
Abstract
We present detailed cytogenetic properties of the Beyşehir nase, Chondrostoma beysehirense from Lake Beyşehir investigated through conventional Giemsa staining, C-banding and Ag-NOR in 10 specimens. The diploid number of chromosomes was 50, and the chromosomes in karyotypes were distinguished as metacentric, submetacentric/subtelocentric and acrocentric. Morphologically differentiated sex chromosomes could not be recognized. A secondary constriction was observed on the short arm of the sixth submetacentric chromosome pair. Some chromosomes had centromeric and pericentromeric C-bands, and the short arm of the sixth submetacentric chromosome pair was entirely C-positive. The active NOR was localized on the short arm of the sixth submetacentric chromosome pair, and this active NOR was associated with a large C-heterochromatin region. This NOR was recorded in all specimens examined and it occurred always in the homomorphic state on both homologous chromosomes. Systematics of Chondrostoma species will be better understood by using both molecular and FISH techniques in the future.
Introduction
Chondrostoma is a genus of ray-finned fish of the family Cyprinidae, and the genus comprises about 26 species that are commonly known as nases (Elvira Citation1997; Freyhof Citation2014). Perea et al. (Citation2010) investigated the phylogenetic relationships and biogeographical patterns of the circum-Mediterranean species of the subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. They proposed revision of taxonomy in certain Chondrostoma species (C. angorense, C. cyri, C. holmwoodii, C. knerii, C. meandrense, C. nasus, C. oxyrhynchu, C. phoxinus, C. prespensis, C. regium, C. soetta, C. vardarense, and C. olisiponensis), and included C. olisiponensis within a separate genus Iberochondrostoma.
Altogether, seven Chondrostoma species (C. angorense, C. colchicum, C. holmwoodii, C. meandrense, C. kinzelbachi, C. regium, and C. beysehirense) are reported to occur in various regions of Turkey (Elvira Citation1997; Küçük et al. Citation2007; Özcan Citation2008; İlhan Citation2009; Freyhof Citation2014). C. beysehirense is endemic to Turkey with a highly restricted distribution in Lake Beyşehir and a few streams connected to this freshwater lake in Central Anatolia (Bogutskaya Citation1997).
Most species belonging to the Leuciscinae subfamily have diploid number of 48, 50 or 52 chromosomes. Almost all of the Chondrostoma species have 2n = 50 but chromosome morphology varies between the species (Arai Citation2011). The conventionally stained karyotypes of Chondrostoma species were described from various geographic regions. Karyotype analysis of the king nase fish, C. regium, from Iran was reported by Esmaeili et al. (Citation2010), and chromosome studies of C. nasus from Europe were performed by Ráb et al. (Citation2008). However, information on differentially stained chromosomes and detailed structure of the karyotype in Chondrostoma species from Turkey is lacking. The aim of this paper is to perform a chromosomal banding analysis of the karyotype of C. beysehirense with the use of C-banding and Ag-NOR staining and to compare the findings with those obtained in previous studies of Chondrostoma species.
Materials and methods
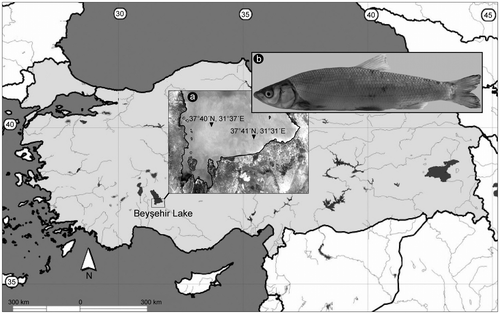
Ten specimens of C. beysehirense were collected from Lake Beyşehir, Konya, Turkey (Figure ). The fish specimens were transported alive to the laboratory, and kept in well aerated aquaria until analysis. Chromosome preparation was carried out according to the technique described by Collares-Pereira (Citation1992). After preparation of chromosome slides, conventional Giemsa-staining was carried out. Constitutive heterochromatin and nucleolar organizer regions (NORs) were detected with modified techniques of C-banding (Sumner Citation1972) and Ag-NOR staining (Howell and Black Citation1980), respectively. From each specimen, 10 to 20 slides were prepared, and at least 20 well-spread metaphase plates were analysed. The system of classification of chromosomes according to the centromere position was adopted after Levan et al. (Citation1964). The number of all chromosomal arms in the female complement (NF) was calculated.
Figure 1. (a) Sampling sites (37°40′N, 31°37′E; 37°41′N, 31°31′E) in Lake Beyşehir, Konya, Turkey of (b) Chondrostoma beysehirense.
Results
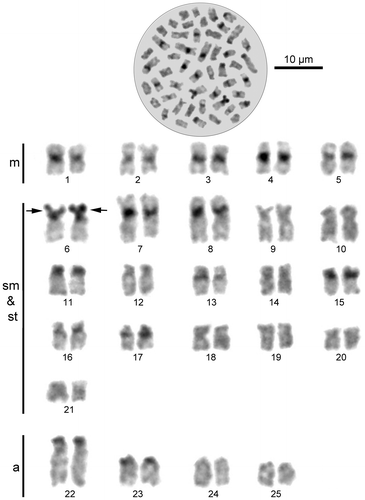
The karyotype of the Beyşehir nase consists of 50 chromosomes including five metacentric pairs (nos. 1–5), 16 submetacentric/subtelocentric pairs (nos. 6–21) and four acrocentric pairs (nos. 22–25) of autosomes (NF = 92). Morphologically differentiated sex chromosomes were not distinguished in the set. A secondary constriction was observed in the short arm of the autosomal pair no. 6 (Figure ).
Figure 2. Metaphase spread and standard Giemsa staining karyotype of Chondrostoma beysehirense. Arrows indicate the secondary constriction (m: metacentric, sm: submetacentric, st: subtelocentric, a: acrocentric).

The C-banded karyotype of the Beyşehir nase is illustrated in Figure . Distinct centromeric or pericentromeric C-bands were observed in some autosomes (nos. 1, 3, 4, 7, 8, and 15). Four bi-armed autosomal pairs (nos. 2, 5, 13, 16 and 17) and two acrocentric pairs (nos. 22 and 23) possessed slight centromeric or pericentromeric bands, whereas no C-positive staining was observed in the other autosomal pairs. The short arm of pair no. 6 was entirely C-positive.
Figure 3. Metaphase spread and C-banded karyotype of Chondrostoma beysehirense. Arrows indicate the C-band associated with the secondary constrictions (m: metacentric, sm: submetacentric, st: subtelocentric, a: acrocentric).
Active Ag-NOR regions were found on one biarmed autosomal pairs (no. 6) in the complements of all specimens studied (Figure ). The NOR was localized within the short arm which was C-positively stained.
Figure 4. Silver-stained metaphase spread and karyotype of Chondrostoma beysehirense. Arrows indicate the active Ag-NOR (m: metacentric, sm: submetacentric, st: subtelocentric, a: acrocentric).

Discussion
The cytogenetic features of C. beysehirense were investigated in this study for the first time. The diploid chromosome number of this species is 50, and there are four acrocentric chromosomes in the chromosome set (NF = 92). This finding is congruent with the reports of the karyotypes with 50 chromosomes found in most species of the genus Chondrostoma (Arai Citation2011). The only exception is C. regium from Iran which possesses 52 chromosomes and all the chromosomes are bi-armed (Esmaeli et al. Citation2010).
Rab et al. (Citation2008) investigated some cyprinid species by the use of the FISH technique. According to this study, the presence of a medium-sized submetacentric NOR-bearing chromosomal pair is ubiquitous among cytogenetically analysed leuciscine species, and the largest pair is characteristically a subtelocentric/acrocentric element representing a leuciscine cytotaxonomic marker. We have found a similar marker chromosome in the karyotype of C. beysehirense. The NOR-bearing chromosome was submetacentric or subtelocentric, and the NOR included almost the whole short arm. The NOR site occurred always in the homomorphic state on both homologous chromosomes. Rodrigues and Collares-Pereira (Citation1996) and Collares-Pereira and Rab (Citation1999) recorded NOR polymorphism in the karyotype of C. lusitanicum (2n = 50). The presence of the active-NORs was detected using both the silver nitrate and FISH techniques in one or two pairs of chromosomes of this species. Similarly to our results, some chromosomes in the karyotype of C. lusitanicum possessed centromeric heterochromatin areas (Rodrigues and Collares-Pereira Citation1996).
Recently, chromosomal banding studies have been performed in various cyprinid species in Turkey and other regions. Arslan and Takı (Citation2012) investigated eight specimens of the tench, Tinca tinca (Tincinae) from Lake Beyşehir and found 48 chromosomes in the diploid complement. A secondary constriction and an active NOR was recorded on the short arm of the third pair of chromosomes and all chromosomes had centromeric and pericentromeric C-bands. These findings are fairly similar to results obtained in C. beysehirense. Contrary to our results, Arslan and Takı (Citation2012) detected in the karyotype of T. tinca also an interstitial dark C-band on the short arm of a chromosome pair, and whole C-heterochromatic short arms in other three chromosome pairs. Gaffaroğlu et al. (Citation2006) determined in the karyotype of an Anatolian leuciscine cyprinid fish, Acanthobrama marmid, NORs in the telomeres of two pairs of medium-sized submetacentric or subtelocentric chromosomes. Nazari et al. (Citation2011) observed NORs in the karyotype of Alburnus filippii at the end of the short arm of one pairs of medium-sized subtelocentric chromosomes. Karasu et al. (Citation2009) found in the complement (2n = 50) of Pseudophoxinus firati from the Euphrates River system C-heterochromatin in the pericentromeric region of six pairs of chromosomes. The NORs were recorded on the short arms of two pairs of medium-sized submeta-subtelocentric chromosomes. These findings support the conservative nature of karyotypes and chromosomal banding patterns in cyprinid or leuciscinae fishes but, at the same time, they indicate moderate variation in certain characters such as the distribution of NORs and C-heterochromatin. This shows that chromosome banding techniques represent a useful tool in understanding chromosomal evolution in the respective systematic groups, and can find their use in the assessment of phylogenetic relationships between and within species. Therefore, it will be desirable to obtain further data on chromosomal banding patterns, as well as to apply molecular cytogenetic approaches in Chondrostoma species in Turkey and neighbouring regions. The cytogenetic studies should be combined with molecular phylogenetic investigations based on sequencing of DNA.
Funding
The paper was supported by grants from the Coordination Committee of Scientific Research Projects [BAP no. 15201011] of Selçuk University.
Acknowledgements
We would like to thank Jan Zima for editing the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Arai R. 2011. Fish karyotypes: a check list. Tokyo (Japan): Springer; p. 1–325.
- Arslan A, Takı FN. 2012. C-banded karyotype and nucleolar organizer regions of Tinca tinca (Cyprinidae) from Turkey. Caryologia. 65(3):246–249.
- Bogutskaya NG. 1997. Chondrostoma beysehirense, a new cyprinid fish from Beyşehir Lake. Central Turkey. Ichthyol Explor Freshwaters. 8(2):151–158.
- Collares-Pereira MJ. 1992. In vivo direct chromosome preparation (protocol for air drying technique). 1st International Workshop on Fish Cytogenetic Techniques. Concarneau-France; 1992 Sep 14–24; p. 15–9.
- Collares-Pereira MJ, Ráb P. 1999. NOR polymorphism in the Iberian species Chondrostoma lusitanicum (Pisces: Cyprinidae) – re-examination by FISH. Genetica. 105(3):301–303.
- Elvira B. 1997. Taxonomy of the genus Chondrostoma (Osteichthyes, Cyprinidae): an updated review. Folia Zool. 46(Suppl. 1):1–14.
- Esmaeili HR, Zareian H, Gholamhosseini A, Ebrahimi M, Gholami Z, Teimori A, Ansari TH. 2010. Karyotype analysis of the King Nase Fish, Chondrostoma regium (Heckel, 1843) (Actinopterygii: Cyprinidae) from Iran. Turk J Fish Aquat Sci. 10(4):477–481.
- Freyhof J. 2014. Chondrostoma beysehirense. The IUCN red list of threatened species. Version 2014.3; [cited 2014 Dec 26]. Available from www.iucnredlist.org
- Gaffaroğlu M, Yüksel E, Ráb P. 2006. Note on the karyotype and NOR phenotype of leuciscine fish Acanthobrama marmid (Osteichthyes, Cyprinidae). Biologia, Bratislava. 61(2):207–209.
- Howell WM, Black DA. 1980. Controlled silverstaining of nucleolus organizer regions with a protective colloidal developer: a 1 step method. Experientia. 36(8):1014–1015.
- İlhan A. 2009. Threatened fishes of the world: Chondrostoma beysehirense Bogutskaya, 1997 (Cyprinidae). Environ Biol Fish. 86(4):483–484.
- Karasu M, Yuksel E, Gaffaroğlu M. 2009. Karyotype, NORs, and C-banding analysis of Pseudophoxinus firati Bogutskaya, Küçük, Atalay, 2007 (Actinopterygii, Cyprinidae) in the Euphrates River. Turkey. Turk J Zool. 35(6):865–868.
- Küçük F, Gümüş E, Gülle İ, Güçlü SS. 2007. The fish fauna of the Göksu River (Türkiye): taxonomic and zoogeographic features. Turk J Fish Aquat Sci. 7(1):53–63.
- Levan A, Fredga K, Sandberg AA. 1964. Nomenclature for centromeric position on chromosomes. Hereditas. 52(2):201–220.
- Nazari S, Pourkazemi M, Porto JIR. 2011. Chromosome description and localization of nucleolus organizing regions (NORs) by Ag-staining technique in Alburnus filippii (Cyprinidae, Cypriniformes) in Anzali Lagoon. North Iran. Iranian J Fish Sci. 10(2):352–355.
- Özcan G. 2008. Threatened fishes of the world: Chondrostoma meandrense Elvira, 1987 (Cyprinidae). Environ Biol Fish. 83:297–298.
- Perea S, Böhme M, Zupančič P, Freyhof J, Šanda R, Özuluğ M, Abdoli A, Doadrio I. 2010. Phylogenetic relationships and biogeographical patterns in Circum-Mediterranean subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. BMC Evolutionary Biology. 10:265.
- Ráb P, Rábová M, Pereira CS, Collares-Pereira MJ, Pelikánová Š. 2008. Chromosome studies of European cyprinid fishes: interspecific homology of leuciscine cytotaxonomic marker – the largest subtelocentric chromosome pair as revealed by cross-species painting. Chromosome Res. 16(6):863–873.
- Rodrigues EM, Collares-Pereira MJ. 1996. NOR polymorphism in the Iberian species Chondrostoma lusitanicum (Pisces: Cyprinidae). Genetica. 98(1):59–63.
- Sumner AT. 1972. A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res. 75(1):304–306.