome numbers and karyotypes of five species in Hieracium L. s.str. (Asteraceae) from Turkey . Caryologia , 59 ( 1 ) : 19 – 24 .
- Chrtek , J Jr and Mráz P , Severa M . 2004 . Chromosome numbers in selected species of Hieracium s. str. (Hieracium subgen. Hieracium) in the Western Carpathians . Preslia , 76 ( 2 ) : 119 – 139 .
- Dyer , AF . 1979 . Investigating chromosomes , London : Edward Arnold Publishers .
- Gottschlich , G . 2005 . Hieracium grovesianum Arv.-Touv. ex Belli (Compositae), endemica italiana poco conosciuta. Tipificazione, note tassonomiche, ecologiche e corologiche. (Revisione delle collezioni di Hieracium Italiani, IV.) . Webbia , 59 ( 2 ) : 269 – 283 .
- Gottschlich , G . 2009a . Die Gattung Hieracium L. (Compositae) in der Region Abruzzen (Italien). Eine floristisch-taxonomische Studie . Stapfia , 89 : 1 – 328 .
- Gottschlich , G . 2009b . New species of the genus Hieracium L. (Compositae) from the Northern Apennine (Italy) . Webbia , 64 ( 2 ) : 175 – 186 .
- Gottschlich , G . 2011 . New taxa of Hieracium L. and Pilosella Vaill. (Compositae) from the Central Apennine (region of Marche incl. Umbrian and Latio parts of Monti Sibillini, Italy). (Revisions of Italian Hieracium collections VII.) . Webbia , 66 ( 2 ) : 195 – 230 .
- Guerra , M . 2012 . New trends in plant cytogenetics and cytoembryology – Dedicated to the memory of Emilio Battaglia . Plant Biosystems , 146 ( 3 ) : 703 – 710 .
- Huziwara , Y . 1962 . Karyotype analysis in some genera of Compositae. VIII. Further studies on the chromosome of Aster . Am J Bot , 49 : 116 – 119 .
- Ilnicki , T , Hasterok , R and Szeląg , Z . 2010 . Cytogenetic analysis of Hieracium transylvanicum (Asteraceae) . Caryologia , 63 ( 2 ) : 192 – 196 .
- Johansen , DA . 1940 . Plant Microtecnique , New York : McGraw-Hill . p. 40
- Kiran , Y , Turkoglu , I , Kirilmaz , F , Arabaci , T , Sahin , A and Bagci , E . 2012 . Karyological investigation of six Achillea L. (Asteraceae) species growing in Turkey . Caryologia , 65 ( 2 ) : 101 – 105 .
- Kruk , J . 2011 . A new stand of Hieracium pilosum Schleich. ex Froelich (Asteraceae) in the Polish Tatra Mountains . Acta Soc Bot Polon , 80 ( 1 ) : 33 – 35 .
- Levan , A , Fredga , K and Sandberg , AA . 1964 . Nomenclature for centromeric position on chromosomes . Hereditas , 52 : 201 – 220 .
- McNeill, J, Barrie, FR, Buck, WR, Demoulin, V, Greuter, W, Hawksworth, DL, Herendeen, PS, Knapp, S, Marhold, K, Prado, J, Prud’homme van Reine, WF, Smith, GF, Wiersema, JH & Turland, NJ. (eds. & comps.): International Code of Nomenclature for algae, fungi, and plants (Melbourne Code), adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011. Regnum Vegetabile, 154. Koeltz Scientific Books, Königstein (ISBN 978-3-87429-425-6).
- Meng , Y , Nie , Z-L , Sun , H and Yang , Y-P . 2012 . Chromosome numbers and polyploidy in Leontopodium (Asteraceae: Gnaphalieae) from the Qinghai-Tibet Plateau of S.W. China . Caryologia , 65 ( 2 ) : 87 – 93 .
- Niketić , M , Vladimirov , V and Mràz , P . 2006 . Chromosome numbers and taxonomic-chorological notes on selected species of Hieracium s. str. (Asteraceae) from Montenegro . Phytol Balcan , 12 ( 1 ) : 85 – 97 .
- Ozcan , M , Hayirlioğlu-Ayaz , S and Inceer , H . 2011 . Chromosome reports in some Cirsium (Asteraceae, Cardueae) taxa from north-east Anatolia . Caryologia , 64 ( 1 ) : 55 – 66 .
- Paszko , A . 2006 . A critical review and a new proposal of karyotype asymmetry indices . Plant Syst Evol , 258 : 39 – 48 .
- Peruzzi , L and Eroğlu , HE . 2013 . Karyotype asymmetry: again, how to measure and what to measure? . Comp Cytogenet , 7 ( 1 ) : 1 – 9 .
- Pignatti S. 1982. Flora d’Italia. 1–3. Bologna: Edagricole.
- Romero , Zarco C . 1986 . A new method for estimating karyotype asymmetry . Taxon , 35 ( 3 ) : 526 – 530 .
- Schuhwerk F. 1996. Published chromosome-counts in Hieracium; [cited 2012 Oct 10]. Available from: http://www.botanischestaatssammlung.de/projects/chrzlit.html
- Selvi , F and Fiorini , G . 1996 . Karyology of Hieracium L. subg. Hieracium (Asteraceae) from Mount Amiata (Central Italy) . Caryologia , 49 : 287 – 299 .
- Severa M, Chrtek J Jr. 2002. On Hieracium villosum and some related taxa in the Western Carpathians. 6th Hieracium Workshop; 2002 Jul 17–24; Hirschegg (Austria). Contribution Abstract volume. p. 22.
- Szeląg , Z and Vladimirov , V . 2005 . Chromosome numbers of Polish Hieracia (Asteraceae) . Polish Bot J , 50 ( 2 ) : 139 – 143 .
- Tyler , T and Jönsson , J . 2009 . Ploidy level analysis of apomictic Hieracium (Asteraceae) reveal unexpected patterns and variation . Nord J Bot , 27 : 490 – 502 .
- Viciani , D and Agostini , N . 2009 . La carta della vegetazione del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (Appennino Tosco-Romagnolo): note illustrative . Quad Studi Nat Romagna , 27 ( 2008 ) : 97 – 134 .
- Viciani , D , Gonnelli , V , Sirotti , M and Agostini , N . 2010 . An annotated check-list of the vascular flora of the “Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna” (Northern Apennines Central Italy) . Webbia , 65 ( 1 ) : 3 – 131 .
- Watanabe , K , Yahara , T , Denda , T and Kosuge , K . 1999 . Chromosomal evolution in the genus Brachyscome (Asteraceae, Astereae): Statistical tests regarding correlation between changes in karyotype and habit using phylogenetic information . J Plant Res , 112 : 145 – 161 .
- Zangheri P. 1966. Repertorio Sistematico e Topografico della Flora e Fauna Vivente e Fossile della Romagna. Tomo 1. Verona: Memorie del Museo Civico di Storia Naturale di Verona., Mem. Fuori serie n. 1.