
Abstract
This study investigated C-heterochromatin and NOR (nucleolar organizer region) characteristics of 2n = 52 and 2n = 60 chromosomal forms of Nannospalax xanthodon (blind mole rat), which are separated by the Sakarya River in Bilecik Province. In addition, a new locality was included to the geographical border of 2n = 52 NF = 70 cytotypes. According to the results of C-banding, centromeric heterochromatin was detected in three pairs of chromosomes and telomeric heterochromatin was detected in three pairs of chromosomes in the 2n = 52 form. Sex chromosomes are negative heterochromatins. In the 2n = 60 form, a telomeric heterochromatin region was found in seven pairs of chromosomes. The sex chromosomes have centromeric heterochromatin. According to the NOR distributions of chromosomes, in 2n = 52 cytotypes, NOR was detected in the telomeric regions of four bi-armed chromosomes, while in 2n = 60 types active NOR was detected in telomeric regions of three bi-armed chromosomes.
1. Introduction
The worldwide distribution of the blind mole rats, members of the Spalacidae family, is in North-East Africa, the Balkans, South-East Europe, Central Asia, the Middle East, Russia and the Caucasus. It is known that this family emerged during the Oligocene epoch of the Paleogene Period in Anatolia or in its neighbours and spread to North Africa, the Middle East, Russia and the Balkans (Savic and Nevo Citation1990; Nevo Citation1991; Coşkun Citation2003). The population that passed to the Balkan Peninsula before the Lower Pleistocene remained separated from the Anatolian population due to the formation of the Istanbul and Çanakkale (Dardanelles) Straits, and therefore speciated independently (Ellerman and Morrison-Scott Citation1951; Harrison and Bates Citation1991; Mitchell-Jones et al. Citation1999).
In Turkey, Spalax and more commonly Nannospalax genus names were used for mole rats due to the problems in their taxonomies and systematics, complex karyotypes and morphological similarities (Aşan and Yağcı Citation2008; Coşkun et al. Citation2010; Arslan and Bölükbaş Citation2010; Arslan et al. Citation2011a). To date, karyological studies have recorded diploid chromosome numbers (2n) ranging from 36 to 60 for blind mole rats in Turkey (Ivanitskaya et al. Citation2008; Sözen et al. Citation2008). In a comprehensive review conducted by Arslan et al. (Citation2016) in three commonly described species of Nannospalax (25 in N. leucodon, 28 in N. xanthodon and 20 in N. ehrenbergi), the presence of 73 different chromosomal races corresponding to the cytotype have been presented. 2n = 60 cytotypes were recorded as Kastamonu and Vasvari and 2n = 52 cytotypes were recorded as Abant and Sebil in 28 different chromosomal races defined for Nannospalax xanthodon (N. xanthodon) which has the widest distribution area in Anatolia.
To clarify the karyotypic evolution in Israeli Spalax populations, Ivanitskaya et al. (Citation2005) studied heterochromatin differences in S. galili (2n = 52) and S. judai (2n = 60) by using C-banding, base-specific fluorochrome staining (CMA3/DA/DAPI), and comparative genomic hybridization (CGH) methods. The ancestral karyotype was found to be 2n = 60 and the importance of Robertsonian metacentrics and heterochromatin deletions caused by fusion events were pointed out in the occurrence of the karyotype differences.
Matur et al. (Citation2011) compared 15 blind mole rat specimens belonging to north, south, east and west of Anatolia with four different 2n = 50 forms using C- and G-banding techniques. According to the C-banding results, telomeric heterochromatin blocks are characteristic for the western blind mole rats of Turkey. Arslan and Zima (Citation2015b) compared four populations of N. xanthodon with 2n = 50 chromosomes to determine their karyotype characteristics using C-banding and AgNOR staining. NOR regions were observed in telomeric regions in the short arms of autosomes. The C-banding and NOR staining models are useful in explaining the evolutionary change of chromosomes and chromosomal characteristics of mole rats, although they are not entirely distinctive between cytotypes.
When karyological studies are evaluated, it is difficult to establish definite and clear rules on the degree of variability or the smallest geographic/ecological boundaries of species with highly variable chromosome numbers and morphologies. However, significant progress has been made in the determination of the phylogeny of the blind mole rats through molecular-based studies in recent years. Hadid et al. (Citation2012) identified four clades (leucodon, xanthodon, ehrenbergi, vasvarii) for Anatolian blind mole rats through phylogenetic analysis of five mtDNA sequences. According to Kryštufek et al. (Citation2012), the lowest genetic diversity was seen in N. leucodon and the highest genetic diversity was seen in N. xanthodon in their study using cytochrome b sequences in Nannospalax specimens. Kankılıç et al. (Citation2013) reported that random amplification of polymorphic DNA (RAPD) bands could be molecular markers among chromosomal races, using the RAPD technique to taxonomically distinguish chromosomal races or species. Other molecular level studies revealed significant differences between some cytotypes in Anatolia, although there is no difference at the species level (Arslan et al. Citation2010; Kandemir et al. Citation2012).
The aim of this study is to comparatively analyse the heterochromatin and nucleolus organizer regions (NOR) of 2n = 52 and 2n = 60 cytotypes of N. xanthodon species, spreading in the province of Bilecik located in the western Anatolia region, and to contribute to the determination of geographically spreading boundaries.
2. Materials and methods
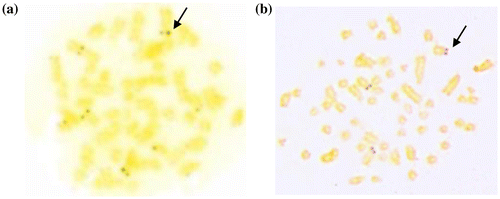
Seven blind mole rat specimens, from six different localities in the eastern and western parts of the Sakarya River separating Bilecik province into two regions, were studied karyologically (Figure ). The specimens were caught with live capture traps designed by Yağcı and Aşan (2007). Chromosome samples were prepared from bone marrow (Ford and Hamerton Citation1956). The C-banding was achieved according to the technique of Gosden (Citation1994) and NOR staining according to the method of Howell and Black (Citation1980). At least 20 well-spread and banded metaphase plates were photographed and arranged to determine the diploid chromosome number (2n), the autosomal fundamental number (NFa) and the fundamental number (NF). Chromosomes were classified according to Levan et al. (Citation1964). Heterochromatin and NOR distributions were examined in the arranged chromosome plates (Figures ).
Figure 1. Distribution of 2n = 52 and 2n = 60 chromosomal forms in and around the Bilecik province. (1) Kastamonu: 2n = 60 NF=74, 78,79 (Arslan et al. Citation2016); (2) Ankara: 2n = 60 NF=80 (Kankılıç et al. 2007b); (3) Eskişehir (İnönü): 2n = 60 NF=78; (4) Bilecik (Bozüyük): 2n = 60 NF=78; (5) Bilecik (İnhisar) 2n = 60 NF=78 (Matur and Sözen Citation2005); (6) Bilecik (Söğüt): 2n = 60 NF=78 (this study, n:1); (7) Bilecik (Pazaryeri): 2n = 60 NF=78 (this study, n:1); (8) Kütahya: 2n = 60 NF=76 (Sözen et. al. 2006); (9) Bursa (İnegöl): 2n = 60 W NF=78 (Ivanitskaya et al. Citation2008); (10) Yalova (Bursa road): 2n = 52 NF=70 (Sözen et al. Citation2013); (11) Yalova: 2n = 52 NF=72 (Matur et al. Citation2013); (12) Bilecik (Gülümbe): 2n = 52 NF=70 (this study, n:2); (13) Bilecik (Osmaneli): 2n = 52 NF=70 (this study n:1); (14) Bilecik (Gölpazarı, Bolatlı): 2n = 52, NF=70 (this study, n:1); (15) Bilecik (Yenipazar, Kavacık): 2n = 52 NF=70 (this study, n:1); (16) Bolu (Abant): 2n = 52 NF=70–72 (Arslan et al. Citation2016) (∆: 2n = 52, ▲: 2n = 60, :
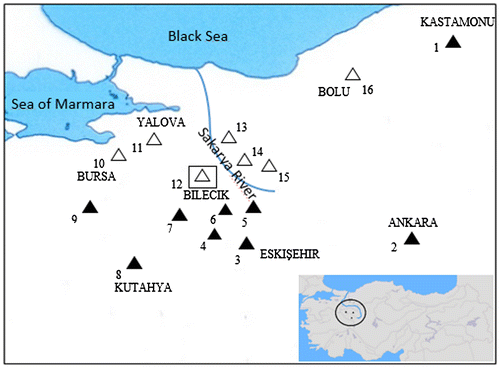
Figure 2. C band model in 2n = 52♀ (a, b) and 2n = 60♀ (c, d) metaphase plates of N. xanthodon.
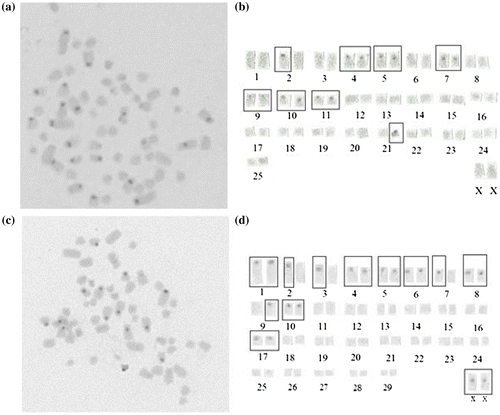
Figure 3. NOR zones in metaphase plates of 2n = 52♀ (a) and 2n = 60♀ (b) of N. xanthodon.
3. Results
3.1. Heterochromatin distribution and C-banding
In the C-band of the 2n = 52 form, it appears that three pairs of chromosomes (fourth, fifth and seventh pairs) have a centromeric heterochromatin region. Telomeric heterochromatin was detected in the ninth, 10th and 11th pair of chromosomes. The second and 21st pair of chromosomes have heteromorphic characteristics. Sex chromosomes are negative heterochromatins. In the C-band of the 2n = 60 form, it appears that seven pairs of chromosomes (first, fourth, fifth, sixth, eighth, 10th and 17th pairs) have a telomeric heterochromatin region. On the second, third, seventh and ninth pairs of chromosomes, a telomeric heterochromatin region was found with heteromorphic character. Sex chromosomes have centromeric heterochromatin characteristics.
3.2. NOR region distribution and NOR staining
In the 2n = 52 NF = 70 cytotype of N. xanthodon, four pairs of active NORs were detected in the telomeric regions of the short arms of bi-armed chromosomes. In the 2n = 60 NF = 78 cytotype, three pairs of active NORs were detected in the telomeric regions of the short arms of bi-armed chromosomes.
4. Discussion
In Bilecik province, N. xanthodon has two cytotypes including 2n = 52 NF = 70 and 2n = 60 NF = 78. The 2n = 52 karyotype, in the north of Anatolia (Karamürsel, Abant, Mudurnu, Nallıhan, Seben, Yeniçağa, Mengen), was recorded as NF = 70 by Sözen (Citation2004). In the province of Bilecik, Matur and Sözen (Citation2005) recorded 2n = 52, NF = 70 cytotypes in the eastern part of the Sakarya river and found 2n = 60, NF = 78 cytotypes in the western part of the river. The researchers recorded 2n = 52 chromosomal forms in specimens obtained from 3 km south, 15 km west of Gölpazarı and Yenipazar. They recorded 2n = 60 chromosomal forms in Kepirler village located in 10 km south-west of Bilecik province, 14 km north of Bozüyük and in Inhisar. Matur et al. (Citation2013) found that the NF value of 2n = 52 cytotypes was 72 in Yalova, in the study conducted in the west of Turkey. The researchers emphasized that the Yalova province is the westernmost point of this cytotype, expanding the spread of 2n = 52 chromosomal races. They also stated that the Sakarya River functions like a barrier between Yalova (2n = 52 NF = 72) and Bilecik and Bolu (2n = 52 NF = 70). Sözen et al. (Citation2013) gave the records of 2n = 52, NF = 70 cytotype from Yalova (Bursa road) and Bolu (Gölköy, Dörtdivan, Yelkenler Mengen and Gerede) locations. Arslan and Zima (Citation2015) gave 2n = 52 NF = 70 records from Gerede/Bolu by performing C and NOR band karyotyping analysis.
In this study, it was determined that the 2n = 52 cytotype also exists in Gülümbe (Bilecik) located in the west of the Sakarya river. Previous studies also recorded 2n = 60 as very common cytotype in this area. Combining previous records of Yalova and Bolu (2n = 52 NF=70,72) with our study, 2n = 52 NF=70 may be the last point of this cytotype extending towards the western part of Sakarya river in Bilecik province. In addition, the 2n = 52 NF = 70 form is a continuation of the populations recorded in Yalova (on Bursa road) and Bolu provinces. Sakarya River is found here to be effective in separating Bilecik province into two chromosomal forms as a geographical barrier. Recently, Arslan and Zima (Citation2015) examined N. xanthodon (Bolu, İçel) and N. ehrenbergi (Gaziantep) specimens of 2n = 52 chromosomal forms with various banding methods. According to their C-band results, in the N. xanthodon specimens centromeric heterochromatin regions were found in five pairs of bi-armed and four pairs of acrocentric chromosomes. Peri-centromeric bands were found in the sex chromosomes. In this study, there are no heterochromatin regions in the sex chromosomes of 2n = 52 specimens and they differ in with regard to heterochromatin distribution.
Ivanitskaya et al. (Citation2008) compared the 2n = 60 W and 2n = 60R cytotypes using G, C, AgNOR, and fluorochrome banding techniques. Although the karyotype of both cytotypes is similar to 2n = 60 as given in this study, the NOR region has different characteristics in terms of localization of heterochromatin blocks in chromosome pairs. Specimens from Inegöl (Bursa), the closest locality to the province of Bilecik, were recorded as 2n = 60 W cytotypes in which centromeric heterochromatin is small or absent in the majority of acrocentric autosomes. In our study, most of the acrocentric chromosomes are also C-negative. Detection of centromeric and pericentromeric heterochromatin regions in bi-armed chromosomes and detection of centromeric C blocks in sex chromosomes show similarities with the cytotypes examined in our study. İnegöl (Bursa) specimens contain NOR in the short arms of five bi-armed chromosomes; on the other hand, our specimens contain NOR in both arms of telomeric regions of three bi-armed chromosomes. Researchers have emphasized that intrapopulation and interpopulation heterochromatin variations, deletions, and the presence of B chromosomes seen in both cytotypes may be major factors in the extraordinary chromosomal diversity of the blind mole rats. Taking these results into consideration, two possibilities are considered for 2n = 52 cytotypes detected in a small area in Bilecik province, where 2n = 60 is intensively located. This cytotype was formed either by Robertsonian metacentrics, heterochromatin variations and deletions from 2n = 60 in the present region, or they are the continuation of Yalova, Bolu and Bursa (Inegol) populations which were determined as the westernmost point of 2n = 52. Further examination of 2n = 52 and 2n = 60 in the studied area and its vicinity by G-banding may help clarify the second possibility.
Matur et al. (Citation2011) through C-banding analysis found that telomeric blocks are characteristic for western blind mole rats in Turkey. In our study, the specimens also have western heterochromatin characteristics. In addition, the theory that climatic properties are not related to chromosomal alteration is supported in our study by variations in a small field in Bilecik province.
Kankılıç et al. (Citation2010) recorded four different chromosome forms in 10 different regions of Western Anatolia (2n = 36, 40, 56, 60). The 2n = 60 one from these forms has two different karyotypes, NF = 78 and NF = 84. The form recorded as NF = 78, has seven telocentric pairs, one submetacentric pair and 21 acrocentric pairs. The X chromosome from the sex chromosomes of both cytotypes is submetacentric in medium size and the Y chromosome is small acrocentric. The 2n = 60 cytotype given in our study is the same as NF=78 cytotype. Sözen et al. (Citation2006a) has karyologically examined the 10 chromosomal forms of Spalax leucodon (2n = 48 NF=71, 2n = 50 NF=70, 72, 2n = 54 NF=75, 2n = 56 NF=72, 78, 2n = 58 NF=72, 2n = 60 NF=74, 76, 77) which he named as a supergenus in 30 localities, and the two chromosomal forms of S. ehrenbergi (2n = 52 NF=74, 2n = 56 NF=72). The Kütahya population, which is given as 2n = 60, is separated from the 2n = 60 NF = 78 karyotypes in our study due to the fact that the Kütahya population is bordered by Bilecik and has karyological values of NF = 76 and NFa = 72. It is not surprising that there are differences between the examples of Bilecik and Kütahya provinces. In fact, even within a small area such as Bilecik province, there are karyological differences. Despite this extraordinary variation in chromosome number and structure, the spreading boundaries of chromosomal forms in Turkey and the factors influencing these borders have not been determined precisely. In order to understand the barrier effect of the river and its immediate surroundings, geographical, climatic and edaphic studies need to be conducted along with the cytogenetic studies.
Disclosure statement
No potential conflict of interest was reported by the author.
Funding
This study was funded by Scientific Research Project fund of Bilecik Şeyh Edebali University [grant number 2015-02.BŞEÜ.04-03].
References
- Arslan A, Bölükbaş F. 2010. C-heterochromatin and NORs distribution of mole rat, Nannospalax xanthodon from Aksaray, Turkey. Caryologia. 63:398–404.
- Arslan E, Gülbahçe E, Arıkoğlu H, Arslan A, Bužan EV, Kryštufek B. 2010. Mitochondrial divergence between three cytotypes of the Anatolian mole rat, Nannospalax xanthodon. Zool in the Middle East. 50:27–34.
- Arslan A, Zima J. 2015b. Chromosome banding pattern retrieves an independent origin of 2n = 50 chromosome populations of Nannospalax xanthodon from Turkey. Mamm Biol. 80:440–445.10.1016/j.mambio.2014.09.002
- Arslan A, Zima J. 2015. Heterochromatin distribution and localization of nucleolar organizing regions in the 2n = 52 cytotypes of Nannospalax xanthodon and N. ehrenbergi from Turkey. Zool Stud. 54:1–6.10.1186/s40555-014-0088-1
- Arslan A, Akan Ş, Zima J. 2011a. Variation in C-heterochromatin and NORs distribution among chromosomal races of mole rats (Spalacidae) from Central Anatolia, Turkey. Mamm Biol. 76(1):28–35.10.1016/j.mambio.2010.01.005
- Arslan A, Kryštufek B, Matur F, Zima J. 2016. Review of chromosome races in blind mole rats (Spalax and Nannospalax). Folia Zool. 65(4):249–301.
- Aşan N, Yağcı T. 2008. Karyotype and Hair Scale Structure of Nannospalax leucodon (Nordmann, 1840) from Central Anatolia (Rodentia: Spalacidae). Turk J Zool. 32:125–130.
- Coşkun Y. 2003. A study on the morphology and karyology of Nannospalax nehringi (Satunin 1898 (Rodentia: Spalacidae) from Northeast Anatolia, Turkey. Turk J Zool. 27:171–176.
- Coşkun Y, Ulutürk S, Kaya A. 2010. Karyotypes of Nannospalax (Palmer 1903) populations (Rodentia: Spalacidae) from central-eastern Anatolia, Turkey. Hystrix It J Mamm (ns). 21:89–96.
- Ellerman JR, Morrison-Scott TCS. 1951. Checklist of Palaeoarctic and Indian Mammals, 1758 to 1946, British Mus. (Nat. Hist.), 1–810.
- Ford CE, Hamerton JL. 1956. A colchicine hypotonic citrate, squash for mammalian chromosomes. Stain Technol. 31:247–251.10.3109/10520295609113814
- Gosden R. 1994. Chromosome analysis protocols methods in molecular biology. Totowa, NJ: Humana Press Inc.10.1385/0896032892
- Hadid Y, Németh A, Snir S, Pavlíček T, Csorba G, Kázmér M, Major A, Mezhzherin S, Rusin M, Coşkun Y, Nevo E. 2012. Is evolution of blind mole rats determined by climate oscillations? PLoS One. 7(1):e30043.10.1371/journal.pone.0030043
- Harrison DL, Bates PJJ. 1991. Mammals of Arabia. Second Edition Harr Zool Mus Pub. 357:1–353.
- Howell WM, Black DA. 1980. Controlled silver staining of nucleolar organizer regions with a protective colloidal developer: a 1-step method. Experientia. 36:1014–1015.10.1007/BF01953855
- Ivanitskaya E, Belyayev A, Nevo E. 2005. Heterochromatin differentiation shows the pathways of karyotypic evolution in Israeli mole rats (Spalax, Spalacidae, Rodentia). Cytogenet Genome Res. 111:159–165.10.1159/000086386
- Ivanitskaya E, Sözen M, Rashkovetsky L, Matur F, Nevo E. 2008. Discrimination of 2n = 60 Spalax leucodon cytotypes (Spalacidae, Rodentia) in Turkey by means of classical and molecular cytogenetic techniques. Cytogenet Genome Res. 122:139–149.10.1159/000163091
- Kandemir İ, Sözen M, Matur F, Kankılıç M, Martinkova N, Çolak F, Özkurt SÖ, Çolak E. 2012. Phylogeny of species and cytotypes of mole rats (Spalacidae) in Turkey inferred from mitochondrial cytochrome b gene sequences. Folia Zool. 61(1):25–33.
- Kankılıç T, Kankılıç T, Şeker PS, Çolak R, Selvi E, Çolak E. 2010. Contributions to the Karyology and Distribution Areas of Cytotypes of Nannospalax leucodon (Rodentia: Spalacidae) in Western Anatolia. Acta Zool Bulg. 62(2):161–167.
- Kankılıç T, Kankılıç T, Sözen M, Çolak E. 2013. Genetic Diversity and Geographic Variation of Chromosomal races of Nannospalax xanthodon (Nordmann, 1840) and Nannospalax ehrenbergi (Nehring, 1898) from Turkey, Revealed by RAPD Analysis. Acta Zool Bulg. 65(1):45–58.
- Kryštufek B, Ivanıtskaya E, Arslan A, Arslan E, Bužan EV. 2012. Evolutionary history of mole rats (genus Nannospalax) inferred from mitochondrial cytochrome b sequence. Biol J Lin Soc. 105:446–455.10.1111/bij.2012.105.issue-2
- Levan A, Fredga K, Sandberg AA. 1964. Nomenclature for centromeric position on chromosomes. Hereditas. 52:201–220.
- Matur F, Sözen M. 2005. A karyological study on Subterrranean Mole Rats of the Spalax leucodon Nordmann, 1840 (Mammalia: Rodentia) superspecies around Bilecik province in Turkey. Zool Middle E. 36:5–10.10.1080/09397140.2005.10638121
- Matur F, Çolak F, Sevindik M, Sözen M. 2011. Chromosome differentiation of four 2n = 50 chromosomal forms of Turkish mole rat. Zoolog Sci. 28:61–67.10.2108/zsj.28.61
- Matur F, Çolak F, Ceylan T, Sevindik M, Sözen M. 2013. Chromosomal evolution of the genus Nannospalax (Palmer 1903) (Rodentia, Muridae) from western Turkey. Turk J Zool. 37:470–487.
- Mitchell-Jones AJ, Amori G, Bogdanowicz W, Krystufek B, Reijnders PJH, Spitzwnberger F, Stubbe M, Thissen JBM, Vohralik V, Zima J. 1999. The atlas of Europaen mammals. London: Academic Pres; p. 262–263.
- Nevo E. 1991. Evolutionary theory and processes of active speciation and adaptive radiation in subterranean mole rats, Spalax ehrenbergi superspecies in Israel. Evol Biol. 25: 1–125.
- Savic I, Nevo E. 1990. The Spalacidae; Evolutionary history, speciation and population biology. In: Evolution of subterranean mammals at the organismal and molecular levels, (E Nevo and AO Reig, editors. New York, NY: Alan R. Liss; p. 129–153.
- Sözen M 2004. A karyological study on subterranean mole rats of the Spalax leucodon
- Sözen M, Matur F, Çolak E, Özkurt Ş, Karataş A. 2006a. Some karyological records and a new chromosomal form for Spalax (Mammalia: Rodentia) in Turkey. Folia Zool. 55(3):247–256.
- Sözen M, Matur F, Nevo E. 2008. Discrimination of 2n = 60 Spalax leucodon cytotypes (Spalacidae, Rodentia) in Turkey by means of classical and molecular. Cytogenet Genome Res. 122:139–149.
- Sözen M, Çolak F, Sevindik M, Matur F. 2013. Cytotypes of Nannospalax xanthodon (Satunin, 1898) (Rodentia, Spalacidae) from western Anatolia. Turk J Zool. 37:462–469.