Abstract
Chromosome number and morphology in nine taxa belonging to 10 populations of Tanacetum L. (Asteraceae) were investigated using karyological techniques. The chromosome number found in eight species was 2n = 18 (T. balsamita, T. budjnurdense, T. sonbolii, T. archibaldii, T. persicum, T. parthenium and T. canescens) while T. polycephalum subsp. duderanum and T. polycephalum subsp. azerbaidjanicum had 2n = 36. Six records are new while four records confirmed the previous data. At mitotic metaphase, all chromosomes had median point (M), median region (m), submedian (sm) or subterminal (st) centromeres. The karyotypes are classified in types 2A or 2B, showing the presence of a primitive symmetrical karyotype in the genus Tanacetum. An updated checklist of karyological data of the genus is also presented.
Introduction
Asteraceae, the largest family of flowering plants, comprises 1600–1700 genera and around 23,000 species distributed worldwide except for Antarctica (Funk et al. Citation2009). The Anthemideae is a medium sized tribe in the family (Vallès et al. Citation2005) comprising 111 genera with c.1800 species (Oberprieler, Himmelreich, et al. Citation2007). Tanacetum with around 160 species is the third largest genus of Anthemideae, distributed in Europe, Asia, northern Africa and North America (Oberprieler, Vogt, et al. Citation2007). This genus is represented by 54 species in the Flora Iranica area (Podlech Citation1986), and in Iran by 35 species according to the most recent findings (Mozaffarian Citation2005, Citation2008; Djavadi Citation2008; Sonboli et al. Citation2010). The most common base chromosome number of the Anthemideae is x = 9 and this is possibly the basic number for the family as a whole (Semple and Watanabe Citation2009). Variation in chromosome number has been brought about mainly by the processes of polyploidy (including both allo- and autopolyploidy), aneuploid change and chromosome loss (Kadereit and Jeffrey Citation2007). Chromosome counts in many members of Anthemideae have already been reported (e.g. Torrell et al. Citation2001; Inceer and Beyazoglu Citation2004; Vallès et al. Citation2005; Garcia et al. Citation2006; Inceer et al. 2007, Citation2012; Li et al. Citation2008; Chehregani and Mehanfar Citation2008; Chehregani and Hajisadeghian Citation2009; Chehregani et al. Citation2011).
A literature survey concerning the available data of chromosome numbers in the Asteraceae (Watanabe Citation2013), has shown the existence of previous cytogenetic records for some Tanacetum species (see Appendix). Karyological studies of the genus Tanacetum in Iran have been performed only by a few researchers (Chehregani and Mehanfar Citation2008; Chehregani and Hajisadeghian Citation2009; Chehregani et al. Citation2011) and the reported chromosome numbers include 2n = 18, 27, 36, 54, 72 and 90. Chromosomal data are still scarce or not existent for numerous Tanacetum taxa. The aim of the present study is to elucidate the chromosome numbers and karyological characteristics of 10 representatives of Tanacetum from different localities of Iran.
Material and methods
Ten Tanacetum taxa including eight species belonging to four sections of the genus were used in this study (Table ). The ripe achenes of the taxa studied were collected from the north, north-east and north-west of Iran, in 2008–2011. Voucher specimens of the studied taxa are deposited in the Medicinal Plants and Drug Research Institute Herbarium (MPH), Shahid Beheshti University of Tehran.
Table 1. Somatic chromosome number, ploidy level, karyotype formula, ranges of chromosome length, total karyotype length (TKL), asymmetry indices (A1, A2) of Romero Zarco (1986), symmetry classes (SC) of Stebbins (Citation1971) and total form percentage (TF%) for Tanacetum taxa.
Mature achenes were collected from natural populations and germinated at 24°C on moist filter paper in Petri dishes. Actively growing root tips, 1 cm in length, were excised from germinating seed and pretreated with 0.002 M 8-hydroxyquinoline for 3–3.5 h at room temperature. The material was fixed with Carnoy (1:3 glacial acetic acid/absolute ethanol) for 24 h at 4°C and stored in 70% ethanol at 4°C until use. They were hydrolysed in 1 N HCl for 10 min at 60°C, rinsed in tap water for a minimum of 3–4 min and then meristematic regions were stained and squashed on slides with 2% aqueous aceto-orcein for three minutes. The best metaphase plates of each specimen were photographed with Olympus BX-51 microscope.
Karyotypes were obtained from 3–5 metaphase plates per population. Long arm, short arm and total length of each chromosome were measured and relative length (RL), arm ratio (AR) and centromeric index (CI) were determined. The chromosome nomenclature followed Levan et al. (Citation1964). The intra-chromosomal asymmetry index (A1) and the inter-chromosomal asymmetry index (A2) were calculated according to the formula proposed by Romero Zarco (1986). The karyotype symmetry nomenclature followed Stebbins (Citation1971). In the haploid idiograms, chromosomes were arranged in decreasing order according to their length.
Results and discussion
The infrageneric classification of Tanacetum is an unresolved problem. In the present study, we have followed the nomenclature in the Flora of Iranica adopted by Podlech (Citation1986). The somatic metaphase chromosomes of the nine Tanacetum taxa belonging to 10 populations studied and their karyomorphological details are presented in Tables – and Figures –. We provide now the information on each taxon.
Table 3. Karyomorphological parameters of the tetraploid Tanacetum taxa. Chromosome pairs are in Roman numerals.
Table 2. Karyomorphological parameters of the diploid Tanacetum taxa. Chromosome pairs are in Roman numerals.
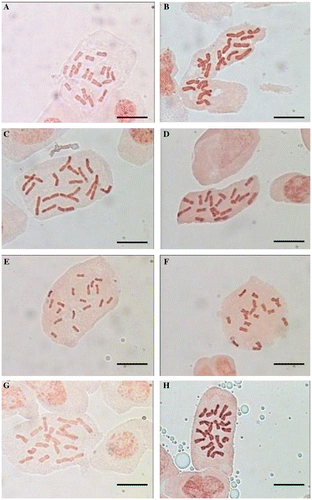
Figure 1. (Color online) Metaphase chromosomes of Tanacetum studied taxa. (A) T. archibaldii; (B) T. balsamita subsp. balsamitoides; (C) T. budjnurdense; (D) T. persicum; (E) T. parthenium (Hamedan); (F) T. parthenium (Tehran); (G) T. canescens; (H) T. sonbolii. Scale bars, 10 μm.
Tanacetum sect. Xylopyrethrum
T. archibaldii Podlech, 2n = 18
Iran: Mazandaran, Chalus road, Pol-e Zanguleh, 3400 m, 17 August 2011, Sonboli and Mehregan, 1790 (MPH).
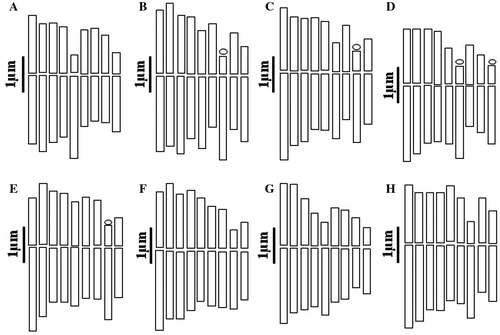
According to our data, this is the first report of the chromosome number and karyomorphology for this species, which is endemic and rare to Iran. It is a diploid based on the most common basic chromosome number in the genus, x = 9 (Inceer et al. Citation2012). The ratio of the longest to the shortest chromosome is 1.6 and the karyotype symmetry is 2A. The karyotype consists of six median region (m) chromosome pairs (1, 2, 3, 4, 6 and 8), one median point (M) (7), one submedian (sm) (9) and one subterminal (st) (5). None of the chromosomes carries an obvious satellite (Table and Figures A, A). The relative length of the chromosomes varies from 8.60 to 13.61 (Table ).
Figure 2. (Color online) Haploid idiograms of the studied taxa from Tanacetum. (A) T. archibaldii; (B) T. balsamita subsp. balsamitoides; (C) T. budjnurdense; (D) T. persicum; (E) T. parthenium (Hamedan); (F) T. parthenium (Tehran); (G) T. canescens; (H) T. sonbolii.
Tanacetum sect. Balsamita
T. balsamita L. subsp. balsamitoides (Sch.-Bip.) Grierson, 2n = 18
Iran: Mazandaran, Chalus road, Pol-e Zanguleh, 3165 m, 17 August 2011, Sonboli and Mehregan, 1788 (MPH).
To our knowledge, this is the second report on chromosome number for this subspecies and the first in an Iranian population. Our study confirms the diploid cytotype of this species which has already been reported by Khandjian (Citation1975). The ratio of the longest to the shortest chromosome is 1.5 and the karyotype symmetry is 2A. The karyotype consists of five median region (m) chromosomes (1, 3, 4, 5 and 8), two median point (M) (2 and 6), one submedian (sm) (9) and one subterminal (st) (7). One satellite was observed on the short arm of subtelocentric chromosome (Table and Figures B, B). The relative chromosome length varies from 8.94 to 13.45 (Table ).
Tanacetum sect. Tanacetum
T. budjnurdense (Rech.f.) Tzvelev, 2n = 1
Iran: Khorasan, Bujnourd, Reein towards Garmak, Zoo valley, 1700 m, 20 June 2006, Zangooei and Arjmandi, 1477 (MPH).
According to our literature review, this is first count on the chromosome number and morphology of this species, an endemic and range-restricted species to north-eastern of Iran. Its chromosome number is 2n = 18 (based on x = 9). The ratio of the longest to the shortest chromosome is 1.8:1 and the karyotype symmetry is 2A. The karyotype consists of six median region (m) chromosome pairs (1, 2, 3, 4, 5 and 9), one median point (M) (7), one submedian (sm) (6) and one subterminal (st) (8). Chromosome pair 8 has satellites connected to the short arms (Table and Figures C, C). The relative length of chromosomes varies from 8.42 to 15.26 (Table ).
Tanacetum sect. Parthenium
T. persicum (Boiss.) Mozaff. 2n = 18
Iran: Chahar Mahal and Bakhtiari, Boroujen, Sabz Kuh protected area, 2800–3000 m, 16 July 2006, Sonboli, Kanani and Gholipour, 1502 (MPH).
This is the first count on the chromosome number and morphology for this species, endemic to central and southern parts of Iran and Iraq. It again shows the existence of the diploid cytotype based on x = 9. The ratio of the longest to the shortest chromosome is 1.7 and the karyotype symmetry is 2A. The karyotype consists of five median region (m) chromosome pairs (1, 2, 4, 5 and 8), two median point (M) (3 and7), one submedian (sm) (9) and one subterminal (st) (6). Chromosomes 6 and 9 have satellites on short arms (Table and Figures D, D). The relative chromosome length varies from 8.40 to 14.46 (Table ).
T. parthenium Sch. Bip. 2n = 18
Iran: Hamadan, Morad Beig valley, 1850 m, 8 July 2011, Olanj, 1903 (MPH).
Iran: Tehran, Velenjak, Tuchal Mountain, 1800 m, 18 July 2011, Olanj, 1483 (MPH).
The chromosome number of this widely distributed and medicinal species has been counted by several authors. Our count confirms some previous reports as diploid level based on x = 9 (Watanabe Citation2013 and references cited therein; Inceer and Hayirlioglu-Ayas Citation2007; Chehregani and Hajisadeghian Citation2009; Inceer at al. Citation2012), although we found three previous records of the tetraploid level in Germany, Russia and Iran (Abd El-Twab and Kondo Citation2001; Zhmyleva and Kondo Citation2006; Chehregani and Mehanfar Citation2008). Two populations of this species were diploid (2n = 2x = 18) with some minor karyomorphological differences (Table ).
In the Hamedan population, the metaphase complement consists of three median point (M) chromosomes (3, 4 and 6), three median region (m) (2, 5 and 7), two submetacentric (sm) (1 and 9) and one subtelocentric (st) (8). Chromosome pair 8 has satellites connected to the short arms. The ratio of the longest to the shortest chromosome is 1.7 and the karyotype symmetry is 2A (Table and Figures E, E). The relative chromosome length varies from 8.31 to 13.87 (Table ). In the Tehran population, karyotype showed a higher degree of symmetry with three median point (M) chromosomes (2, 4 and 5), four median region (m) (1, 3, 6 and 7) and two submedian (sm) (8 and 9). The arm ratio was between 1 and 1.7, the relative chromosome length range is from 8.17 to 14.08 and the karyotype symmetry is 2A (Tables , and Figures F, F).
Tanacetum sect. Xanthoglossa
T. canescens DC. 2n = 18
Iran: Zanjan, Soltaniyeh towards Ghidar, km 9, 2024 m, 20 July 2011, Olanj, 1912 (MPH).
This count of chromosome number and morphology is the first for T. canescens. It is a diploid based on x = 9. The karyotype shows a relatively high degree of symmetry (2B), consisting of six metacentric (1, 2, 3, 6, 7 and 8) and three submetacentric (4, 5 and 9) chromosomes. No satellite was observed in the karyotype of this species. Arm ratio is 2.1 and the relative chromosome length varies from 7.43 to 15.60 (Tables , and Figures G, G).
T. sonbolii Mozaff. 2n = 18
Iran: West Azerbaijan, Takab, Baderlu village, 2400 m, 13 July 2003, Sonboli, 305 (MPH).
According to our information this is the first report of the chromosome number and morphology of T. sonbolii, an endemic and recently described species from Iran. The species is a diploid based on x = 9. The karyotype shows a high degree of symmetry, as all the chromosomes pairs have median region centromeres except for one pair (chromosome 7) which has subterminal centromeres; there is no microsatellite. The karyotype symmetry is 2A, with arm ratio between 1 and 1.6 and the relative length of chromosome varies from 8.98 to 14.02 (Tables , and Figures H, H).
Tanacetum polycephalum Sch.Bip. subsp. azerbaidjanicum Podlech, 2n = 36
Iran: West Azerbaijan, Urmia, Urmia towards Salmas, Ghushchi pass, 1860 m, 2 July 2007, Sonboli, 1212 (MPH).
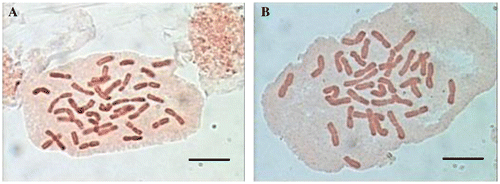
The chromosome number of this taxon has not been reported before and this is also the first report on the chromosome number and morphology of this species, which is endemic to Iran and Iraq. It is a tetraploid based on x = 9. Twelve pairs (1, 3, 4, 5, 6, 7, 9, 10, 11, 13, 16 and 17) have median region centromeres, four pairs (2, 8, 12 and 15) submetacentric, and two pairs of subtelocentric ones (14 and 18). Regarding the karyotypic symmetry, this species belongs to the 2B class of Stebbins. The chromosome length ranges from 3.64 to 8.50 and the arm ratio is 2.33 (Tables , and Figures A, A).
Figure 4. Haploid idiograms of the studied taxa from Tanacetum. (A) T. polycephalum subsp. azerbaidjanicum; (B) T. polycephalum subsp. duderanum.
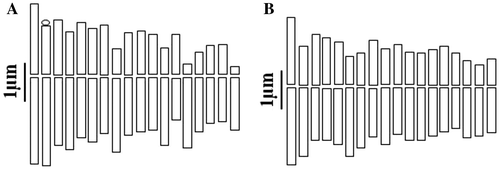
Figure 3. Metaphase chromosomes of the studied taxa from Tanacetum. (A) T. polycephalum subsp. azerbaidjanicum; (B) T. polycephalum subsp. duderanum. Scale bars, 10μm.
T. polycephalum Sch.Bip. subsp. duderanum (Bioss.) Podlech, 2n = 36
Iran: Mazandaran, Chalus road, Pol-e Zanguleh, 3400 m, 17 August 2011, Sonboli and Mehregan, 1795 (MPH).
This is the second count for this species, an endemic to Iran. Our count agrees with the previous one (Ghaffari and Kelich Citation2006) in which the gametic chromosome number of this species was reported as n = 18. Our report is therefore the first one based on somatic cells. The karyotype shows a high degree of symmetry (2B), 14 pairs (1, 2, 4, 5, 7, 8, 9, 10, 11, 12, 13, 14, 15 and 18) have median region centromeres, three submedian pairs are 6, 16 and 17 and the single median point pair is 3. None of the chromosomes carried an evident satellite. The arm ratio was between 1 and 1.4 and the relative chromosome length varies from 4.26 to 8.52 (Tables , and Figures B, B).
Discussion
This research contributes karyological data in a relevant set of Tanacetum taxa, sharing the opinion that chromosome number is now a crucial parameter in plant systematics and evolution (Stuessy and Lack Citation2011, and references therein). Out of the nine taxa examined, four are endemic to Iran (T. budjnurdense, T. sonbolii, T. archibaldii and T. polycephalum subsp. duderanum) and two endemic to Iran and Iraq (T. persicum and T. polycephalum subsp. azerbaidjanicum).
In this study the karyology of six out of nine taxa with unknown karyotype characters to date were presented for the first time (T. budjnurdense, T. sonbolii, T. archibaldii, T. polycephalum subsp. azerbaidjanicum, T. persicum and T. canescens). The other counts (T. parthenium, T. polycephalum subsp. duderanum and T. balsamita subsp. balsamitoides) confirmed previous reports.
All the studied taxa had the basic chromosome number of x = 9 (Table ), predominant in tribe Anthemideae and the family Asteraceae (Inceer and Beyazoglu Citation2004 and references cited therein; Valles et al. 2005; Inceer et al. Citation2012). Seven taxa were diploid with 2n = 18, and two taxa were tetraploid with 2n = 4x = 36. In all the studied taxa, the most frequent chromosome type was found to be m, followed by sm and M. The st (subterminal chromosome) was observed in six taxa, i.e. T. balsamita subsp. balsamitoides, T. budjnurdense, T. sonbolii, T. archibaldii, T. persicum and T. parthenium (Table ).
The asymmetry of the karyotype according to the Stebbins (Citation1971) system classifies the studied taxa into two groups, namely 2A (70%) and 2B (30%). Species from the 2A group show the lowest proportion of chromosomes with an arm ratio > 2.00 and include T. balsamita subsp. balsamitoides, T. budjnurdense, T. sonbolii, T. archibaldii, T. persicum and T. parthenium. Group 2B includes T. polycephalum subsp. duderanum, T. polycephalum subsp. azerbaidjanicum and T. canescens with a slightly more asymmetrical karyotype than group 2A. In a scattergram (Figure ) on the basis of the asymmetry indices (A1 and A2) of Romero Zarco (1986), two major groups are formed which clearly correspond to the Stebbins classes 2A and 2B. According to Stebbins (Citation1971) this is an indicative of derived karyotypes, which is in agreement with the polyploid character of these populations (derived in respect to diploid ones) and, in general, of the polyploidy as an evolutionary mechanism. Six species of Achillea L. in Turkey were investigated, all chromosomes had 2n = 18 and increase in asymmetry was not observed in the karyotypes of the species studied (Kiran et al. Citation2012).
Figure 5. Scattergram of intrachromosomal (A1) and interchromosal (A2) asymmetry indices of all taxa of Tanacetum investigated. The taxa are numbered as in Table .
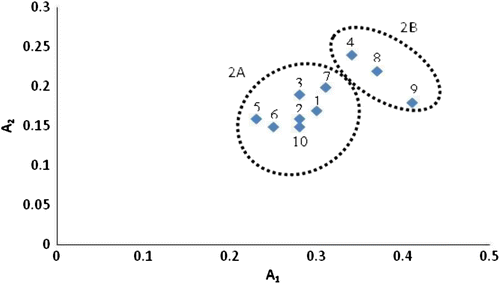
The intra chromosomal asymmetry index (A1) ranges from 0.23 in T. parthenium (Hamedan) to 0.37 in T. polycephalum subsp. azerbaidjanicum. Inter chromosomal asymmetry index (A2) ranges from 0.15 in T. sonbolii and T. parthenium (Tehran) to 0.24 in T. canescens (Table ). The asymmetry index based on A1 values revealed the highest asymmetry of the karyotype for T. polycephalum subsp. azerbaidjanicum and this species is considered as more derived in relation to chromosomal evolutionary processes. Two studied populations of T. parthenium (Hamedan and Tehran) show the shortest mean chromosome length (1.81 and 1.63 μm) among the studied taxa, while the longest chromosome pair was observed in T. polycephalum subsp. duderanum (7.85 μm), and the longest total haploid chromosome length (91.6 μm) of all the studied species. The shortest total haploid chromosome length (20 μm) was observed in the Tehran population of T. parthenium. The karyotype length is positively correlated with genome size (Garnatje et al. Citation2004; Garcia et al. Citation2005), so it could be concluded that T. polycephalum subsp. duderanum and T. parthenium (Tehran population) would possess highest and lowest genome size among the studied taxa, respectively.
Several ploidy levels (2x, 4x, 6x, 8x, 10x) have already been reported within the Tanacetum polycephalum complex group (Khandjian Citation1975, Citation1992; Ghaffari and Kelich Citation2006; Chehregani and Mehanfar Citation2008; Chehregani and Hajisadeghian Citation2009; Chehregani et al. Citation2011). Our results for the presence of tetraploidy in two subspecies of T. polycephalum, i.e. T. polycephalum subsp. duderanum and T. polycephalum subsp. azerbaidjanicum, confirm the role of this evolutionary process as one of the possible diversification forces in this genus. According to Oberprieler et al. (Citation2009), the reconstruction of chromosome number evolution has shown that x = 10 is the most reasonable base chromosome number for the common ancestor of the tribe Anthemideae. A transition to x = 9, however, is observed very close to the base of the tree and this base chromosome number is reconstructed for some genera such as Tanacetum, Tripleurospermum, Achillea and Anthemis.
Current status of chromosome number knowledge in Tanacetum
No cases of dysploidy, frequent in other genera of the tribe Anthemideae such as Artemisia, have been reported in Tanacetum, which has been useful in establishing systematic and taxonomic position of some taxa (Sonboli et al. Citation2011). Of the listed taxa (see Appendix), 29 show 2x complement (78.4%) of which 21 are exclusively diploid (2n = 18); three taxa show 3x complement (8.1%) (2n = 27); eight taxa show 4x complement (21.6%) of which three are exclusively tetraploid (2n = 36); five taxa show 6x complement (13.5%) of which two are exclusively hexaploid (2n = 54); one taxon (2.7%) is 8x (2n = 72); one taxon (2.7%) is 10x (2n = 90); and 11 taxa show aneuploidy or haploidy (29.7%) of which only one is exclusively aneuploid (2n = 14). Of the considered taxa, 58.33% are diploid, 2.77% triploid, 8.33% tetraploid, 5.55% hexaploid, 2.77% octaploid, 2.77% decaploid, and 2.77% aneuploidy; 13.97% show two or more cytotypes (representing more or less completely euploid series) or the very rare possibility of aneuploidy/haploidy phenomena (2.74%). The highest chromosome number occurs in T. polycephalum Sch. Bip. (2n = 90), while the lowest (excluding a probable case of aneuploidy), 2n = 12, is in T. cinerariifolium Sch. Bip. The percentages of the genus Tanacetum now covered by information on chromosome counts in general and Iran are 20% and 23% respectively. Further investigation is needed regarding Tanacetum karyology, cytogenetics, systematic and evolution.
Appendix. List of chromosome numbers of Tanacetum taxa retrieved from published data.
Acknowledgements
Financial support is acknowledged from the Research and Technology Council of Shahid Beheshti University. Appreciation is expressed to three anonymous referees with helpful comments.
The authors are thankful to Prof. Dr. Joan Vallès and Dr. Sonia Garcia from Barcelona University for their kind comments.
References
- Abd El-Twab MH , Kondo K . 2001. Genome territories of Dentranthema horaimontana in mitotic nuclei of F1 hybrid between D. horaimontana and Tanacetum parthenium . Chromosome Sci. 5(2):63–71.
- Abd El-Twab MH , Kondo K . 2004. Rapid genome changes in diploid F1 hybrids investigated by GISH and physical mapping of 5S and 45S rDNA by FISH. Chromosome Sci. 8(4):198.
- Arohonka T . 1982. Chromosome counts of vascular plants of the island Seili in Nauvo, southwestern Finland. Turun Yliopiston Julkaisuja: Sarja A II, Biologia-Geographica. 3(1):1–12.
- Baltisberger M . 1993. Zytologische Untersuchungen an Compositen aus Albanien. Candollea. 48(2):437–448.
- Baltisberger M . 2006. Cytological investigations on Bulgarian phanerogams. Willdenowia. 36(1):205–216.
- Brown AMG , Edwards CM , Hartman TPV , Marshall JA , Smith RM , Daver MR , Power JB , Lowe KC . 1999. Sexual hybrids of Tanacetum: biochemical, cytological and pharmacological characterization. J. Exp. Bot. 50 (333): 435–444.
- Chehregani A , Atri M , Sarmadi J , Asgari M . 2011. Chromosome number variation in Tanacetum polycephalum Schultz Bip. (L.) (Asteraceae) in west of Iran. Caryologia. 64(3):302–308.
- Chehregani A , Hajisadeghian S . 2009. New chromosome counts in some species of Asteraceae from Iran. Nord J Bot. 27(3):247–250.
- Chehregani A , Mehanfar N . 2008. New chromosome counts in the tribe Anthemideae (Asteraceae) from Iran. Cytologia. 73(2):189–196.
- Chinnappa CC , Chmielewski JG . 1987. Documented plant chromosome numbers 1987: 1. Miscellaneous counts from western North America. Sida. 12(1):409–417.
- Corsi G . 1962. Contributo alla cariologia di Chrysanthemum cinerariaefolium Vis. Caryologia. 15(1):123–129.
- Davlianidze MT . 1985. Chromosome numbers in the representatives of the flora from Georgia. Bot Zh Bot. Zhurn. 70 (5):698–700.
- Dawe, JC , Murray DF . 1981. Atlas of chromosome numbers documented for the Alaskan flora. University of Alaska Museum Herbarium USA. p. 1–3.
- Dempsey RE , Gornall RJ , Bailey JP . 1994. Contributions to a cytological catalogue of the British and Irish flora. Watsonia. 20(1):63–66.
- Djavadi SB . 2008. Three new records of Tanacetum for the flora of Iran. Rostaniha. 9(1):23–32.
- Dmitrieva SA , Parfenov VI . 1983. Khromosomnye chisla nekotorykh vidov flory Belorussii. Izvestiia Akademii Nauk Belorusskoi SSR: Ser. Biol. Nauk. 2:7–12.
- Dowrick GJ . 1952. The chromosomes of Chrysanthemum . I. The species. Heredity. 6(3):365–375.
- Febles R , Fernández-Peratla AM , González-Aguilera JJ . 1989. Análisis cariotipico de las especies endémicas del género Tanacetum L. (Anthemidae-Asteraceae). Bot Macaron. 18(8):59–68.
- Ferakova V . 1978. Index of chromosome numbers of Slovakian flora. Part 6. Acta Fac. Rerum Nat. Univ. Comenianae Bot. Bratislava. 26:1–42.
- Ferrarella A , Grisafi F , Lentini F , Melati MR . 1981. In: Numeri cromosomici per la Flora Italiana: 860–867. Inform Bot Ital. 13:189–194.
- Frey L , Mizianty M , Mirek Z . 1981. Chromosome numbers of Polish vascular plants. Fragm. Florist. Geobot. 27(4):581–590.
- Funk VA , Susanna A , Stuessy TF , Bayer RJ . 2009. Systematics, evolution and biogeography of the Compositae. Vienna: International Association of Plant Taxonomy.
- Gagnidze RI , Gviniashvili T . 1997. IOPB chromosome data. Newsl Int Organ Plant Biosyst. 11(26/27):20–21.
- Garcia S , Garnatje T , Dariimaa SH , Tsooj S , Vallès J . 2006. New or rarely reported chromosome numbers in taxa of subtribe Artemisiinae (Anthemideae, Asteraceae) from Mongolia. Bot J Linn Soc. 150(2):203–210.
- Garcia S , Inceer H , Garnatje T , Vallès J . 2005. Genome size variation in some representatives of the genus Tripleurospermum . Biol Plant. 49(3):381–387.
- Garnatje T , Vallès J , Garcia S , Hidalgo O , Sanz M , Canela MA , Siljak Yakovlev . 2004. Genome size in Echinops L. and related genera (Asteraceae, Cardueae): karyological, ecological and phylogenetic implications. Biol Cell. 96(2):117–124.
- Ghaffari SM , Kelich K . 2006. New or rare chromosome counts of some angiosperm species from Iran. Iran J Bot. 12(1):81–86.
- Gregor T , Hand R . 2007. Chromosomenzahlen von Farn- und Samenpflanzen aus Deutschland 2. Kochia. 2:31–35.
- Gvinianidze ZI , Avazneli AA . 1982. Khromosomnye chisla nekotorykh predstavitelej vysokogornykh floristichesken kompleksov Kavkaza. Soobsc Acad Nauk Gruzinsk SSR (Tbilisi). 106(3):577–580.
- Hollingsworth PM , Gornall RJ , Bailey JP . 1992. Contribution to a cytological catalogue of the British and Irish flora, 2. Watsonia. 19(1):134–137.
- Inceer H , Beyazoglu O . 2004. Karyological studies in Tripleurospermum (Asteraceae, Anthemideae) from north-east Anatolia. Bot J Linn Soc. 146(4):427–438.
- Inceer H , Hayirlioglu-Ayaz S . 2007. Chromosome numbers in the tribe Anthemideae (Asteraceae) from north-east Anatolia. Bot J Linn Soc. 153(2):203–211.
- Inceer H , Hayirlioglu-Ayaz S , Guler HS , Aksu N , Ozcan M. 2012. Karyological studies of some representatives of Tanacetum L. (Anthemideae-Asteraceae) from north-east Anatolia. Plant Syst Evol. 298(4):827–834.
- Kadereit JW , Jeffrey C. 2007. The families and genera of vascular plants. 8: Flowering plants, eudicots, Asterales. Berlin: Springer.
- Kamel EA . 2001. Chromosome counts and karyological studies on six taxa of the Egyptian Asteraceae. Comp. Newsl. 36:81–93.
- Khandjian NS . 1975. New data on chromosome number in Tanacetum L. and Leucanthemum Mill. Biol. Zhurn. Armenii. 28(1):87–89.
- Khandjian NS . 1992. The taxonomic significance of the achene’s structure in the subtribe Anthemideae (Asteraceae). Bot. Zhurn. 77:89–98.
- Kiehn M , Vitek E , Dobea C . 2000. Documented chromosome number checklist of Austrian vascular plants. Vienna: Verlag des Naturhistorischen Museums.
- Kiran Y , Turkoglu I , Kirilmaz F , Arabaci T , Sahin A , Bagic E . 2012. Karyological investigation of six Achillea L. (Asteraceae) species growing in Turkey. Caryologia. 65(1):101–105.
- Krasnikov AA , Lomonosova MN . 1990. Chromosome numbers in representatives of some families of vascular plants in the flora of the Novosibirsk region. I. Bot. Zhurn. 75(1):116–118.
- Krogulevich RE . 1978. Karyological analysis of the species of the flora of eastern Sayana Mts. In: Flora of the Prebaikal Region Novosibirsk, edited by Malyshev LI , GA Peshlcova , p. 19–48. Nauka.
- Krogulevich RE . 1984. Khromosomnye Chisla Tsvetkovykh Rastenii i Sibiri Dal’nego Vostoka. (Chromosome numbers of flowering plants of the Siberia and Far East). In: Izdatel’stvo “Nauka”, Sibirskoe Otdelenie. Krogulevich RE , Rostovtseva TS , Novosibirsk: Nauka. 286.
- Kuzmanov BA , Georgieva SB , Nikolova VA . 1986. Chromosome numbers of Bulgarian flowering plants. I. Fam. Asteraceae. Fitologija. 31:71–74.
- Kuzmanov BA , Nikolova V . 1980. In: Love A, editor. IOPB chromosome number reports. LXIV. Taxon. 29(5/6):714–715.
- Lavrenko AN , Serditov NP . 1987. Chromosome numbers in some members of the Urals flora (the Komi Autonomous Soviet Socialist Republic.). Bot. Zhurn. 72(6):846–847.
- Lavrenko AN , Serditov NP , Ulle ZG . 1991. Chromosome numbers in some species of vascular plants from the Pechoro-Ilychsky Reservation (Komi ASSR). Bot. Zhurn. 76:473–476.
- Lavrenko AN , Serditov NP , Ulle ZG . 1988. Chromosome number in some species of Asparagaceae, Asteraceae and Ranunculaceae from the north Urals (Komi ASSR). Bot. Zhurn. 73(2):605–607.
- Lavrenko AN , Serditov NP , and Ulle ZG. 1990. Chromosome numbers in some species of flowering plants of the Urals (the Komi Autonomous Soviet Socialist Republic). Bot. Zhurn. 75(11):1622–1624.
- Levan A , Fredga K , Sandberg AA . 1964. Nomenclature for centromeric position on chromosomes. Hereditas. 52(2):201–220.
- Li J , Chen SM , Chen FD , Fang WM . 2008. Karyotype and meiotic analyses of six species in the subtribe Chrysantheminae. Euphytica. 164(1):293–301.
- Lövkvist B , Hultgård UM . 1999. Chromosome numbers in south Swedish vascular plants. Opera Bot. 137:1–42.
- Martin-Noguet D . 1969. Contribution à l’etude cytotaxinomique des Achillea et des Phyrethum de Turquie. Bull. Soc. Bot. 116:197–206.
- Morton JK . 1977. A cytological study of the Compositae (excluding Hieracium and Taraxacum) of the British Isles. Watsonia. 11:211–223.
- Morton JK . 1981. Chromosome numbers in Compositae from Canada and the U. S. A. Bot J Linn Soc. 82(4):357–368.
- Mozaffarian M . 2005. Notes on the tribe Anthemideae (Compositae), new species, new records and new combinations for Iran. Iran J Bot. 11(1):115–127.
- Mozaffarian M. 2008. Flora of Iran, Compositae: Anthemideae and Echinopeae. In: Tanacetum L. edited by M. Assadi et al. 59:134–198. Tehran: Research Institute of Forests and Rangelands.
- Murín A . 1993. Karyologiche studium okrasnych rastlin flory Slovenska. Biologia. 48(1):441–445.
- Oberprieler C , Himmelreich S , Kallersjo M , Valles J , Watson LE , Vogt R . 2009. Anthemideae. In: Funk VA , Susanna A , Stuessy TF , Bayer RJ , editors. Systematics, evolution and biogeography of the Compositae. Vienna: International Association Plant Taxonomy, p. 631–666; 965p.
- Oberprieler C , Himmelreich S , Vogt R . 2007. A new subtribal classification of the tribe Anthemideae (compositae). Willdenowia. 37(1):89–114.
- Oberprieler C , Vogt R , Watson LE . 2007. Anthemideae. In: The families and genera of vascular plants VIII. Kubitzki K , editor. Springer. p. 342–374.
- Podlech D 1986. Tanacetum . In: Flora Iranica edited by Rechinger KH , Vol. 158. Gruck V. – Verlagsanstalt, pp. 88–148.
- Powell AM , Kyhos DW , Raven PH . 1974. Chromosome numbers in Compositae. X. Am J Bot. 61(8):909–913.
- Queiros M . 1975. Contribuicao para o conhecimento citotaxonomico das Spermatophyta de Portugal. X. Chenopodiaceae. Boletim da Sociedade Broteriana. Coimbra. 49:121–142.
- Romero Zarco C . 1986. A new method for estimating karyotype asymmetry. Taxon. 35(3):526–530.
- Rostovtseva TS . 1983. Chromosome numbers of some species of the family Asteraceae. II. Bot. Zhurn. 68:1538–1543.
- Semple JC , Watanabe K. 2009. A review of chromosome numbers in Asteraceae with hypotheses on chromosomal base number evolution. In: Funk VA , Susanna A , Stuessy TF , Bayer RJ , editors. Systematics, evolution and biogeography of the Compositae. Vienna: International Association of Plant Taxonomy. p. 61–72; 965 p.
- Sokolovskaya AP , Strelkova OS . 1948. Geografitcheskoe raspredelenie polyploidov. III. Issledovanie florei alpiskoi oblasti centralenogo Kavkazskogo krebta. Uchenye Zapiski, Leningr. Univ. 66:195–216.
- Sokolovskaya AP , Strelkova OS . 1960. Geographical distribution of the polyploid species of plants in the Euro-Asiatic Arctic. Bot. Zhurn. 45(3):369–381.
- Sonboli A , Kazempour Osaloo Sh , Riahi H , Mozaffarian V . 2010. Tanacetum joharchii sp. nov. (Asteraceae-Anthemideae) from Iran and its taxonomic position based on molecular data. Nord J Bot. 28(1):74–78.
- Sonboli A , Kazempour Osaloo Sh , Vallès J , Oberprieler C . 2011. Systematic status and phylogenetic relationships of the enigmatic Tanacetum paradoxum Bornm. (Asteraceae, Anthemideae): evidences from nrDNA ITS, micromorphological, and cytological data. Plant Syst Evol. 292(1–2):85–93.
- Sopova M , Sekovski Z . 1982. Chromosome atlas of some Macedonian angiosperms. III. Ann. Fac. Biol. Univ. Skopje. 35:145–161.
- Spooner DM , De Jong DC , Sun BY , Stuessy TF , Gengler KM , Nesom GL , Berry PE . 1995. Chromosome counts of Compositae from Ecuador and Venezuela. Ann MO Bot Gard. 82(4):596–602.
- Stebbins GL . 1971. Chromosomal evolution in higher plants. London: Edward Arnold.
- Strid A . 1987. Chromosome number of Turkish mountain plants. Notes Roy Bot Gard Edinb. 44(2):351–356.
- Strid A , Franzen R . 1984. Chromosome numbers in flowering plants from Greece. Willdenowia. 13(2):329–333.
- Stuessy TF , Lack HW. 2011. Monographic plant systematics: Fundamental assessment of plant biodiversity. A.R.G. Gantner Verlag K.G., FL-9491 Ruggell.
- Sz.-Borsos O . 1970. Contributions to the knowledge on the chromosome numbers of Phanerogams growing in Hungary and southeastern Europe. Acta Bot. Acad. Set. Hung. 16(3–4):255–265.
- Tahara M . 1915. Parthenogenesis in Erigeron annuus Pers. Bot Mag. 29(344):245–254.
- Tominaga Y . 1982. Studies on breeding of Pyrethrum flowers. Bull Hiroshima Agric Coll. 7(1):19–29.
- Torrell M , Vallès J , Garcia-Jakas N , Mozaffarian V , Gabrielian E . 2001. New or rare chromosome counts in the genus Artemisia L. (Asteraceae, Anthemideae) from Armenia and Iran. Bot J Linn Soc. 135(1):51–60.
- Vallès J , Garnatje T , Garcia S , Sanz M , Korobkov A . 2005. Chromosome numbers in the tribes Anthemideae and Inuleae (Asteraceae). Bot J Linn Soc. 148(1):77–85.
- Vilmorin R , Chopinet R . 1954. Contribution à l’étude des nombres chromosomiques der races et variétés cultivées chez nos plantes ornementales. Caryologia. 6:1006–1015.
- Watanabe W. 2013. Index to chromosome numbers in Asteraceae. Available from: http://www.lib.kobe-u.ac.jp/infolib/meta_pub/G0000003asteraceaeresult-en
- Zhmyleva AO , Kondo K . 2006. Comparison of somatic chromosomes in some species of Chrysanthemum sensu lato in Russia. Chromosome Bot. 1(1):13–22.
- Zhukova PG . 1964. The caryology of some species of Compositae growing in the arcto-alpine garden of the Botanic Garden. Kola Peninsula. Bot. Zhurn. 49:1656–1659.
- Zhukova PG. 1967. Karyology of some plants cultivated in the Arctic-Alpine Botanical Garden. Plantarum in Zonam Polarem Transportatio (Avrorin NA.). 2:139–149. Leningrad.
- Zhukova PG , Korobkov AA , Tikhonova AD . 1977. Chromosome numbers of some plant species in the eastern Arctic Yakutya. Bot. Zhurn. 62(2):229–234.